Влияние совместного действия постоянного освещения и хронической алкогольной интоксикации на ультраструктуру гепатоцитов самцов и самок крыс «Вистар»

Автор: Арешидзе Д.А., Михалва Л.М., Кактурский Л.В., Кондашевская М.В., Козлова М.А., Черников В.П.
Журнал: Ульяновский медико-биологический журнал @medbio-ulsu
Рубрика: Биологические науки
Статья в выпуске: 3, 2023 года.
Бесплатный доступ
Световое загрязнение, вызывающее дефицит мелатонина и нарушение циркадианной ритмичности, ассоциировано с развитием ряда патологий печени. Рассогласование суточной ритмики приводит к повышению восприимчивости печени к вызываемым алкоголем повреждениям, усилению тяжести алкогольной болезни. Целью исследования было изучение сочетанного действия постоянного освещения и хронической алкогольной интоксикации (ХАИ) на ультраструктуру гепатоцитов крыс обоих полов. Материалы и методы. Исследование проведено на 120 самцах и 80 самках крыс аутбредного стока «Вистар» в возрасте 6 мес. Длительность эксперимента составляла 3 нед. Визуально оценивали различия в морфологической картине ультраструктур гепатоцитов, количественно определяли процент гепатоцитов, содержащих липидные вакуоли. Результаты. Постоянное освещение и хроническая алкогольная интоксикация вызывали формирование состояния аллостаза. В клетках печени крыс обоих полов были выявлены отек гепатоцитов, набухание митохондрий, сморщивание ядра, развитие жировой дистрофии, гибель гепатоцитов. Половые различия заключались в меньшей выраженности указанных патологических изменений у самок. В сочетании с отсутствием у крыс женского пола разрастания коллагеновых прослоек в тканях печени перечисленные данные свидетельствуют о том, что в печени самок, в отличие от печени самцов, сочетанное воздействие постоянного освещения и ХАИ не приводит к развитию изменений воспалительного характера и нарушению архитектоники. С учетом ранее охарактеризованной нами морфологической картины печени крыс обоих полов и ряда биохимических параметров, описывающих ее функциональное состояние, полученные данные о межполовых различиях в ультраструктурных характеристиках гепатоцитов крыс, подверженных воздействию ХАИ и постоянного освещения, позволяют нам высказать утверждение о более успешном протекании адаптации гепатоцитов к стрессорным условиям у самок крыс, нежели у самцов.
Реакция гепатоцитов самок и самцов крыс «вистар» на сочетанное действие постоянного освещения и потребление алкоголя, мелатонин, электронная микроскопия
Короткий адрес: https://sciup.org/14128737
IDR: 14128737 | УДК: 616-091.8 | DOI: 10.34014/2227-1848-2023-3-151-166
Influence of combined effect of constant illumination and chronic alcohol intoxication on hepatocyte ultrastructure in male and female Wistar rats
Light pollution causes melatonin deficiency and circadian rhythm disruption. Thus, it is associated with hepatopathy. Circadian rhythm disruption increases liver damage from alcohol and the severity of alcoholism. The aim of the study was to examine the combined effect of constant illumination and chronic alcohol intoxication (CAI) on hepatocyte ultrastructure in male and female rats. Materials and Methods. The study was carried out on 120 male and 80 female Wistar outbred rats aged 6 months. The experiment lasted 3 weeks. Differences in morphological patterns of hepatocyte ultrastructure were visually assessed. The percentage of hepatocytes containing lipid vacuoles was calculated. Results. Constant illumination and chronic alcohol intoxication caused allostasis. Hepatocyte edema, mitochondria swelling, karyopyknosis, fatty degeneration, and hepatocyte death were detected in the liver cells of male and female rats. The above-mentioned pathological changes were less severe in females. Moreover, collagen layers did not proliferate into the liver tissues in female rats. So, we can conclude that the combined effect of constant illumination and CAI do not lead to inflammatory changes and architectural distortion in the liver of female rats. Taking into account the morphological picture of the liver of male and female rats and a number of biochemical parameters that describe its functional state, the obtained data on gender differences in the hepatocyte ultrastructure in rats exposed to CAI and constant illumination allow us to report that hepatocyte adaptation to stress is more successful in female rats than in male ones.
Текст научной статьи Влияние совместного действия постоянного освещения и хронической алкогольной интоксикации на ультраструктуру гепатоцитов самцов и самок крыс «Вистар»
Введение. Жизнедеятельность организмов, функционирование органов и систем, обмен веществ, передача энергии и информации в живых системах подчиняются закону биологической структурно-временной интеграции. К настоящему времени в организме млекопитающих охарактеризовано более 500 различных функций и процессов, протекающих в зависимости от циркадианных (околосуточных) ритмов [1].
Циркадианные ритмы (ЦР) - сформировавшиеся в ходе эволюции и генетически обусловленные колебания интенсивности протекания биологических функций и процессов в организме с периодом 24±4 ч. У живых организмов ЦР с различными амплитудно-фазовыми характеристиками строго согласованы между собой и с факторами внешней и внутренней среды, благодаря чему достигается поддержание функционирования систем организма на оптимальном уровне [2, 3].
К наиболее значимым факторам дезорганизации биоритмов в современном мире относят различные нарушения светового режима, в частности световое загрязнение – воздействие света в ночное время [4]. Световое загрязнение, вызывающее десинхроноз, у человека обусловлено рядом социальных причин: продолжительным взаимодействием с цифровой техникой, сверхурочной и сменной работой, трансмеридианными перелетами (jetlag) и т.д. [5]. Согласно принятой большинством хронобиологов гипотезе циркадианной деструкции воздействие света в ночные часы является стрессорным фактором, который нарушает эндогенный циркадианный ритм и препятствует нормальной синхронизации эндогенных ритмов с ритмами смены света и темноты, а также препятствует синтезу эпифизарного мелатонина как у человека, так и у животных [6].
Мелатонин – физиологически активное вещество, основной гормон эпифиза, один из основных модуляторов циркадианного ритма у млекопитающих, в т.ч. человека, синхронизирующий эндогенные циркадианные ритмы организма, его физиологию и поведение с окружающей средой. Вопреки устоявшемуся мнению данный гормон не является только лишь модератором цикла «сон – бодрствование»: системный гормональный ответ, в котором принимает участие мелатонин, состоит в координации и синхронизации нейроиммуно-физиологических процессов и поведенческой адаптации к геофизическому дню и переменам в окружающей среде. С помощью суточных циклов синтеза мелатонина в организме согласовываются адаптационные физиологические процессы в темное время суток (в случае как сна, так и бодрствования, в зависимости от видовых паттернов суточной активности) и создаются предпосылки для процессов адаптации, протекающих в дневное время, когда выработка гормона супрессируется воздействием света [7].
Снижение содержания мелатонина вызывает множество структурно-функциональных отклонений, провоцирует ускоренное старение, является одной из причин развития забо- леваний органов желудочно-кишечного тракта, сердечно-сосудистой системы и др. [8, 9].
Установлено, что световое загрязнение, вызывающее дефицит мелатонина и нарушение циркадианной ритмичности, ассоциировано с развитием ряда патологий печени, включая неалкогольную жировую болезнь, злокачественные новообразования, билиарный цирроз и др. [10, 11].
Помимо непосредственного деструктивного влияния светового загрязнения на морфофункциональное состояние печени, немалое значение имеет также тот факт, что люди, вынужденные постоянно пребывать в агрессивных условиях неблагоприятного светового режима (например, при работе в ночное время), часто демонстрируют повышенную склонность к употреблению кофе, алкоголя или алкоголесодержащих энергетических напитков [12, 13]. Как известно, злоупотребление алкоголем составляет одну из наиболее важных медико-социальных проблем как в нашей стране, так и в современном мире в целом [14].
Алкоголь, являясь высокоактивным стрессором, оказывает прямое и опосредованное деструктивное влияние как на структуру суточных ритмов организма в целом, так и на ЦР отдельных органов, в частности печени, что может проявиться как на уровне часовых генов, так и на иерархически более высоких уровнях системы регуляции циркадианной ритмичности организма млекопитающих [15].
В частности, у людей, страдающих алкоголизмом, выявлено нарушение нормальной экспрессии основных часовых генов Clock, Bmal1, Per1, Per2, Cry1 и Cry2 , приводящее к нарастающей десинхронизации гомеостатических процессов в организме [16, 17], что принято связывать с обусловленным алкоголем окислительным стрессом.
Исследования метаболизма мелатонина у больных алкоголизмом показали, что у пациентов с алкогольной зависимостью отмечается резкое снижение концентрации этого гормона в сыворотке крови в ночное время суток. Также сообщается об инверсии циркадианного ритма продукции мелатонина у больных алкоголиз- мом как в период алкоголизации, так и в первые дни отмены этанола. Показано, что хроническая алкогольная интоксикация (ХАИ) истощает тканевые ресурсы эпифиза и приводит к тяжелым нарушениям его функций, что в свою очередь способствует возникновению более тяжелых форм алкоголизма [18].
Рассогласование ЦР в свою очередь приводит к повышению восприимчивости органов желудочно-кишечного тракта (ЖКТ) и печени к вызываемым алкоголем повреждениям, усилению тяжести алкогольной болезни. Таким образом, совместное действие светового загрязнения и алкоголизации создает порочный круг взаимоусугубляющих нарушений функционирования печени [19].
К настоящему времени хорошо известно, что циркадианной ритмичности практически всех физиологических процессов присущи половые различия [20, 21]. Гендерные особенности циркадной ритмичности описаны как для людей, так и для животных, являющихся моделью для исследования суточных ритмов. Необходимо отметить, что исследования, проведенные на лабораторных грызунах, могут быть экстраполированы на человека [22]. Кроме того, сочетанное воздействие светового загрязнения и алкоголя не означает простого сложения влияния этих стрессорных факторов, но может носить гораздо более интенсивный патогенный характер. Однако при этом, несмотря на высокую социальную значимость проблемы, вопрос о гендерных различиях морфофункционального состояния печени при действии этих факторов остается малоизученным.
Цель исследования. Изучение сочетанного действия постоянного освещения и ХАИ на ультраструктуру гепатоцитов крыс обоих полов .
Материалы и методы. Работа выполнена на 120 самцах и 80 самках крыс аутбредного стока «Вистар» в возрасте 6 мес. и с массой тела 350±15 г. Животные были получены из питомника ФГБУН НЦБМТ ФМБА России «Столбовая». Все крысы содержались в пластиковых клетках по 10 особей. Первоначально животных содержали при естественном освещении, температуре 20–22 °С и относительной влажности воздуха 60–70 %. Крысы имели свободный доступ к питьевой воде и стандартному брикетированному корму ПК-120-1 (ООО «Лабораторснаб», сертификат соответствия № POCCRU.nO81.B00113, ГОСТ P50258–92). Все экспериментальные процедуры проводились в соответствии с Директивой Европейского парламента 2010/63/ЕС «О защите животных, используемых в экспериментальных целях» (от 22.09.2010). На проведение исследования получено разрешение биоэтического комитета ФГБНУ НИИМЧ им. А.П. Авцына (протокол № 34 (10) от 14.03.2022).
Крысы были случайным образом разделены на 4 равные группы.
I группа – самцы (контроль, n=60), содержавшиеся при фиксированном световом режиме (свет/темнота – 10/14 ч с включением света в 8:00 и выключением в 18:00).
II группа – самки (контроль, n=40), также содержавшиеся при фиксированном световом режиме (свет/темнота – 10/14 ч с включением света в 8:00 и выключением в 18:00).
-
III группа – самцы (n=60), содержавшиеся при постоянном освещении и получавшие в качестве питья 15 % раствор этанола ad libitum вместо воды, т.е. подвергавшиеся хронической алкогольной интоксикации (свет + ХАИ).
-
IV группа – самки (n=40), также содержавшиеся при постоянном освещении и получавшие в качестве питья 15 % раствор этанола ad libitum (свет + ХАИ) .
Отбор самок крыс в экспериментальные группы осуществлялся после определения фазы эстрального цикла по кольпоцитограмме, чтобы в эксперимент вошли животные в фазе диэструса.
Критерием отбора крыс в исследование, наряду с отсутствием видимых отклонений поведения и внешнего вида животного (состояние шерстного покрова, глаз, конечностей), было исходное предпочтение алкоголя (15 % раствор этилового спирта) водопроводной воде. Для этого был проведен предварительный эксперимент: в течение 3 сут крыс содержали в индивидуальных клетках со свободным доступом к обеим жидкостям.
В течение эксперимента ежедневно определяли объем выпитого раствора этанола, за- тем вычисляли массу спирта на 1 кг массы тела. В среднем животные обоего пола выпивали 15,48±1,28 мл/сут, что в пересчете на абсолютный этанол составляет 7 г/кг массы тела.
Длительность эксперимента составляла 3 нед.
Выведение крыс из эксперимента осуществляли в углекислотной камере, оборудованной устройством для верхней подачи газа (100 % СО 2 ) в 9:00, 15:00, 21:00 и 3:00. Заполнение объема камеры газом осуществляли со скоростью 20 % в минуту во избежание возникновения у животных диспноэ и боли. Под действием газа животные засыпали, после чего проводилась эвисцерация печени.
Образцы печени размером 2 мм3 фиксировали 2,5 % раствором глутарового альдегида на фосфатном буфере (рН 7,4), дофиксировали в 1 % растворе четырехокиси осмия (OsO 4 ), обезвоживали в этаноле по общепринятой схеме, в процессе обезвоживания контрастировали 1 % уранилацетатом на 70 % этаноле и проводили заливку в смесь эпон-аралдит по стандартной методике [23]. Ультратонкие срезы получали на ультратоме LKB-III (LKB Produkter, Швеция), срезы дополнительно контрастировали цитратом свинца по Рейнольдсу и просматривали в просвечивающем электронном микроскопе JEM-100CX (JEOL, Япония). Фотофиксацию препаратов осуществляли с помощью камеры Gatan ES500W Erlangshen, Model 782 (Gatan Inc., США).
На полученных препаратах визуально оценивали различия в морфологической картине ультраструктур гепатоцитов, количественно определяли процент гепатоцитов, содержащих липидные вакуоли.
Результаты. Ранее было продемонстрировано, что морфофункциональное состояние гепатоцитов крыс «Вистар», находившихся под воздействием ХАИ и постоянного освещения, имеет ряд существенных гендерных различий. Наши предыдущие исследования показали, что постоянное освещение в сочетании с ХАИ оказывает существенное влияние на морфофункциональную целостность печени крыс-самцов, что проявляется в развитии у них жировой дистрофии и стеатогепатита с появлением у некоторых животных признаков цирроза печени. У самок в тех же условиях мы отмечали только умеренно выраженную жировую дистрофию. При этом нами была показана повышенная экспрессия Ki-67 и р53 у животных обоих полов, что свидетельствует об усилении процессов пролиферации и апоптоза в печени крыс всех экспериментальных групп, при этом интенсивность данных процессов была выше у самок [24, 25].
Результаты электронно-микроскопического исследования гепатоцитов крыс, подвергавшихся совместному воздействию ХАИ и постоянного освещения, подтверждают и более подробно иллюстрируют вышеописанные гендерные различия в морфофункциональном состоянии печени.
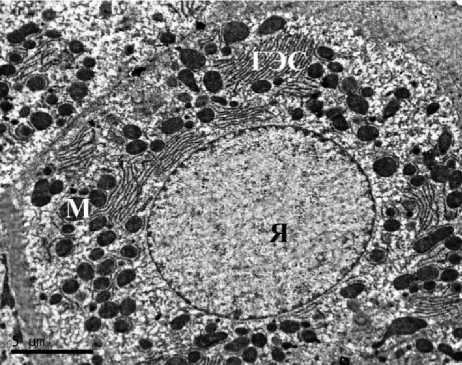
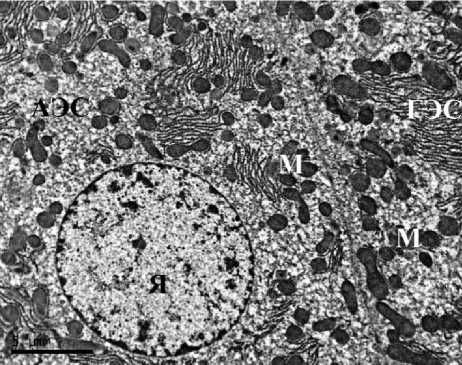
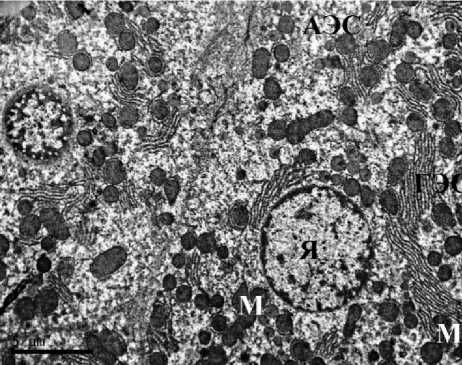
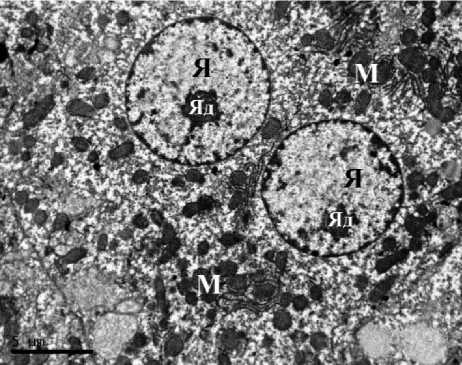
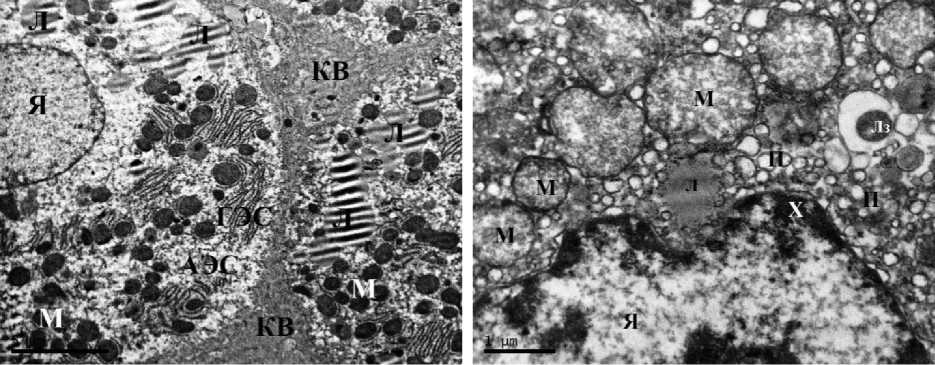
Ультраструктурная картина гепатоцитов животных контрольных групп соответствовала норме как у самцов, так и у самок. В гепатоцитах полигональной формы наблюдались округлые ядра, расположенные по центру клетки, в цитоплазме визуализировались зерна гликогена, митохондрии содержали плотный матрикс, развитые эндоплазматические сети не имели признаков вакуолизации. Таким образом, гепатоциты животных групп контроля не имели признаков патологических изменений и некроза. В строме печени визуализировались единичные тонкие ретикулярные волокна (рис. 1).
Б / B
В / C
Рис. 1. Ультраструктура гепатоцитов крыс: А, Б – I группы (контроль, самцы);
В, Г – II группы (контроль, самки). ТЭМ, ×6700 (АЭС – агранулярная эндоплазматическая сеть, ГЭС – гранулярная эндоплазматическая сеть, М – митохондрия, Я – ядро, Яд – ядрышко)
Г / D
Fig. 1. Ultrastructure of rat hepatocytes: A, B – Group 1 (control, males); C, D – Group 2 (control, females). TEM, ×6700. Symbols: АЭС – agranular endoplasmic reticulum; ГЭС – granular endoplasmic reticulum;
M – mitochondrion; Я – nucleus; Яд – nucleolus
Состояние гепатоцитов животных экспериментальных групп на ультраструктурном уровне заметно отличалось от такового у крыс контрольных групп. При наличии сходной картины ряда адаптационных и патологических изменений у самцов и самок степень выраженности данных явлений значительно разнилась.
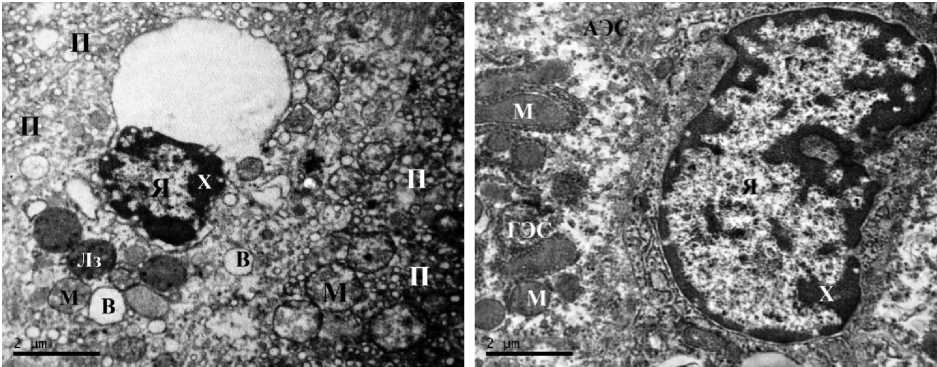
Методом трансмиссионной электронной микроскопии (ТЭМ) удалось обнаружить в гепатоцитах самцов крыс, подвергнутых сочетанному действию света и ХАИ, значительное количество пикнотичных ядер, что сопровождалось конденсацией ядерного хроматина, аккумуляцией пероксисом, а также выраженной гиперплазией митохондрий. Гранулярный эн- доплазматический ретикулум характеризовался выраженной везикуляцией и дилатацией цистерн; в цитоплазме гепатоцитов отмечалось увеличение количества вакуолей, в т.ч. липидосодержащих. Локально наблюдалось набухание митохондрий, сопровождаемое лизисом их крист и матрикса и наличием неровности наружной мембраны. В образцах печени самцов крыс, подвергавшихся сочетанному воздействию постоянного освещения и ХАИ, при использовании методов ТЭМ хорошо визуализировались как одиночные некротизированные гепатоциты, так и очаги микронекроза, а также клетки печени в состоянии апоптоза (рис. 2 А–В).
Кроме вышеописанных изменений, характерных в той или иной степени для всех самцов группы сочетанного воздействия постоянного освещения и ХАИ, у части особей отмечалось образование в толще паренхимы узких прослоек соединительной ткани, представленных разрастаниями коллагеновых волокон, обладающих характерной исчер-ченностью. Это наблюдение согласуется с вышеупомянутыми результатами гистологических исследований, в которых мы показали развитие печеночного фиброза у 13,3 % (n=8) самцов крыс, подвергавшихся одновремен- ному воздействию постоянного освещения и ХАИ.
Отмечались признаки репаративной регенерации. Так, новообразовавшиеся в результате репаративного митоза гепатоциты характеризовались округлой формой клеток с крупными светлоокрашенными ядрами, дилатацией эндоплазматического ретикулума, мелкими митохондриями, присутствием липидных глобул, дефицитом гликогена в цитоплазме, невозможностью идентифицировать комплекс Гольджи; также наблюдалось расширение межклеточных контактов (рис. 2 Г).
А / A Б / B
В / C
Г / D
Рис. 2. Ультраструктура гепатоцитов III группы (свет + ХАИ): А – ×8000; Б, В – ×14 000;
Г – ×20 000 (АЭС – агранулярная эндоплазматическая сеть, ГЭС – гранулярная эндоплазматическая сеть, В – вакуоль, КВ – коллагеновые волокна, Л – липиды, Лз – лизосома, М – митохондрия,
П – пероксисомы, Х – хроматин, Я – ядро)
Fig. 2. Ultrastructure of Group 3 hepatocytes (light + chronic alcohol intoxication): A – ×8000;
B, C – ×14 000; D – ×20 000. Symbols: АЭС – agranular endoplasmic reticulum;
ГЭС – granular endoplasmic reticulum; В – vacuole; КВ – collagen fibers; Л – lipids; Лз – lysosome; M – mitochondrion; П – peroxisomes; X – chromatin; Я – nucleus
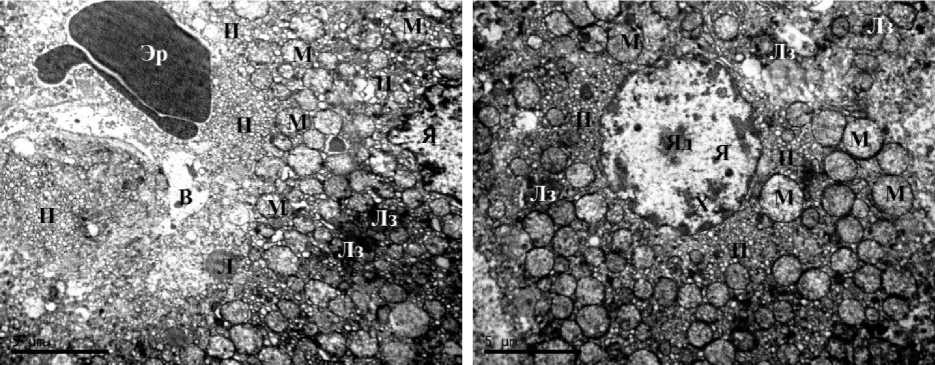
У самок крыс, содержавшихся в условиях постоянного освещения, сопровождавшегося ХАИ, отмечен ряд сходных с самцами изменений в гепатоцитах: отек и присутствие вакуолей в цитоплазме некоторых клеток, сморщивание ядер и присутствие значительного числа пероксисом. В отличие от самцов у самок наличие выраженной мелко- и крупнокапельной жировой дистрофии было отмечено только в 2 % случаев (рис. 3 А–В). Отечность, как и набухание митохондрий, встречалась только в единичных клетках, некротизированных гепатоцитов отмечено не было, однако в большинстве образцов встречались клетки в состоянии апоптоза, что успешно иллюстрирует ранее полученные нами данные о более высоком уровне апоптической гибели гепатоцитов у самок по сравнению с самцами.
Как и у самцов крыс, подвергавшихся совместному воздействию постоянного освещения и ХАИ, у самок данной группы наблюдалось значительное количество гепатоцитов, обладающих характерными признаками новообразованных клеток (рис. 3 Г). Важно отметить, что, в отличие от самцов, у самок не обнаружено разрастания коллагеновых прослоек в тканях печени.
А / A Б / B

В / C
Г / D
Рис. 3. Ультраструктура гепатоцитов крыс IV экспериментальной группы (свет + ХАИ):
А, Б – ×8000; В, Г – ×14000 (В – вакуоль, Г – комплекс Гольджи, ГЭС – гранулярная эндоплазматическая сеть, Л – липиды, Лз – лизосома, М – митохондрия, П – пероксисомы, Х – хроматин, Я – ядро, Яд – ядрышко)
Fig. 3. Ultrastructure of rat hepatocytes, Group 4 (light + chronic alcohol intoxication):
A, B – ×8000; C, D – ×14000. Symbols: B – vacuole; Г – Golgi complex;
ГЭС – granular endoplasmic reticulum; Л – lipids; Лз – lysosome; M – mitochondrion; П – peroxisomes;
X – chromatin; Я – nucleus; Яд – nucleolus
Обсуждение. Известно, что хроническая алкогольная интоксикация вызывает комплекс метаболических нарушений, осложняющих и усугубляющих токсическое действие алкоголя, что приводит к развитию морфофункциональных нарушений в тканях печени. Постоянное освещение (темновая депривация) в свою очередь является значительным стрессирующим фактором, также отражающимся на морфофункциональном состоянии гепатоцитов. Однако сочетание обоих факторов практически не рассматривалось в научной литературе. Проведенное нами изучение сочетанного воздействия исследуемых факторов дало результаты, в значительной степени отличные от результатов их отдельного действия.
Выявленные в наших экспериментах уль-траструктурные изменения в гепатоцитах крыс обоих полов, наличие обширных дистрофических проявлений, единичной и очаговой некротической гибели клеток свидетельствуют о значительной выраженности альтера-тивных процессов вследствие кумуляции токсического воздействия этанола и его метаболитов со светиндуцированными девиациями мелатонинассоциированных эффектов [26].
Хроническое употребление алкоголя приводит к окислительному стрессу за счет интенсификации метаболизма через систему цитохрома P450 2E1, индуцирующую активные формы кислорода, ацетальдегид и ДНК-бел-ковые аддукты. Они активируют воспалительные сигнальные пути в печени, что приводит к экспрессии провоспалительных медиаторов, вызывающих апоптоз и некроз гепатоцитов. Воздействие активных форм кислорода в свою очередь приводит к митохондриальному стрессу, провоцируя структурную и функциональную дизрегуляцию митохондрий и активацию апоптотических сигналов [27]. Митохондриальная гиперплазия, наблюдавшаяся у животных обоих полов в нашем эксперименте, представляет собой физиологический ответ на увеличение потребности в энергии в условиях митохондриального стресса [28].
Наблюдаемые нами явления набухания гепатоцитов, «разрежение» (дистрофия) их цитоплазмы и нарушение цитоскелета, а также появление в поле зрения некротически и апоптотически гибнущих клеток описаны ранее как проявление влияния алкоголя на печень [29], но при малой продолжительности хронического употребления алкоголя, каковым, несомненно, является 21-дневный срок, выраженность подобных изменений в печени обычно весьма незначительна [30, 31]. Однако сочетанное влияние ХАИ с постоянным освещением, изученное в данном исследовании, привело у самцов к развитию в печени картины гепатита, отягощенного фиброзом, а у самок - к дистрофическим изменениям, сопровождающимся апоптотической гибелью гепатоцитов.
Развитие в печени фиброза является исходом ее хронического повреждения (в основном воспалительного характера), спровоцированного воздействием алкоголя. Алкоголь и ацетальдегид могут напрямую активировать клетки Ито, способствуя тем самым фиброге-незу в печени, однако опосредованное алкоголем ингибирование нескольких антифиброз-ных путей также создает дополнительные условия для развития фиброза [32, 33]. При сочетанном действии хронической алкогольной интоксикации и постоянного освещения фиб-рогенез был значительно интенсифицирован. Этот факт объясняется выраженным снижением вследствие темновой депривации уровня эпифизарного мелатонина, который, как известно, обладает противофиброзным действием, существенно ограничивая потенциал клеток Ито к дифференцировке в сторону миофибробластов с дальнейшим ограничением аккумуляции фибронектина в перисину-соидальном пространстве печени [34].
Накопление липидов в печени крыс экспериментальных групп является закономерным следствием кумулятивного действия обоих изученных патогенных факторов, каждый из которых имеет свою форму влияния на жировой обмен. Известно, что воздействие этанола на гепатоциты приводит к интенсификации высвобождения липидов из жировых депо и биосинтеза жирных кислот, а также усилению синтеза и накопления холестерина [35]. Эти процессы в свою очередь находятся под модулирующим влиянием уровня эпифизарного мелатонина, выработка которого значительно снижается у лабораторных грызунов под действием постоянного освещения [36]. Кроме того, развитие мелкокапельной жировой дистрофии в гепатоцитах описано как характерное проявление хронического стресса, каковым, безусловно, является воздействие постоянного освещения для всех млекопитающих независимо от ночного или дневного видового типа активности [37].
Ранее нами было показано, что при раздельном действии алкоголя и световой нагрузки адаптация гепатоцитов к патогенному воздействию осуществляется преимущественно за счет их гипертрофии вследствие внутриклеточной регенерации. Этот вывод подтверждается данными литературы, свидетельствующими о том, что на начальных этапах адаптации к патогенному воздействию сохранение функциональных способностей печени осуществляется именно за счет внутриклеточной регенерации путем гипертрофии гепатоцитов без их пролиферации [38, 39].
Сочетанное же воздействие алкоголя и постоянного освещения оказывает значительно более выраженное влияние, при котором процесс адаптационной гипертрофии клеток, представляющий собой основной механизм срочной репарации, оказывается недостаточным, что приводит к интенсификации пролиферативного процесса [24, 25].
Размер ядра и его форма в норме и при патологических процессах зависят от целого ряда биологических факторов, влияющих на клетку, и ее состояния на момент воздействия. Как известно, деформация ядер гепатоцитов при хронической алкогольной интоксикации ассоциирована, в частности, со снижением содержания в гепатоцитах ламинов B-типа, что вызывает серьезные изменения цитоскелета гепатоцитов. Также показано, что ламины агрегируются при окислительном повреждении печени, что сопровождается изменением формы и размера ядер гепатоцитов. При этом ядерная оболочка претерпевает серьезные изменения, однако коллапс цитоскелета способствует закрытию ядерных пор, препятствуя проникновению токсинов из цитоплазмы в ядро [40].
Характерным ультраструктурным признаком гепатоцитов крыс обоих полов в нашем эксперименте было чрезвычайно выраженное увеличение числа пероксисом. Как известно, каталазы пероксисом являются одним из инструментов окисления этанола в гепатоцитах, значимость роли которого возрастает при хронической алкогольной интоксикации, приводящей к постепенному истощению резервов основного пути метаболизма алкоголя – окисления с участием алкогольдегидрогеназы [41]. Выявлена положительная связь между уровнем этанола в крови и активностью каталазно-пероксидазной системы его окисления, а также возрастание данной активности при длительном систематическом употреблении алкоголя, закономерно сопровождаемом значительным увеличением числа пероксисом. Способность пероксисомных каталаз к окислению этанола возрастает также под действием оксидативного стресса, который развивается при отсутствии антиоксидантного действия мелатонина, синтез которого угнетается у крыс в условиях постоянного освещения [42]. При этом также известно, что повышение числа пероксисом и образование ими своеобразных ретикулярных структур (peroxisomal reticular arrangements) характерно для активно регенерирующей печени [43].
Выявленные половые различия в уль-траструктурных особенностях гепатоцитов животных, заключающиеся в меньшей выраженности дистрофических проявлений и отсутствии очагов некроза гепатоцитов и фиброза у самок, можно объяснить влиянием на печень половых гормонов [44]. В частности, известно, что в гепатоцитах экспрессируются преимущественно β-рецепторы эстрогена. При нокауте гена, отвечающего за их экспрессию, у лабораторных грызунов обоих полов возрастает риск развития стеатогепатита, а его мутация у человека ассоциирована с нарушением углеводного обмена. Гепатопротектор-ное действие эстрадиола осуществляется посредством подавления процессов воспаления, развития фиброза и накопления липидов в клетках. Кроме того, эстрадиол усиливает окисление жирных кислот в печени, снижает экспрессию ряда генов липогенеза, таким образом уменьшая синтез жирных кислот [45]. Как известно, воспаление и фиброз в печени запускаются клетками Купфера, секретирующими провоспалительные цитокины, включая IL-6. Было показано, что эстрогены ингибируют секрецию IL-6, что обусловливает снижение его концентрации в плазме крови и уменьшение интенсивности воспаления в клетках печени [46, 47].
Таким образом, проведенное исследование ультраструктурных особенностей гепатоцитов самцов и самок крыс при воздействии постоянного освещения и хронической алкогольной интоксикации позволило установить, что стресс вызвал выраженные изменения гомеостаза, в результате сформировалось состояние аллостаза. Ценой этого вида адаптации является аллостатическая нагрузка, возникающая в результате постоянной или повторяющейся активации выделения гормонов стресса и ведущая к повреждению защитных механизмов организма. В клетках печени у крыс обоих полов были выявлены отек гепатоцитов, набухание митохондрий, сморщивание ядра, развитие жировой дистрофии, гибель гепатоци- тов. Половые различия заключались в меньшей выраженности указанных патологических изменений у самок. В сочетании с отсутствием у крыс женского пола разрастания коллагеновых прослоек в тканях печени перечисленные данные свидетельствует о том, что в печени самок сочетанное воздействие постоянного освещения и ХАИ не приводит к развитию изменений воспалительного характера и нарушению архитектоники.
Заключение. С учетом ранее охарактеризованной нами морфологической картины печени крыс обоих полов и ряда биохимических параметров, описывающих ее функциональное состояние, полученные данные о межполовых различиях в ультраструктурных характеристиках гепатоцитов крыс, подверженных воздействию хронической алкогольной интоксикации и постоянного освещения, позволяют нам высказать утверждение о более успешном протекании адаптации гепатоцитов к стрессорным условиям у самок крыс, нежели у самцов.
Работа выполнена в рамках государственного задания НИИ морфологии человека им. Авцына ФГБНУ «Петровский национальный исследовательский центр хирургии» № 122030200535-1.
Список литературы Влияние совместного действия постоянного освещения и хронической алкогольной интоксикации на ультраструктуру гепатоцитов самцов и самок крыс «Вистар»
- Агаджанян Н.А., Петров В.И., Радыш И.В., Краюшкин С.И. Хронофизиология, хронофармаколо-гия и хронотерапия. Вологоград; 2005. 336.
- Zimmet P., Alberti K.G.M.M., Stern N., Bilu C., El-Osta A., Einat H., Kronfeld-Schor N. The Circadian Syndrome: is the Metabolic Syndrome and much more. Journal of internal medicine. 2019; 286 (2): 181-191.
- Gubin D.G., Kolomeichuk S.N., Weinert D. Circadian clock precision, health, and longevity. J. Chro-nomed. 2021; 23 (1): 3-15.
- Bumgarner J.R., Nelson R.J. Light at Night and Disrupted Circadian Rhythms Alter Physiology and Behavior. Integr Comp Biol. 2021; 61 (3): 1160-1169.
- Farkova E., Schneider J., SmotekM., Bakstein E., Herlesova J., Koprivova J., FriedM. Weight loss in conservative treatment of obesity in women is associated with physical activity and circadian phenotype: A longitudinal observational study. BioPsychoSocial Medicine. 2019; 13: 1-10.
- Пудиков И. Некоторые аспекты количественной оценки влияния света на циркадианные физиологические функции. Полупроводниковая светотехника. 2020; 2: 30-36.
- Хабаров С.В., Стерликова Н.А. Мелатонин и его роль в циркадной регуляции репродуктивной функции (Обзор литературы). Вестник новых медицинских технологий. 2022; 29 (3): 17-31.
- Talib W.H., Alsayed A.R., Abuawad A., Daoud S., Mahmod A.I. Melatonin in cancer treatment: current knowledge and future opportunities. Molecules. 2021; 26 (9): 2506.
- Han Y., Chen L., Baiocchi L., Ceci L., Glaser S., Francis H., Alpini G., Kennedy L. Circadian Rhythm and Melatonin in Liver Carcinogenesis: Updates on Current Findings. Crit Rev Oncog. 2021; 26 (3): 69-85.
- Nelson R.J., Chbeir S. Dark matters: effects of light at night on metabolism. Proc Nutr Soc. 2018; 77 (3): 223-229.
- Walker W.H. 2nd, Bumgarner J.R., Walton J.C., Liu J.A., Melendez-Fernandez O.H., Nelson R.J., DeVriesA.C. Light Pollution and Cancer. Int J Mol Sci. 2020; 21 (24): 9360.
- Kramer A., Lange T., Spies C., Finger A.M., Berg D., Oster H. Foundations of circadian medicine. PLoS biology. 2022; 20 (3): e3001567.
- Gonzalez D., Justin H., Reiss S., Faulkner J., Mahoney H., Yunus A., Gulick D. Circadian rhythm shifts and alcohol access in adolescence synergistically increase alcohol preference and intake in adulthood in male C57BL/6 mice. Behavioural Brain Research. 2023; 438: 114216.
- Karlsson H., Persson E., Perini I., Yngve A., HeiligM., Tinghog G. Acute effects of alcohol on social and personal decision making. Neuropsychopharmacology. 2022; 47 (4): 824-831.
- Tamura E.K., Oliveira-Silva K.S., Ferreira-Moraes F.A., Marinho E.A., Guerrero-Vargas N.N. Circadian rhythms and substance use disorders: A bidirectional relationship. Pharmacology Biochemistry and Behavior. 2021; 201: 173105.
- Katary M., Abdel-Rahman A.A. Alcohol suppresses cardiovascular diurnal variations in male normotensive rats: Role of reduced PER2 expression and CYP2E1 hyperactivity in the heart. Alcohol. 2020; 89: 27-36.
- Tice A.L., Laudato J.A., Fadool D.A., Gordon B.S., Steiner J.L. Acute binge alcohol alters whole body metabolism and the time-dependent expression of skeletal muscle-specific metabolic markers for multiple days in mice. American Journal of Physiology-Endocrinology and Metabolism. 2022; 323 (3): E215-E230.
- KurhalukN. Alcohol and melatonin. Chronobiol Int. 2021; 38 (6): 785-800.
- Costa R., Mangini C., Domenie E.D., Zarantonello L., Montagnese S. Circadian rhythms and the liver. Liver International. 2023; 43 (3): 534-545.
- Nicolaides N.C., Chrousos G.P. Sex differences in circadian endocrine rhythms: Clinical implications. Eur J Neurosci. 2020; 52 (1): 2575-2585.
- Walton J.C., Bumgarner J.R., Nelson R.J. Sex Differences in Circadian Rhythms. Cold Spring Harb Per-spect Biol. 2022; 14 (7): a039107.
- Layton A. T. His and her mathematical models of physiological systems. Mathematical Biosciences. 2021; 338: 108642.
- Балканов А.С., Розанов И.Д., Голанов А.В., Гаганов Л.Е., Черников В.П. Состояние эндотелия капилляров перитуморальной зоны после адъювантной лучевой терапии глиобластомы головного мозга. Клиническая и экспериментальная морфология. 2021; 10 (1): 33-40.
- Kozlova M.A., Kirillov Y.A., Makartseva L.A., Chernov I., Areshidze D.A. Morphofunctional state and circadian rhythms of the liver under the influence of chronic alcohol intoxication and constant lighting. International Journal of Molecular Sciences. 2021; 22 (23): 13007.
- Areshidze D.A., Kozlova M.A. Morphofunctional State and Circadian Rhythms of the Liver of Female Rats under the Influence of Chronic Alcohol Intoxication and Constant Lighting. International Journal of Molecular Sciences. 2022; 23 (18): 10744.
- Tsomaia K., Patarashvil L., Karumidze N., Bebiashvili I., Azmaipharashvili E., Modebadze I., Kordzaia D. Liver structural transformation after partial hepatectomy and repeated partial hepatectomy in rats: A renewed view on liver regeneration. World Journal of Gastroenterology. 2020; 26 (27): 3899.
- Tan H.K., Yates E., Lilly K., Dhanda A.D. Oxidative stress in alcohol-related liver disease. World journal of hepatology. 2020; 12 (7): 332.
- Prasun P., Ginevic I., Oishi K. Mitochondrial dysfunction in nonalcoholic fatty liver disease and alcohol related liver disease. Transl Gastroenterol Hepatol. 2021; 6: 4. DOI: 10.21037/tgh-20-125.
- Sergi C.M., Sergi C.M. Parenchymal GI Glands: Liver. Pathology of Childhood and Adolescence: An Illustrated Guide. Springer; 2020: 425-549.
- Shi Y., Liu Y., Wang S., Huang J., Luo Z., Jiang M., You J. Endoplasmic reticulum-targeted inhibition of CYP2E1 with vitamin E nanoemulsions alleviates hepatocyte oxidative stress and reverses alcoholic liver disease. Biomaterials. 2022; 288: 121720.
- Teschke R., Xuan T.D. Heavy metals, halogenated hydrocarbons, phthalates, glyphosate, cordycepin, alcohol, drugs, and herbs, assessed for liver injury and mechanistic steps. Frontiers in Bioscience-Landmark. 2022; 27 (11): 314.
- Slevin E., Baiocchi L., Wu N., Ekser B., Sato K., Lin E., Meng F. Kupffer cells: Inflammation pathways and cell-cell interactions in alcohol-associated liver disease. The American Journal of Pathology. 2020; 190 (11): 2185-2193.
- BlázovicsA. Alcoholic liver disease. In: Influence of Nutrients, Bioactive Compounds, and Plant Extracts in Liver Diseases. Academic Press; 2021: 57-82.
- НалобинД.С., Супруненко Е.А., ГоличенковВ.А. Влияние мелатонина на дифференцировочный потенциал клеток Ито при индуцированном фиброзе печени мыши. Бюллетень экспериментальной биологии и медицины. 2016; 161 (6): 807-811.
- Greuter T., Malhi H., Gores G.J., Shah V.H. Therapeutic opportunities for alcoholic steatohepatitis and nonalcoholic steatohepatitis: exploiting similarities and differences in pathogenesis. JCI Insight. 2017; 2 (17): e95354.
- Owino S., Contreras-Alcantara S., Baba K., Tosini G. Melatonin signaling controls the daily rhythm in blood glucose levels independent of peripheral clocks. PloS One. 2016; 11 (1): e0148214.
- Guan Q., Wang Z., Cao J., Dong Y., Chen Y. The role of light pollution in mammalian metabolic homeostasis and its potential interventions: A critical review. Environmental Pollution. 2022; 312: 120045.
- Ozaki M. Cellular and molecular mechanisms of liver regeneration: Proliferation, growth, death and protection of hepatocytes. Seminars in cell & developmental biology. 2022; 100: 62-73.
- Solhi R., Lotfinia M., Gramignoli R., Najimi M., Vosough M. Metabolic hallmarks of liver regeneration. Trends in Endocrinology & Metabolism. 2021; 32 (9): 731-745.
- Maitra D., Carter E.L., Richardson R. Oxygen and conformation dependent protein oxidation and aggregation by porphyrins in hepatocytes and light-exposed cells. Cellular and Molecular Gastroenterology and Hepatology. 2019; 8 (4): 659-682.
- Contreras-Zentella M.L., Villalobos-García D., Hernández-Muñoz R. Ethanol metabolism in the liver, the induction of oxidant stress, and the antioxidant defense system. Antioxidants. 2022; 11 (7): 1258.
- Sato K., Meng F., Francis H., Wu N., Chen L., Kennedy L., Zhou T., Franchitto A., Onori P., Gaudio E., Glaser S., Alpini G. Melatonin and circadian rhythms in liver diseases: Functional roles and potential therapies. J Pineal Res. 2020; 68 (3): e12639.
- Mierke C.T., Mierke C.T. Lysosomes and Peroxisomes. In: Cellular Mechanics and Biophysics: Structure and Function of Basic Cellular Components Regulating Cell Mechanics. Springer; 2020: 277-332.
- Palmisano B.T., Zhu L., Stafford J.M. Role of Estrogens in the Regulation of Liver Lipid Metabolism. Adv Exp Med Biol. 2017; 1043: 227-256.
- Lee C., Kim J., Jung Y. Potential Therapeutic Application of Estrogen in Gender Disparity of Nonalcoholic Fatty Liver Disease/Nonalcoholic Steatohepatitis. Cells. 2019; 8 (10): 1259.
- Schmidt-Arras D., Rose-John S. IL-6 pathway in the liver: from physiopathology to therapy. Journal of hepatology. 2016; 64 (6): 1403-1415.
- Dukic M., Radonjic T., Jovanovic I., Zdravkovic M., Todorovic Z., Kraisnik N. Alcohol, Inflammation, and Microbiota in Alcoholic Liver Disease. International Journal of Molecular Sciences. 2023; 24 (4): 3735.
