Влияние сеноманских вод на сообщества раковинных амеб

Автор: Кулюкина Е.В., Карташев А.Г.
Журнал: Вестник Нижневартовского государственного университета @vestnik-nvsu
Рубрика: Экология животных
Статья в выпуске: 1, 2017 года.
Бесплатный доступ
В данной статье представлены результаты исследования по воздействию сеноманских вод на численный и видовой состав сообщества раковинных амеб в светло - серой лесной почве. Было показано, что почвенные беспозвоночные животные могут быть использованы в качестве биоиндикаторов уровня загрязнения окружающей среды. Изучалось влияние сеноманских вод апт-сеноманского водоносного комплекса на сообщества раковинных амеб в лабораторных условиях. Сеноманские воды вносились в концентрациях 50, 100, 200 г/кг сухой почвы, в качестве контроля рассматривался участок без внесения концентрации. Загрязнение производилось искусственно сеноманскими водами (Среднеугутского месторождения), имеющей следующие характеристики: величина минерализации 19,3 г/л., общая жесткость составляет 29,1 мг - экв/дм³., хлор имеет показатель (11347,3-12431,3 г - экв/дм³), натрий - (6650,2-7400,3 г - экв/дм³), ионы кальция - (44,0-53,7 г - экв/дм³) ионы магния - (0,9-12,7 г - экв/дм³), гидрокарбонат - ионы присутствуют в количестве 1,8-5,4 г - экв/дм³...
Раковинные амебы, почвенные беспозвоночные, сеноманские воды, пластовые воды
Короткий адрес: https://sciup.org/14116974
IDR: 14116974 | УДК: 574.21
The influence of cenomanian water on testate amoebae community
This article presents the results of a study on the effects of Cenomanian water on the numerical and species composition of testate amoebae community in a light gray forest soil. It was shown that soil invertebrates can be used as bioindicators of the pollution of the environment. We investigated the effect of water Cenomanian Aptian-Cenomanian water-bearing complex in the community of testate amoebae in the laboratory. Cenomanian water were made at concentrations of 50, 100 and 200 g/kg dry soil as control was considered the site without any concentration. Pollution produced artificially Cenomanian water (Sredneuralskie field) having the following characteristics: the amount of salinity of 19.3 g/l, total hardness is 29.1 mg - EQ/dm3., chlorine has an index (11347,3-12431,3 g - EQ/dm3), sodium (6650,2-7400,3 g - EQ/dm3) calcium ions - (44,0-53.7 g - EQ/dm3) magnesium ions - (0,9-12,7 g - EQ/dm3), hydrogen carbonate ions present in the amount of 1,8-5,4 g - EQ/dm3. Calculation and analysis of invertebrate organisms was performed using the microscope BIOMED-2 at magnification ×160...
Текст научной статьи Влияние сеноманских вод на сообщества раковинных амеб
Развитие нефтедобывающей отрасли Западной Сибири привело к региональным экологическим изменениям. Нарушение естественной среды обитания начинается с проведения разведочных работ. Механическому разрушению подвергается почвенный покров на кустовых площадках, буровых скважинах, автодорогах. Высокотоксичные буровые отходы складируются в непосредственной близости от водоемов, что приводит к их долговременному загрязнению. Снеговые и дождевые воды способствуют распространению токсических веществ и негативному влиянию их на растительные и животные сообщества (Борисович 1985: 43).
Разливы высоко полиминерализирован-ных сеноманских вод приводят к деградации биоценозов па площадях в десятки квадратных километров. В зависимости от типа экосистемы, количеств нефтезагрязнений и площади разливов, можно выделить различные уровни деградации экосистем с характерными комплексами биоиндикационных показателей. Степень нарушености растительного покрова зависит от глубины проникновения сеноманских вод в почвенный слой. Наиболее чувствительными являются растения с поверхностным типом корневой системы, к наиболее устойчивым относятся многолетние кустарники и деревья. Деградация поверхностного слоя приводит к существенному изменению плодородия почв и сообществ беспозвоночных животных (Карташев 2007: 218). Состояние сообществ почвенных простейших при воздействии нефти изменяется в зависимости от уровня деграда- ции почв (Залялетдинова, Карташев 2016; Карташев, Смолина 2011). В естественных условиях сложно выявить зависимость трансформации биосистем от концентрации нефтезагряз-нения и длительности его воздействия.
Для мониторинга почв наиболее широко используемым методом остается анализ уровней концентраций токсичных соединений с использованием физико-химических методов. Для получения наиболее точных данных, включая весь комплекс воздействующих факторов, необходимо использовать не только химический анализ, но и внедрять новые подходы. К таковым относятся биоиндикационные методы, позволяющие определить состояние среды по изменению состояния живых существ (Шмара-ев, Шорина 2009: 642-645). Биоиндикационные методы в настоящее время широко используются в экологических исследованиях. К наиболее чувствительным биоиндикаторам относятся почвенные микроорганизмы, в том числе и раковинные амебы (Биоиндикация ... 1988).
Раковинные амебы среди почвенных простейших являются одним из самых удобных объектов для почвенно-зоологических исследований, что связано с наличием твердой раковинки, сохраняющейся в почве даже после отмирания самой амебы и дающей сведения не только о таксономическом статусе организма, но и о составе жизненных форм и экологических групп в локальной фауне. Сообщества раковинных амеб распространены достаточно широко во всех влажных почвах и устойчивы к нефтезагрязнениям. В то же время недостаточная изученность влияния сеноманских вод на сообщества раковинных амеб не позволяет перейти к комплексной биоиндикационной оценке уровня деградации почв при нефтедобыче. Целью исследований являлось изучение изменений сообществ раковинных амеб в зависимости от концентрации сеноманских вод природных комплексов.
Методика исследований
Объектом исследования являлись сообщества раковинных амеб. Строение амеб характеризуется наличием твердой раковинки, сохраняющейся в почве после отмирания самой амебы и дающей сведения о таксономическом статусе организма, о жизненных формах и экологических группах в локальной фауне. В работе рассматривалось влияние сеноманских вод на раковинных амеб, относящихся к 11 видам: Euglypha anodonta, Chlamydohprys minor, Phry- ganella acropodial, Cryptodifflugia compressa, Cyclopyxis aroelloides, Nebela dentistoma, Nebela militaris, Nebela lageniformis, Nebela collaris, Centropyxis vandeli, Plagiopyxis glyphostoma.
Исследования проводились в контролируемых лабораторных условиях в течение четырех недель с концентрациями внесения сеноманских вод: 50, 100, 200 г/кг почвы при параллельном контроле. Опыты проводились в пластиковых кюветах с размерами: 0,5x0,2x0,15 м. В лабораторные кюветы помещалась смешанная проба гумусового слоя (Ai, 0-20 см) серых лесных почв зернистокомковатой структуры влажностью 35-40%, по килограмму почвы в каждой кювете. Каждая кювета делилась на две части: опытную и контрольную.
Пробы почв для анализа брали в поверхностном горизонте на глубине 0-10 см. Для исследования влияния высокополиминерализиро-ванных сеноманских вод на сообщества раковинных амеб использовались минеральные, подземные воды апт-альб-сеноманского водоносного комплекса нижнего гидрогеологического этажа Среднеугутского месторождения. Воды относятся к классу соленых, с величиной минерализации 19,3 г/л. Они сохраняют хлорид-ный натриевый состав и хлоркальциевый тип, имеют пластовую температуру от 28 до 35 °C (Сулин 1935: 174). Водородный показатель варьирует от 5,5 до 7,6 и изменяется от слабокислых до слабощелочных. Хлор является доминирующим ионом в общем химическом облике и в ряду катионов стоит на первом месте и составляет 11347,3-12431,3 г-экв/дм3, натрий стоит на втором месте и составляет 6650,27400,3 г-экв/дм3, ионы кальция (44,053,7 г-экв/дм3) превалирует над ионами магния (0,9-12,7 г-экв/дм3). Гидрокарбонат-ионы присутствуют в количестве 1,8-5,4 г-экв/дм3. Воды очень жесткие, диапазон проявлений общей жесткости неширок и составляет 29,1 мг-экв/дм3.
Анализ численности раковинных амеб и видового состава производился прямым микроскопированием водной почвенной суспензии в чашках Петри в определенном количестве полей зрения (Гельцер и др. 1985: 79; Гельцер 1980: 154-165). Почвенные образцы грунта, доведенные до воздушно-сухого состояния, распределялись тонким слоем на ровной поверхности, отбирались крупные корешки и мелкие корешки растений. Водная суспензия приготавливалась из 200 мг почвы и 20 мл дистиллированной воды. Взвесь отстаивали в те- чение суток, надосадочную прозрачную жидкость сливали, оставшееся количество фильтрата переносили в градуированную емкость и повторно отстаивали. Суспензию, содержащую 5 г субстрата в 10 мл воды, окрашивали раствором эритрозина в течение суток. Для микроскопирования 2 мл фильтрата помещали в чашку Петри. Фильтрат разбавляли водой до объема, удобного для микроскопирования и равномерно распределяли по дну чашки. Под микроскопом БИОМЕД-2 при увеличении х160 по полям зрения просматривали суспензию. Определяли видовой состав раковинных амеб, количество живых тестаций и пустых раковинок в двукратной повторности. В каждой пробе подсчитано не менее 150 экземпляров. Полученные значения численности раковинок пересчитывали на 1 г сухого субстрата. Влажность определяли весовым методом (Соколов, Аскинази 1965). Виды амеб определяли при помощи руководств (Bonnet, Thomas 1960; Lechowicz М., Bell. 1991; Ettema, Wardle 2002; Bonnet 1975: 378).
Результаты и обсуждения
Видовой спектр раковинных амеб в зависимости от разной концентрации сеноманских вод неоднороден. Основное количество видов раковинных амеб в лабораторных исследованиях составляют представители семейств Phryganella — до четырех видов, остальные семейства
Раковинки обнаруженных видов раковинных амеб относятся к четырем морфологическим типам, имеющим как однокамерное строение раковинки, так и двухкамерное, обуславливающее дополнительную изоляцию цитоплазмы относительно внешней среды. К ак-ростомному типу относятся раковинные амебы с терминально расположенным устьем при осевой симметрии с возможным латеральным сжатием: Euglypha anodonta, Nebela dentistoma, Nebela militaris, Nebela lagenijbrmis, Nebela colla-ris. Плагиостомный тип характеризуется эксцентричным расположением раковинки на вентральной стороне и полость не разделена на брюшко и козырек: Chlamydohprys minor, Cryptodifflugia compressa, Centropyxis vandeli. К циклостомному типу относятся Phryganella acropodia, Cyclopyxis aroelloides, имеющие сферическую или полусферическую форму с уплощенной центральной поверхностью и центрально расположенным устьем. К крипто-стомному типу относятся раковинные амебы, имеющие щелевидные, эксцентрично расположенные на вентральной подошве псевдостомы, прикрытые выступами дорзальной стенки - Plagiopyxis glyphostoma (Мазей, Цыганов 2006). Изменение видового разнообразия раковинных амеб при внесении различной концентрации сеноманских вод в период исследований представлено в таблице 1.
представлены по одному виду.
Таблица 1
Изменение видового состава раковинных амеб при внесении различной концентрации сеноманских вод в период исследований
|
Виды раковинных амеб |
Концентрации растворов, г/кг |
Особенности строения |
||||
|
0 |
50 |
100 |
200 |
Морфотип |
Раковинка |
|
|
Euglypha anodonta |
+ |
Акс* |
Однокамерная |
|||
|
Chlamydohprys minor |
+ |
+ |
+ |
Плк* |
Однокамерная |
|
|
Phryganella acropodia |
+ |
+ |
+ |
+ |
Ц* |
Однокамерная |
|
Cryptodifflugia compressa |
+ |
+ |
+ |
+ |
Плк* |
Однокамерная |
|
Cyclopyxis aroelloides |
+ |
+ |
+ |
+ |
Ц* |
Двухкамерная |
|
Nebela dentistoma |
+ |
+ |
+ |
+ |
Акс* |
Однокамерная |
|
Nebela militaris |
+ |
+ |
+ |
+ |
Акс* |
Однокамерная |
|
Nebela lageniformis |
+ |
+ |
+ |
+ |
Акс* |
Однокамерная |
|
Nebela collaris |
+ |
+ |
+ |
+ |
Акс* |
Однокамерная |
|
Centropyxis vandeli |
+ |
+ |
+ |
+ |
Плк* |
Двухкамерная |
|
Plagiopyxis glyphostoma |
+ |
Крк* |
Двухкамерная |
|||
Примечание. *Акс - акростомный сжатый; Плк - плагиостомный простой; Ц - центростомный; Крк - крип-тостомный с козырьком.
Анализ данных, представленных в таблице 1, позволяет заметить, что с увеличением концентрации сеноманских вод происходит гибель раковинных амеб. Вид Chlamydohprys minor отсутствует только при концентрации сеноманских вод 100 г на кг почвы, Euglypha anodonta и Plagiopyxis glyphostoma наблюдаются только в контрольной пробе, при внесении
загрязнителя не встречаются, что свидетельст
органическим загрязнениям двух видов рако
вует о низкой устойчивости к химическим не винных амеб.
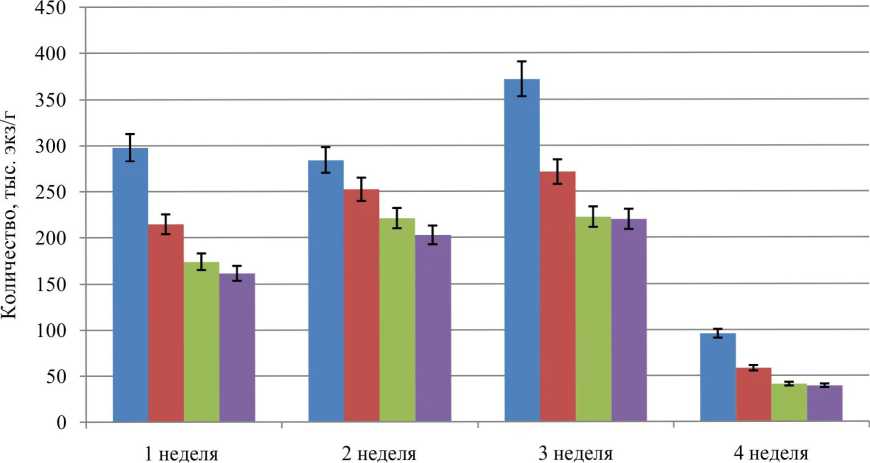
■ 1
■ 2
■ 3
■ 4
Продолжительность эксперимента
Рис. 1. Изменение общей численности раковинных амеб в зависимости от концентрации внесения сеноманских вод: 1 - контроль; 2 - при концентрации 50 г сеноманских вод на 1 кг почвы, 3 - 100 г на 1 кг почвы; 4 - 200 г на 1 кг почвы
Таблица 2
Изменение численности раковинных амеб при концентрации сеноманских растворов 50 г/кг почвы
|
Виды раковинных амеб |
Концентрация сеноманского раствора 50 г/кг |
|||
|
Длительность действия токсиканта |
||||
|
1 неделя |
2 неделя |
3 неделя |
4 неделя |
|
|
Euglypha anodonta |
8,3±0,2 0 |
0 0 |
0 0 |
0 0 |
|
Chlamydohprys minor |
3,3 ± 0,1 |
1,6 ± 0,1 |
0 |
0 0 |
|
1,5 ± 0,1 |
0 |
1,8 ± 0,04 |
||
|
Phryganella acropodia |
76,9 ± 1,6 |
41,9 ± 0,6 |
46,1 ± 3,2 |
8,7 ± 0,4 |
|
53,9 + 1,9 |
35,5 ± 2,6 |
32,2 ± 1,3 |
6,6 ± 0,7 |
|
|
Cryptodifflugia compressa |
5,1 ± 0,8 |
27,4 ± 1,3 |
34 ± 2 |
9,5 ± 0,4 |
|
0 |
25,3 ± 1,6 |
20,8 + 1,9 |
5,3 ± 0,2 |
|
|
Cyclopyxis aroelloides |
41,8 ± 0,9 |
32,2 ± 1 |
53,4 ± 1,2 |
14,8 ± 0,4 |
|
34,9 ± 1,6 |
27 ± 1,9 |
34,1 ± 2,7 |
11,3 ± 1,3 |
|
|
Nebela dentistoma |
48,4 ± 1,6 |
48,5 ± 0,8 |
70,4 ± 6 |
18,3 ± 0,5 |
|
38,1 ± 1,1 |
45,6 + 1,3 |
53 ± 3,7 |
10,6 ±0,4 |
|
|
Nebela militaris |
41,9 ± 2,6 |
33,8 ± 1,1 |
48,6 ± 1,5 |
11,3 ± 1,3 |
|
31,7 ± 4 |
32,1 ± 1,6 |
34,1 ± 2,2 |
6,6 ± 0,2 |
|
|
Nebela lageniformis |
41,8 ± 1 |
40,3 ± 1,1 |
51 ± 1,2 |
15,6 ± 1,5 |
|
33,3 ± 1,6 |
35,5 ± 1,7 |
47,4 ± 0,8 |
8,6 ± 0,2 |
|
|
Nebela collaris |
25 ± 1,5 |
48,4 ± 0,7 |
60,7 ± 1,3 |
16,5 ± 1,1 |
|
17,4 + 2,3 |
45,6 ± 1,3 |
43,6 ± 2 |
8,6 ± 0,1 |
|
|
Centropyxis vandeli |
5 ± 0,05 |
6,4 ± 0,1 |
7,2 ± 0,5 |
0,8 ±0,01 |
|
3,1 ± 0,03 |
5 ± 0,9 |
3,7 ± 0,6 |
0 |
|
|
Plagiopyxis glyphostoma |
0 0 |
3,2 ± 0,1 0 |
0 0 |
0 0 |
Примечание. В числителе дроби - среднестатистическая численность вида в контроле, в знаменателе - в опыте.
Анализ среднестатистических результатов, представленных на рисунке 1, позволяет считать, что увеличение концентрации токсических веществ коррелирует со снижением общей численности раковинных амеб. Максимальная общая численность тестаций наблюдается в третью неделю исследования в контрольной пробе. Минимальная численность амеб отмечается в четвертую неделю исследований при концентрации сеноманских вод 100— 200 г на 1 кг почвы. Снижение общей численности на 50% относительно контрольных значений свидетельствует о критическом уровне выживания сообществ раковинных амеб при действии сеноманских растворов.
Таблица 3
Изменение численности раковинных амеб при концентрации сеноманских растворов 100 г/кг почвы
|
Виды раковинных амеб |
Концентрация растворов 100 г/кг почвы |
|||
|
Длительность влияния сеноманских растворов |
||||
|
1 неделя |
2 неделя |
3 неделя |
4 неделя |
|
|
Euglypha anodonta |
8,3 ± 0,2 0 |
0 0 |
0 0 |
0 0 |
|
Chlamydohprys minor |
3,3 ± 0,1 0 |
1,6 ±0,1 0 |
0 0 |
0 0 |
|
Phryganella acropodia |
76,9 ± 1,6 |
41,9 ± 0,6 |
46,1 ± 3,2 |
8,7 ± 0,4 |
|
39,9 ± 0,6 |
29,6 ± 0,7 |
27,9 ± 1,2 |
3,8 ± 0,1 |
|
|
Cryptodifflugia compressa |
5,1 ± 0,8 |
27,4 ± 1,3 |
34 ± 2 |
9,5 ± 0,4 |
|
9,9 ± 1,3 |
19,1 ± 1 |
20,5 ± 1 |
4,3 ± 0,1 |
|
|
Cyclopyxis aroelloides |
41,8 ± 0,9 |
32,2 ± 1 |
53,4 ± 1,2 |
14,8 ± 0,3 |
|
27 ± 1,4 |
24,3 ± 1 |
20,6 ± 1,1 |
8,2 ± 0,3 |
|
|
Nebela dentistoma |
48,4 ± 1,6 |
48,5 ± 0,8 |
70,4 ± 6 |
18,3 ± 0,5 |
|
45,6 ± 1,6 |
41,6 ± 1,6 |
46,6 ± 1,4 |
7,6 ± 0,2 |
|
|
Nebela militaris |
41,9 ± 2,6 |
33,8 ± 1,1 |
48,6 ± 1,5 |
11,3 ± 1,3 |
|
5,7 ± 0,4 |
29,5 ± 1,2 |
29,8 ± 0,7 |
4,9 ± 0,1 |
|
|
Nebela lageniformis |
41,8 ± 1 |
40,3 ± 1,1 |
51 ± 1,2 |
15,6 ± 1,5 |
|
19,9 ± 0,8 |
29,5 ± 1,3 |
39,1 ± 1 |
5,4 ± 0,2 |
|
|
Nebela collaris |
25 ± 1,5 |
48,4 ± 0,7 |
60,7 ± 1,3 |
16,5 ± 1,1 |
|
25,6 ± 1,3 |
43,4 ± 1,5 |
35,4 ± 1 |
6 ± 0,09 |
|
|
Centropyxis vandeli |
5 ± 0,05 |
6,4 ± 0,1 |
7,2 ± 0,5 |
0,8 ± 0,01 |
|
0 |
3,4 ± 0,08 |
1,8 ± 0,06 |
0 |
|
|
Plagiopyxis glyphostoma |
0 0 |
3,2 ± 0,1 0 |
0 0 |
0 0 |
Примечание. В числителе дроби - среднестатистическая численность вида раковинных амеб в контроле, в знаменателе - в опыте.
Представленные в таблице 3 результаты показывают замещение по степени устойчивости к раствору вида Phryganella acropodia, наблюдаемого в первую неделю исследования, видом Nebela dentistoma с максимальной численностью 46,6±1,4тыс. экз/г абсолютно сухой почвы. Минимальная численность отмечается у вида Centropyxis vandeli и составляет 1,8±0,06 тыс. экз/г абсолютно сухой почвы, в контроль
Анализ представленных в таблице 2 данных позволяет заметить, что при исследуемой концентрации сеномана происходит снижение численности сообществ раковинных амеб. Максимальная численность наблюдается у вида Phryganella acropodia в первую неделю исследований и составляет 53,9±1,9 тыс. экз/г абсолютно сухой почвы, что в 1,4 раза меньше по сравнению с контролем. Минимальная численность наблюдается у вида Chlamydohprys minor и составляет 1,5±0,1 тыс. экз/г абсолютно сухой почвы, в то время как в контрольной кювете численность превышает в 2 раза. Виды Euglypha anodonta, Plagiopyxis glyphostoma при действии сеноманских вод с концентрацией 50 г/кг элиминируются.
ной серии наблюдается превышение в 4 раза. Вид Chlamydohprys minor встречался в лабораторных кюветах с концентрацией 50 г/кг и элиминируется при концентрации сеноманских растворов 100 г/кг. Виды Euglypha anodonta, Plagiopyxis glyphostoma не встречаются.
Анализ представленных в таблице 4 данных позволяет заметить, что доминантный вид Nebela dentistoma сохраняет свое положение и характеризуется численностью, равной 44,7±1 тыс. экз/г абсолютно сухой почвы в третью неделю исследований. Минимальная численность наблюдается у вида Chlamydohprys minor в первую неделю исследования и составляет 1,7±0,4 тыс. экз/г абсолютно сухой почвы и по сравнению с контрольной серией снижается в 1,9 раз. Вид встречается только в первую неделю исследования. Виды Plagiopyxis glyphostoma, Euglypha anodonta наблюдаются только в контрольных кюветах.
Таблица 4
Изменение численности раковинных амеб при концентрации сеноманских растворов 200 г/кг почвы
|
Виды раковинных амеб |
Концентрация растворов 200 г/кг |
|||
|
Длительность влияния сеноманских растворов |
||||
|
1 неделя |
2 неделя |
3 неделя |
4 неделя |
|
|
Euglypha anodonta |
8,3 ± 0,2 0 |
0 0 |
0 0 |
0 0 |
|
Chlamydohprys minor |
3,3 ± 0,1 |
1,6 ±0,1 |
0 0 |
0 0 |
|
1,7 ± 0,4 |
0 |
|||
|
Phryganella acropodia |
76,9 ± 1,6 |
41,9 ± 0,6 |
46,1 ± 3,2 |
8,7 ± 0,4 |
|
37,4 ± 1,2 |
25 ± 1,1 |
29,1 ± 1,4 |
3,8 ± 0,7 |
|
|
Cryptodifflugia compressa |
5 ± 0,8 |
27,4 ± 1,3 |
34 ± 2 |
9,5 ± 0,4 |
|
10,2 ± 0,7 |
19,2 ± 1,9 |
20,1 ± 0,6 |
2,2 ± 0,1 |
|
|
Cyclopyxis aroelloides |
41,8 ± 0,9 |
32,2 ± 1 |
53,4 ± 1,2 |
14,8 ± 0,3 |
|
30,6 ± 1,6 |
17,3 ± 0,8 |
22,3 ± 1,4 |
7,6 ± 0,2 |
|
|
Nebela dentistoma |
48,4 ± 1,6 |
48,5 ± 0,8 |
70,4 ± 6 |
18,3 ± 0,5 |
|
25,5 ± 0,6 |
38,5 ± 1,2 |
44,7 ± 1 |
8,3 ± 0,1 |
|
|
Nebela militaris |
41,9 ±2,6 |
33,8 ± 1,1 |
48,6 ± 1,5 |
11,3 ± 1,3 |
|
13,6 ± 1,8 |
26,9 ± 0,3 |
33,5 ± 2,5 |
4,5 ± 0,1 |
|
|
Nebela lageniformis |
41,8 ± 1 |
40,3 ± 1,1 |
51 ± 1,2 |
15,6 ± 1,5 |
|
18,7 ± 1,1 |
28,9 ± 0,9 |
33,7 ± 1,7 |
5,3 ± 0,2 |
|
|
Nebela collaris |
25 ± 1,5 |
48,4 ± 0,7 |
60,7 ± 1,3 |
16,5 ± 1,1 |
|
23,8 ± 1,3 |
44,3 ± 0,8 |
33,5 ± 1,7 |
6,8 ± 0,1 |
|
|
Centropyxis vandeli |
5 ± 0,05 |
6,4 ±0,1 |
7,2 ± 0,5 |
0,8 ± 0,01 |
|
0 |
1,9 ± 0,04 |
2,2 ± 0,01 |
0 |
|
|
Plagiopyxis glyphostoma |
0 0 |
3,2 ± 0,1 0 |
0 0 |
0 0 |
Примечание. В числителе дроби - среднестатистическая численность вида раковинных амеб в контроле, в знаменателе - в опыте.
Таким образом, на основании проведенных исследований можно сделать следующие выводы. Выявлены устойчивые к сеноманским водам виды раковинных амеб: Phryganella acropodial, Cryptodifflugia compressa, Cyclopyxis aroelloides, Nebela dentistoma, Nebela militaris, Nebela lageniformis, Nebela collaris.
Показано снижение видового разнообразия раковинных амеб при концентрации сеноманских вод (50 г/кг), проявляющееся в элиминации Euglypha anodonta, Plagiopyxis glyphostoma.
При концентрации сеноманских растворов, равных 100 г/кг, установлена элиминация видов: Euglypha anodonta, Plagiopyxis glyphostoma, Chlamydohprys minor и снижение-численности устойчивых видов амеб.
При концентрации 200 г/кг происходит снижение видового разнообразия сообщества раковинных амеб, снижение численности в опытной группе больше 50%, что является критическим уровнем выживания популяций раковинных амеб.
Список литературы Влияние сеноманских вод на сообщества раковинных амеб
- Биоиндикация загрязнения наземных экосистем. 1988 / Шуберт Р. (ред.). М.: Мир.
- Борисович Т. В. 1985. Влияние нефтяного загрязнения на почвенных простейших // Матер. докл. IX Междунар. коллоквиума по почвенной зоологии. Вильнюс.
- Гельцер Ю. Г. 1980. Методы изучения почвенных простейших // Почвенные простейшие. Серия Протозоология. Вып. 5. Л.: Наука, 154-165.
- Гельцер Ю. Г., Корганова Г. А., Алексеев Д. А. 1985. Почвенные раковинные амебы и методы их изучения. М.: Наука.
- Залялетдинова Н. А., Карташев А. Г. 2016. Влияние экологических факторов на сообщества почвенных инфузорий. Томск: ТУСУР.
- Карташев А. Г. 2007. Экологические аспекты нефтедобывающей отрасли Западной Сибири. Томск: ТУСУР.
- Карташев А. Г., Смолина Т. В. 2011. Влияние нефтезагрязнений на почвенных беспозвоночных животных. Томск: В-Спектр.
- Мазей Ю.А., Цыганов А. Н. 2006. Пресноводные раковинные амебы. Москва: Товарищество научных изданий КМК.
- Соколов А. В., Аскинази Д. Л. 1965. Агротехнические методы исследования почв. М.: Наука.
- Сулин В.А. 1935. Воды нефтяных месторождений СССР. М.; Л.: ОНТИ НКТП.
- Шмараев, А.В., Шорина Т. С. 2009. Влияние нефти и нефтепродуктов на различные компоненты окружающей среды // Вестник Оренбургского государственного университета. 6 (112), 642-645.
- Bonnet L. 1975. Types morphologiques, ecologie et evolution de la thegue chez les Thecamoebiens // Protistologica 11/3, 303-378.
- Bonnet L., Thomas R. 1960. Thécamoebiens du sol // Vie et Milieu. Suppl. 5, 1-113.
- Ettema Ch., Wardle D. 2002. Spatial soil ecology // Trends in ecology and evolution. 17, 177-183.
- Lechowicz M., Bell G. 1991. The ecology and genetics of fitness of forest plants. II. Microspatial heterogeneity of the edaphic environment // Journal of Ecology. Vol. 79, 687-696.
