Влияние разного уровня техногенного засоления на некоторые показатели обмена веществ растений

Автор: Кусакина Маргарита Григорьевна, Еремченко Ольга Зиновьевна, Четина Оксана Александровна
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Экология
Статья в выпуске: 1, 2011 года.
Бесплатный доступ
Представлены результаты исследований содержания РНК и фенольных соединений у растений, произрастающих в условиях техногенного засоления. Показано, что различия в содержании различных форм фенольных соединений и РНК зависят как от видовой специфичности, так и от уровня засоления.
Техногенное засоление, метаболизм растений, фенольные соединения
Короткий адрес: https://sciup.org/147204538
IDR: 147204538 | УДК: 581.1.632.1
Influence of different level technogenic salinization on some indicators of a metabolism of plants
Results of researches of content РНК and phenolic connections at the plants growing in the conditions of technogenic salinization are presented. It is shown that distinctions in the content of various forms of phenolic connections and РНК depend both on specific species of specificity, and from level salinization.
Текст научной статьи Влияние разного уровня техногенного засоления на некоторые показатели обмена веществ растений
Современный техногенез и, в частности, добыча полезных ископаемых сопровождаются рядом отрицательных последствий, оказывающих влияние на все компоненты ландшафта. В результате антропогенного прессинга происходит нарушение почвеннорастительного покрова, что наблюдается, в частности, в зоне солеотвалов Верхнекамского месторождения солей, где добывается минеральное сырье для магниевого производства, получения калийных удобрений, технической и пищевой соли. Основным компонентом отходов является хлорид натрия. Около солеотвалов произрастают травяные растения, адаптировавшиеся к засоленной среде (Еремченко, Лымарь, Орлова, 2005; Еремченко, Лымарь, 2007).
Значимость эколого-биохимических исследований в этом направлении связана с недостаточной изученностью экосистем, формирующихся в условиях промышленного производства и добычи минерального сырья.
Действие техногенного солевого стресса снижает не только продуктивность, но и биоразнообразие фитоценозов. Как отмечают В.С. Никитина и М.М. Абдуллин (2009) в условиях техногенной нагрузки изменения в содержании вторичных метаболитов и, в частности, фенольных соединений, могут служить биоиндикатором состояния растений и среды их обитания. Накапливаясь в органах растений в ответ на действие стрессовых факторов, они повышают устойчивость организмов к влиянию таких неблагоприятных факторов среды, как ионизирующее излучение, засоление, низкие положительные температуры и др. (Запрометов, 1993).
В зависимости от концентрации, пространственной изомерии и формы они могут проявлять свое действие как ингибиторы или стимуляторы ростовых процессов растений (Кефели, 1978). Оказывая влияние на проницаемость мембран для ионов и активность мембраносвязанных ферментов, они вызывают изменения в энергетических и метаболических процессах (Simagina, Glumova, 2008). Кроме того, фенольные соединения проявляют антиоксидантные свойства, связывая активные формы кислорода и другие свободные радикалы, возникающие при действии стрессовых факторов (Загоскина и др., 2007).
Действие срессовых факторов среды приводит к таким структурным и функциональным изменениям организмов, которые направлены на поддержание их жизнедеятельности в новых условиях. В значительной степени это поддержание определяется взаимной сопряженностью процессов синтеза и распада за счет механизмов саморегуляции на разных уровнях организации живого. Среди этих механизмов существенную роль играют приспособительные изменения в обмене макромолекул, связанные с деятельностью генетического аппарата. Процессы, происходящие на уровне транскрипции и трансляции, деградации биополимеров и взаимодействий разных компартментов клетки, быстро реагируют на стрессовые изменения условий среды (Блехман, Шеламова, 1992).
В связи с этим целью нашей работы явилось изучение влияния антропогенного засоления на содержание РНК и фенольных соединений в растениях.
Методика исследований
Объектами исследований являлись растения, принадлежащие к различным экологическим группам: гликофиты – донник желтый ( Melilotus officinalis (L.) Pall., одуванчик Остенфельда ( Taraxacum ostenfeldii Raunk.), мать-и-мачеха обыкновенная ( Tussilago farfara L.) и галофит молокан татарский ( Lactuca tatarica (L.) C.A. Mey.), встречающиеся как в зоне устойчивого засоления (на расстоянии 1–5 м
от солеотвала, так и на удалении от складирования отходов (50–70 м) – зона неустойчивого засоления. Пробы для анализов были взяты в июне 2009 г. в районе солеотвалов г. Соликамска.
Определение РНК проводили по методу Волгина и Партье, основанному на свойстве пуриновых и пиримидиновых оснований интенсивно поглощать ультрафиолетовые лучи в зоне 270 ммк (Ермаков и др., 1987).
Фракционирование фенольных соединений проводили по методу М.Н. Запрометова (1971). Он основан на различной растворимости фенольных соединений в органических растворителях. Первую фракцию экстрагировали серным эфиром, вторую – этилацетатом, третью – этиловым спиртом. Экстракцию проводили на холоде до полного извлечения фенолов.
Определение фенольных соединений проводили по методу Левенталя. Он основан на легкой окисляемости фенольных соединений перманганатом калия на холоде в присутствии индикатора индигокармина.
Была проведена статистическая обработка результатов, определена достоверность различий между контрольным и опытным вариантами при 95%-ном уровне значимости.
Результаты и их обсуждение
Засоление корневой среды является многофакторным воздействием, вызывающим значительные изменения физиолого-биохимических процессов в растениях. Как отмечают И.Л. Радюкина и др. (2007), устойчивость гликофитов и галофитов в основном определяется одними и теми же протекторными механизмами. Но наблюдаемые различия в толерантности этих двух групп растений к стресс-факторам зависят от многообразия регуляторных механизмов и обусловлены сложной системой регуляции экспрессии стресс-зависимых генов (Dajic, 2006).
Интенсивность и направленность внутриклеточных биохимических процессов в значительной мере регулируются нуклеиновыми кислотами. Изучение нуклеинового обмена имеет особое значение в связи с выяснением механизмов адаптации растительных организмов к высоким концентрациям солей, в том числе, к техногенному засолению.
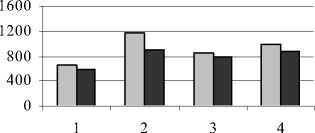
Нами исследовалось влияние техногенных солей на содержание РНК в листьях и корнях растений. Как показали опытные данные, устойчивое техногенное засоление по сравнению с неустойчивым вызвало более значительное снижение количества РНК как в листьях, так и в корнях у одуванчика Остенфельда, обладающего более высоким конститутивным уровнем РНК по сравнению с другими видами (рис. 1).
Уменьшение содержания РНК в условиях действия техногенных солей свидетельствует о пере- ходе метаболизма на более низкий уровень. По-видимому, это является защитно-приспособительной реакцией растений на неблагоприятные условия. Как отмечают Г.В. Удовенко и Л.А. Чудинова (1986), при высоком содержании хлоридов в почве наблюдается значительное повышение активности фермента РНК-азы, с деятельностью которой связан распад РНК. Другой вероятной причиной снижения нуклеиновых кислот в клетке при засолении может быть снижение их синтеза. Это связано с тем, что избыток засоляющих ионов подавляет активность ферментов биосинтеза нуклеиновых кислот.
Листья
Корни
-
□ неустойчивое засоление □ устойчивое засоление
Рис. 1 . Содержание РНК в листьях и корнях растений, (мг на 1 г сухой массы):
-
1 – донник желтый; 2 – одуванчик Остенфельда; 3 – мать-и-мачеха обыкновенная; 4 – молокан татарский
Устойчивое техногенное засоление существенно не повлияло на содержание РНК у таких видов, как галофит молокан татарский и гликофит мать-и-мачеха обыкновенная. Сравнивая количество РНК в органах растений в условиях разного уровня засоления, можно отметить, что техногенные соли в большей степени оказали ингибирующее влияние на содержание РНК в листьях по сравнению с корнями.
Как отмечают Е.И. Ценов, В.В. Кабанов, Б.П. Строгонов (1983), уменьшение синтеза РНК под действием высоких концентраций хлорида натрия может быть связано с уменьшением активности рибосом при повышении ионной силы раствора.
В адаптации растений к техногенному засолению важная роль принадлежит веществам фенольной природы, которые являются уникальными вторичными метаболитами. Они обладают широким биологическим действием, регулируют ряд внутриклеточных процессов, оказывают влияние на синтез белков и нуклеиновых кислот (Антонюк, Косян, Таран, 2009; Никитина, Абдуллин, 2009).
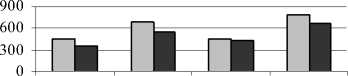
При изменении условий среды и, в частности, при воздействии техногенного засоления в составе фенольного комплекса происходят определенные изменения. Результаты наших исследований показали, что под влиянием устойчивого техногенного засоления в листьях всех изученных растений, независимо от уровня их конститутивной солеустой-чивости, произошло снижение суммарного количества фенольных соединений по сравнению с зоной неустойчивого засоления (рис. 2).
Листья
Корни
□ неустойчивое засоление □ устойчивое засоление
Рис. 2 . Общее содержание фенольных соединений в листьях и корнях растений (мг на 1 г сухой массы):
1 – донник желтый; 2 – одуванчик Остенфельда; 3 – мать-и-мачеха обыкновенная; 4 – молокан татарский
Вероятно, это связано с тем, что под влиянием техногенных солей происходит уменьшение активности ферментов биосинтеза фенольных соединений. В большей степени это проявилось в листьях гликофита донника желтого, характеризующегося более низкой солеустойчивостью по сравнению с другими видами. Устойчивое засоление менее значительно отразилось на содержании фенольных соединений в корнях у соленакапливающего галофита молокана татарского. В основе солеустойчивости этого растения лежит выраженная солеаккумулирующая способность. Молокан татарский накапливает свободные ионы натрия для создания повышенного осмотического давления клеточного сока.
В отличие от галофита молокана татарского у исследуемых гликофитов отсутствуют эволюционно сложившиеся механизмы солеустойчивости. Их адаптация, по-видимому, идет за счет эффективных неспецифических механизмов засухоустойчивости. Важной ее характеристикой, также как и солеустойчивости, является способность к осморегуляции (Hare, Cress, Van Staden, 1998).
Как и соленепроницаемые галофиты, гликофиты стремятся снизить накопление засоляющих ионов, затрачивая большое количество энергии, что- бы противостоять накоплению солей в организме. Фенольные соединения могут использоваться этими растениями в качестве субстрата для дыхания, повышая тем самым их энергообеспеченность (Филатова, Еремченко, Кусакина, 2009).
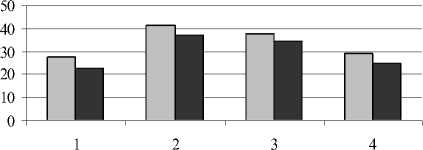
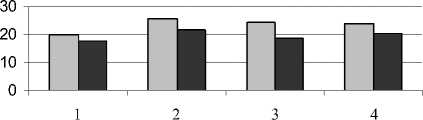
Кроме общего содержания, нами определялся фракционный состав фенольных соединений – эфирная, этилацетатная и спиртовая фракции (рис. 3).
Эфирная фракция
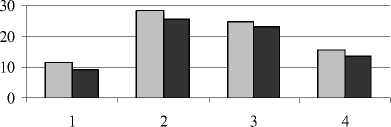
Этилацетатная фракция
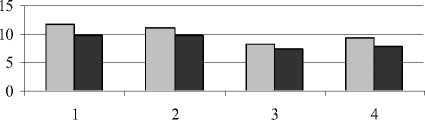
□ неустойчивое засоление □ устойчивое засоление
Рис. 3 . Содержание фенольных соединений разных фракций в листьях растений (мг на 1 г сухой массы):
1 – донник желтый; 2 – одуванчик Остенфельда; 3 – мать-и-мачеха обыкновенная; 4 – молокан татарский
Проведенные исследования показали, что гликофит одуванчик Остенфельда отличался более высоким уровнем фенольных соединений эфирной фракции по сравнению с другими растениями. Таким образом, высокий конститутивный уровень фенолов у одуванчика в значительной степени обусловлен эфирорастворимыми формами.
В состав эфирной фракции фенольных соединений входят такие кислоты, как кофейная, феруловая, синаповая, n-оксикоричная и др. Их характерной особенностью является способность к цистранс-изомерии. Цис-формы стимулируют рост растений, а транс-формы или не оказывают влияния на этот процесс, или подавляют его. Цисформы оксикоричных кислот, запасая некоторое количество энергии, играют важную роль в метаболизме и адаптации растений к действию неблагоприятных факторов (Запрометов, 1993).
Усиление техногенного засоления вызвало несколько большее снижение фенольных соединений эфирной фракции у донника и одуванчика. Как отмечает Т.В. Ряховская (1973), избыток в питательной среде солей натрия способствует накоплению эфирорастворимых форм при заметном снижении гликозидов этилацетатной фракции. Техногенные соли существенно не повлияли на количество этих соединений в листьях мать-и-мачехи, однако в корнях у данного вида произош- ло уменьшение количества фенолов этилацетатной фракции. Этилацетатная фракция включает в свой состав такие соединения, как лейкоантоцианы, гликозиды флавонолов, хлорогеновая кислота и др. Более высокий уровень фенольных соединений этой фракции характерен для донника желтого по сравнению с другими видами.
В состав спиртовой фракции входят такие соединения, как флавоноиды, гликозиды, конденсированные дубильные вещества и другие соединения, разнообразные по своему физиологическому воздействию. Как показали наши исследования, техногенное засоление не оказало существенного влияния на количество фенольных соединений спиртовой фракции. Аналогичные данные были получены В.С. Никитиным и М.Н. Абдуллиным (2009). Они также отмечают, что изменение количественного состава фенольного комплекса под влиянием стрессовых факторов среды в большей степени происходит за счет водной и эфирной фракции. Таким образом, в осуществлении приспособительных функций фенольных соединений роль отдельных фракций неравноценна.
Выявленные изменения в содержании РНК и фенольных соединений свидетельствуют о мобилизации защитных сил растительных организмов в условиях повышенного техногенного засоления корневой среды.
Список литературы Влияние разного уровня техногенного засоления на некоторые показатели обмена веществ растений
- Антонюк Т.М., Косян А.М., Таран Н.Ю. Флавоноиды как биомаркеры абиотического стресса на примере представителей рода Rhododendron L.//Материалы VII Междунар. симп. по фенольным соединениям. М., 2009. С. 20-21.
- Блехман Г.И., Шеламова Н.А. Синтез и распад макромолекул в условиях стресса//Успехи совр. биол. 1992. Т. 112, вып. 2. С. 281-287.
- Еремченко О.З., Лымарь О.А., Орлова Н.В. Солевой обмен растений в условиях техногенного засоления//Вестн. Перм. ун-та. 2005. Вып. 6. Биология. С. 164-167.
- Еремченко О.З., Лымарь О.А. Почвенно-экологические условия зоны солеотвалов и адаптация к ним растений//Экология. 2007. Вып. 1. С. 18-23.
- Ермаков А.И. и др. Методы биохимического исследования растений. Л.: Агропромиздат, 1987. 430 с.
- Загоскина Н.В., Гончарук Е.А., Алявина А.К. Изменения в образовании фенольных соединений при действии кадмия на каллусные культуры, инициированные из различных органов чайного растения//Физиол. раст. 2007. Т. 54, № 2. С. 267-274.
- Запрометов М.Н. Фенольные соединения и методы их исследования//Биохимические методы в физиологии растений. М.: Наука, 1971. С. 185-197.
- Запрометов М.Н. Фенольные соединения: распространение, метаболизм и функции в растениях. М.: Наука, 1993. 245 с.
- Кефели В.И. Рост растений и природные регуляторы//Физиол. раст. 1978. Т. 25, № 5. С. 65-73.
- Никитина В.С., Абдуллин М.И. Растительные фенольные соединения -индикаторы промышленного загрязнения среды//Материалы VII Междунар. симп. по фенольным соединениям. М., 2009. С. 188-189.
- Радюкина И.Л. и др. Изучение индуцибельных и конститутивных механизмов устойчивости к солевому стрессу у гравилата городского//Физиол. раст. 2007. Т. 54, № 5. С. 692-698.
- Ряховская Т.В. Флавоноидные компоненты полыни метельчатой//Фенольные соединения и их физиологические свойства. Алма-Ата, 1973. С. 76-78.
- Удовенко Г.В., Чудинова Л.А. Влияние засоления среды на изменение активности РНК-азы и ДНК-азы у растений разного уровня солеустойчивости//Физиол. раст. 1986. Т. 33, вып. 6. С. 166-172.
- Филатова Л.А., Еремченко О.З., Кусакина М.Г. Содержание фенольных соединений у представителей разных групп галофитов//Материалы VII Междунар. симп. по фенольным соединениям. М., 2009. С. 275-276.
- Ценов Е.И., Кабанов В.В., Строгонов Б.П. Влияние NaCl на содержание и синтез различных фракций нуклеиновых кислот в апексах стебля гороха//Физиол. раст. 1983. Т. 3, вып. 3. С. 537-544.
- Dajic Z. Salt stress//Physiology and Molecular Biology of stress tolerance in plants. 2006. P. 41-101.
- Hare P.D., Cress W.A., Van Staden J. Dissecting the roles of osmolyte accumulation during stress//Plant, Cell and Environ. 1998. № 6. P. 535-553.
- Simagina N.O., Glumova N.V. Phenol substances of Artemisiа santonica L., Halocnemum strobilaceum (Pall) M. Bied. with allelopathic activity//Biology, chemistry. 2008. Vol. 21, № 2. Р. 113-120.
