Влияние современного глобального потепления на лесные экосистемы Жигулевского массива (прогнозное эмпирико-статистическое моделирование)
")
Автор: Коломыц Эрланд Георгиевич, Сурова Наталья Анатольевна
Журнал: Самарская Лука: проблемы региональной и глобальной экологии @ssc-sl
Рубрика: Наземные экосистемы
Статья в выпуске: 3 т.30, 2021 года.
Бесплатный доступ
Прогноз осуществлен по разработанной методике на основе трех глобальных прогнозно-климатических моделей - умеренных GISS и E GISS и экстремальной HadCM3. Рассматриваются механизмы отклика локальных экосистем на глобальные гидротермические сигналы через летнее влагосодержание почвы как ведущий экологический канал связи. Описаны траектории преобразований в предстоящие 100-200 лет семигумидных и семиаридных лесных экосистем Жигулевского низкогорного массива к термоаридному сигналу. Прогнозируется резкая трансформация мезофильных широколиственнолесных биогеоценозов жигулевского плато. Начнется их интенсивный распад и поглощение луговой степью. Еще более быстрая аридизация пойдет в ксерофитных лесо-лугово-степных сообществах и в сосново-широколиственных лесах.
Волжский бассейн, жигулевский массив, лесные экосистемы, глобальное потепление, прогнозные модели, аридизации лесного покрова
Короткий адрес: https://sciup.org/148323668
IDR: 148323668 | УДК: 91.929+57.557+51/519.2 | DOI: 10.24412/2073-1035-2021-10408
The impact of modern global warming for forest ecosystems of Zhigulevsky massif (predictive empirical-statistical modeling)
Prognosis was made using a method worked out by the authors on the basis of three global prediction-climatic models - temperate GISS and E GISS and extremal HadCM3. The mechanisms of the response of local ecosystems to global hydrothermal signals through the summer moisture content of the soil are considered as the leading ecological communication channel. The trajectories of transformations of semi-humid and semi-arid forest ecosystems to the thermo-arid signal during forthcoming 100-200 years are described. A sharp transformation of mesophilic broadleaf biogeocoenoses of the Zhiguli plateau is predicted. They will be intensively decomposed and substituted by meadow steppe. The most xerophyte forest-meadow-steppe associations and pine broad-leaved forest will be aridity in still more greater degrees.
Текст научной статьи Влияние современного глобального потепления на лесные экосистемы Жигулевского массива (прогнозное эмпирико-статистическое моделирование)
ВВЕДЕНИЕ (Le Quere et al., 2015). При этом региональное
В последние десятилетия резко возросло число публикаций, в которых приводятся основные глобальные (а в некоторых случаях и региональные) прогнозные характеристики предстоящих и, по существу, уже начавшихся антропогенных изменений климата. Климатические прогнозы, основанные на сценариях техногенных выбросов парниковых газов в атмосферу, предполагают повышение средней глобальной температуры поверхности Земли в пределах 1,4–5,8° за период с 1990 г. по 2100 г., что в 2–10 раз больше величины потепления, произошедшего в ХХ в. (Албриттон и др., 2003). Однако реальная картина значительно превосходит и эти расчеты. Существующие в настоящее время тренд потепления приведет к повышению
потепление на территории России может составить порядка 6–11° (Швиденко и др., 2017).
Необходимость сохранения в условиях меняющегося климата лесных экосистем и воспроизводства лесных ресурсов на южной границе лесной зоны умеренных широт, где лесные сообщества находятся в состояниях, близких к критическим, относится к числу важных проблем биоэкологического и геосистемного мониторинга. Ключевое значение здесь имеет установление причинных механизмов локального отклика на глобальные и региональные сигналы, с выявлением передаточные функции в ландшафтных связях, которые трансформируют эти сигналы при их прохождении через сопряженные ансамбли как отдельных геокомпонентов, так и природных комплексов в целом.
Пути решения этой задачи наметились в процессе выполнения наших региональных прогнозно-экологических исследований
(Коломыц, 2008; Сурова, 2015). Эмпирически было установлено, что основной пропускной канал связей региональных и локальных экосистем с фоновой климатической системой проходит через летнее влагосодержание почвы, что, с одной стороны, служит достаточно надежным геофизическим индикатором состояния природных комплексов, а с другой, – является наиболее мощным прямодействующим экологическим фактором, который предопределяет территориальное распределение растительности, почв и в целом природных комплексов. Для основного водосбора Волжского бассейна найдены тесные связи июльских запасов продуктивной влаги в почве с годовым коэффициентом увлажнения Высоцкого-Иванова – важнейшим ландшафтногеофизическим показателем, отражающим зональные условия территории (Коломыц, 2008). С другой стороны, существует достаточно тесная зависимость распределения лесных топоэкосистем от летнего влагосодержания почвы (рис. 1). По нашим измерениям и расчетам, для Жигулевского массива информационные коэффициенты сопряженности (Пузаченко, Скулкин, 1981) экосистем с запасами продуктивной влаги в слоях почвы 0–20 см ( W-20 ), 0–50 см ( W-50 ) и 0– 100 см ( W-100 ) равны соответственно 0,138, 0,148 и 0,171, что существенно превышает критический порог значимости связей (0,070). Наиболее эффективное воздействие на состояние региональных и локальных гидроэдафотопов оказывают гидротермические факторы вегетационного периода: средняя температура июля, осадки теплого периода года, суммарное испарение, речной сток.
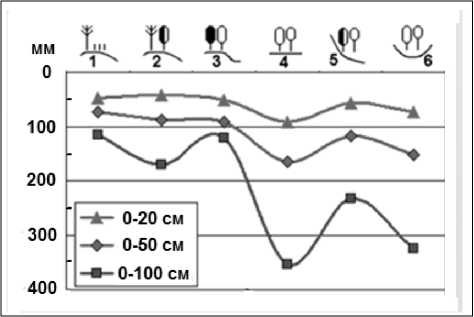
Рис. 1. Распределение таксономических норм среднеиюльского базового содержания продуктивной влаги в различных слоях почвы по группам биогеоценозов Жигулевского массива.
Fig. 1. Distribution of taxonomic norms of the Middle July basic content of productive moisture in various soil layers by groups of biogeocoenoses of the Zhigulevsky massif.
Установлено также, что приоритетным объектом ландшафтно-экологического прогнозирования в пределах столетия должна служить не структурная эволюция экосистем, а направленная смена их функционирования, т. е. сдвиги в малом биологическом круговороте (продуцировании и разложении фитомассы), которые занимают в таежной зоне первые несколько лет, а в подзоне широколиственных лесов завершаются в течение года (Коломыц, 2003).
ИСХОДНЫЕ МАТЕРИАЛЫ
Прогнозно-экологическое моделирование локального отображения глобальных климатических изменений целесообразно проводить в первую очередь на зональных экотонах, которые отличаются повышенной чувствительностью природных экосистем к воздействию климата (Меняющийся мир…, 1991; Коломыц, 2003). В этом отношении особое место занимает переходная полоса умеренного пояса от леса к степи, где при годовом соотношении тепла и влаги, близком к 1, лесные экосистемы оказываются в критических состояниях, испытывая значительную конкуренцию со стороны травянистых формаций. Жигулевский низкогорный массив расположен как раз на зональной границе южной лесостепи с северной степью, создавая местные климатические условия более северной подзоны – типичной (средней) лесостепи. Благодаря этому, а также историческим причинам, на территории Самарской Луки произошло взаимопроникновение с севера и северо-запада не только неморальнолесных, но и бореальных (преимущественно боровых) элементов флоры, а с юга и юго-востока – степных видов. Это привело к смешению флор и большому фитоценотическому разнообразию жигулевских топоэкосистем, которое предопределено в первую очередь солярно-экспозиционными контрастами склонов низ-когорья. Возникла даже смешанная лесо-луговостепная зонально-географическая группа (своеобразного горного облика), с примесью бореальных боровых элементов.
В июле 1996 г. нами была проведена ландшафтно-экологическая съемка на экспериментальном полигоне площадью около 20 км2 в районе Ширяевской долины и ее окрестностей, с выходом на жигулевское плато (координаты полигона – 53o23ˊ с.ш. и 50o00ˊ в.д.). Методика проведения съемки изложена нами ранее (Коломыц и др., 1993; Коломыц, 2008). Всего было описано 40 пробных площадей, каждая из них представляла собой конкретный лесной биогеоценоз. Все они были сгруппированы в шесть биогеоценотических (фациальных) групп, краткая характеристика которых представлена в табл. 1. Информация, зафиксированная на каждой пробной площади размером 20 × 20 м, состояла из 76 геокомпонентных и комплексных признаков, объединенных в пять блоков: 1) гидро-геоморфологический; 2) почвенный морфологический; 3–4) фитоцено-тические структурный и функциональный; 5) ландшафтногеофизический. Сбор фактического материала на каждой пробной площади, его обработка и последующие расчеты проводились в соответствии с известными методическими руководствами.
На основе отношений мер включения (Андреев, 1980) 23 параметров функционального фито-ценотического блока топоэкосистем: фитомасс, продуктивности и комплексных параметров метаболизма (см. Коломыц, 2008, Приложение 1), – установлены их экологические группы (экотипы), отвечающие различным уровням производства и разложения органического вещества. На структурной схеме результирующая картина уровней организации лесных биогеоценозов зафиксирована ребрами орграфов их отношений включения (рис. 1). Стрелки связей направлены от менее развитых объектов (биогеоценотиче-ских групп) в сторону более высокоразвитых. Для каждого объекта определялись число входных связей (ni) и число связей выходных (nj). Подсчитывалось также N – общее число связей в системе. Исходя из этого, определялся структурно-метаболический индекс Smk k-го объекта как
Smk = Pi – Pj + 1, где Pi = ni / N, Pj = nj / N. Очевидно,
0 ≤ Sm k ≤ 2.
Таблица 1
Группы биогеоценозов (ландшафтных фаций) Жигулевского низкогорного массива и их условные обозначения
Groups of biogeocoenoses (landscape facies) of the Zhigulevsky low-mountain massif and their symbols
Краткая характеристика
-
1. Ксероморфные и мезоморфные сосняки, остепненные, карагановые и разнотравнозлаковые, верхних и средних частей крутых (20–30°) солнцепечных склонов (трансэлювиальные и транзитные), с дерново-карбонатными типичными маломощными легкосуглинистыми почвами на элювии известняка.
-
2. Мезоморфные, ксеро-мезоморфные и ксероморфные сосняки сложные (липоводубовые) и сосново-широколиственные леса, лещинные, неморальнотравяные, верхних и средних частей сравнительно крутых (до 20–25°) нейтральных и теневых склонов (трансэлювиальные и транзитные), с дерново-карбонатными типичными легко- и среднесуглинистыми почвами на элювии известняка.
-
3. Мезоморфные и ксеро-мезоморфные дубняки кленово-липовые и производные от них осинники, лещинные, разнотравно-ландышевые, верхних, средних и нижних частей нейтральных и теневых склонов средней и высокой крутизны (от трансэлювиальных до трансаккумулятивных), с дерново-карбонатными маломощными среднесуглинистыми почвами на элювии известняка.
-
4. Мезо-гидроморфные и мезоморфные теневые широколиственные леса (вязово-кленово-липовые), разнотравно-снытьевые, липовые и осинники кленово-липовые, разнотравно-снытьевые, пологонаклонных водоразделов (крутизной до 3–5° всех экспозиций (элювиальные), с дерново-карбонатными и темносерыми лесными среднесуглинистыми почвами на элювии глин и известняков.
-
5. Мезоморфные и мезо-гидроморфные теневые широколиственные леса (дубово-вязово-липовые), а также осинники и березняки (дубово-кленово-липовые) лещинные, разнотравно-снытьевые и волосисто-осоковые, средних и нижних частей крутых склонов (транзитные и трансаккумулятивные) с дерново-карбонатными типичными и выщелоченными среднесуглинистыми почвами на делювии.
-
6. Мезоморфные и мезо-гидроморфные липняки, кленовники и осинники (иногда с лещиной), разнотравно-снытьевые, пологонаклонных днищ глубоко врезанных долин малых водотоков, с дерновыми намытыми и дерново-карбонатными выщелоченными среднесуглинистыми почвами на делювии.
Обозначение


По параметру S m k группы лесных фитоценозов были подразделены на метаболические экотипы . Как известно, понятие экотипов используется для выделения экосистем, адаптированных к местным почвенно-климатическим условиям (Одум, 1986). При S m k >> 1 относительный уровень развития объекта достаточно высок и в системе фитоценотических и ландшафтных переходов он входит в группу абсорбентов. Это субоптимальная категория – субдоминант , который в функциональном аспекте именуется как суб-макроболический экотип. Если при этом n j = 0, то объект входит в оптимальную категорию, становится доминантом – макроболитом . По мере снижения своего метаболического индекса объект становится сначала рядовым членом данного геопространства, т. е. мезоболипом . Наконец, при S m k < < 1 он отличаются наиболее слабо развитой структурой, а в системе переходов оказывается редуцентом, поэтому переходит в субпессималь-ную и пессимальную категории редуцированных экотипов – субмикроболитов и микроболитов. В последнем случае на структурной схеме доминирования оно не имеют входных связей ( n i = 0).
Повышенная раздробленность функционального пространства в региональной экосистеме Жигули (табл. 1 и рис. 1) предопределяет достаточно пеструю мозаику локальных экосистемных реакций на одно и тоже фоновое климатическое воздействие. При этом низкогорные экосистемы весьма жестко «привязаны» к контрастным типам местоположений, и это создает орографические барьеры на пути латерального распространения волн гидротермических сигналов. Доминирующими экосистемами (макроболитами) являются умеренно влажные плакорные неморальнолесные биогеоценозы (мезофильные липняки). Им свойствен наиболее интенсивный метаболизм, соответствующий нормам восточноевропейских широколиственных лесов: максимальные величины фитомасс и продуктивности и наибольшая скорость разложения мертвой органики. Пониженный уровень функционирования имеют субдоминанты (субмакроболиты) – сосново-широколиственные леса и липо-дубняки верхних теневых и нейтральных склонов и их подножий.
Мезоболиты в Жигулевском массиве почти отсутствуют, и наблюдается сразу переход к редуцированным топоэкосистемам – субмикроболи-там и микроболитам. Они отличаются минимальными запасами живой фитомассы, наименьшей продуктивностью фитоценозов и сильно заторможенным биологическим круговоротом, который характерен для зональных условий северотаежных лесов Восточной Европы и/или подтайги резко континентальной Сибири. Это, с одной стороны, ксероморфные остепненные сосновые боры крутых солнцепечных склонов и привершинные дубовые леса нейтрально-теневых экспозиций, а с другой, – гидроморфные лиственные леса днищ малых глубоко врезанных долин.
Местные геоморфологические и почвенно-эдафические факторы преломляют фоновые био-климатические условия и формируют на климатических экотонах, помимо плакорных топоэко-систем, отражающих зонально-региональный фон данной территории (Высоцкий, 1960), множество экстразональных категорий (Спрыгин, 1986) как своего рода представителей других зональных типов географической среды – не только соседних, но и весьма удаленных. Образуются так называемые региональные системы локальной зональности (Коломыц, 2008), которые имитируют основные направления и масштабы местных экосистемных перестроек, создавая тем самым эмпирическую основу для прогнозных построений. Выделенные нами группы лесных биогеоценозов отчетливо дифференцируются по их различной зональной принадлежности (табл. 2), поэтому в функциональных и структурных переходах экосистем локального уровня должны адекватно отразиться фоновые сдвиги зонально-региональной биоклиматической системы. По сменам зональных групп биогеоценозов можно оценить, насколько локальные метаморфозы способны преобразовать весь зональнорегиональный фон данной территории под действием того или иного климатического тренда. По степени гидроморфности все лесные экосистемы Жигулевского массива объединены в две плеяды – и семиаридную (группы фаций 1 и 2) и семигумидную (все остальные группы).
ОСНОВНЫЕ ЧЕРТЫ КЛИМАТИЧЕСКОГО ПРОГНОЗА
Ландшафтно-экологические прогнозные оценки основаны на двух глобальных прогнозноклиматических моделях общей циркуляции атмосферы AOGCMs: 1) модели GISS (Hansen et al., 1988), дающей климатический прогноз до 2100 г., и ее поздней версии E GISS (Hansen et al., 2007) c прогнозом до 2200 г.; 2) модели HadCM3, версия А2 (Pope et al., 2000). Выбранные для Самарского Поволжья значения климатических прогнозов приведены в табл. 3. Био-климатический прогноз состоит из 4-х шагов: 1996–2050–2100–2150–2200 гг. Базовым считается период 1981–1980 гг., с конца которого и началось современное глобальное потепление (Houghton et al., 1996).
Как видно, модель HadCM3 дает более экстремальные значения антропогенного потепления, нежели модели GISS и E GISS. Уже на первом шаге прогнозного периода (1996–2050 гг.)
разница в приращении среднеиюльских температур составляет около 1,2°. Затем эта разница резко увеличивается и составляет к концу второго, третьего и четвертого прогнозных шагов соответственно 1,3°, 2,1° и 2,8°. Соответственно по модели HadCM3 более кардинальными должны быть и экологические сдвиги, чему в первое столетие будет способствовать также более резкое повышение зимних температур. Экологический прогноз по этой модели позволяет оценить реакцию лесных экосистем на такие глобальные климатические изменения, которые могут превышать порог выносливости основных лесообразующих пород и привести к распаду на обширных пространствах не только неморальных, но и бореальных лесов.
Таблица 2
Распределение групп биогеоценозов Жигулевского массива в поле их уровней функционирования и годовых коэффициентов атмосферного увлажнения ( К увл )
Distribution of groups of biogeocoenoses of the Zhigulevsky massif in the field of their levels of functioning and annual atmospheric humidity factors (Fhum)
|
cd 02 S о к & 02 S О Щ а 9 и Щ |
I Макроболиты |
99 4 |
|||
|
II Субмакроболиты |
TV 2 |
t? 5 |
|||
|
IV Субмикроболиты |
1 III 1 |
3 |
|||
|
V Микроболиты |
Я 6 |
||||
|
К увл ( F hum) |
1 – 0.82 |
4 – 0.95 |
2 – 0.93 6 – 0.91 |
5 – 1.06 3 – 1.05 |
|
|
Зоны /подзоны – аналоги, восточный сектор Русской равнины (зональный коэффициент увлажнения) |
Типичная и южная лесостепь (0,76–0,95) |
Широколиственные леса (0,95–1,05) |
Типичная лесостепь (0,86–0,95) |
Смешанные леса (1,05–1,35) |
|
Рост летних и зимних температур будет сопровождаться определенным увеличением количества осадков, особенно в теплый период года. Однако прибавка осадков не сможет нейтрализовать повышение температуры, поэтому годовой и летний коэффициенты увлажнения начнут почти повсеместно и весьма существенно снижаться (см. табл. 3). Начнется также прогрессирующее летнее иссушение почвы. По нашим расчетам, основанным на эмпирических данных, для того, чтобы июльские запасы продуктивной почвенной влаги не изменились, повышение летней температуры на 1oС должно быть скомпенсировано прибавкой осадков для слоев почвы 0–20 см, 0–50 см и 0–100 см соответственно в 135 мм, 165 мм и 250 мм. Согласно модели HadCM3, к 2100 г. средние июльские температуры в районе Самарской Луки вырастут более чем на 4°, и для нейтрализации такого роста необходима мини- мальная прибавка годовых осадков около 570 мм, что не прогнозируется для данного региона ни одной моделью семейства AOGCMs (Houghton et al., 1996).
Таким образом, вплоть до конца XXII столетия на территории Самарской Луки, равно как и по всему Волжскому бассейну (Коломыц, 2003), будет развиваться термо-аридный климатический тренд, подтверждаемый общим снижение годового коэффициента атмосферного увлажнения (см. табл. 2) . Ожидаемое глобальное антропогенное потепление вызовет общую аридиза-цию региональной биоклиматической системы, и чем дальше по времени, тем эта аридизация будет все более глубокой и тотальной. Наиболее значительное иссушение намечается во второй половине XXI в., когда повышение температуры будет сопровождаться сокращением количества осадков.
Таблица 3
Значения основных климатических параметров и соответствующие ландшафтно-зональные условия, прогнозируемые по расчетным моделям GISS /E GISS и HadCM3 для территории Самарской Луки (метеостанция Сосновый Солонец)
The values of the main climatic parameters and the corresponding landscape-zonal conditions predicted by the computational models GISS / E GISS and HadCM3 for the territory of Samarskaya Luka (meteorological station Sosnovyy Solonets)
А. Согласно моделям GISS и E GISS
|
Климатические параметры *) |
Базовый период |
Прогнозируемые сроки |
|||
|
Модель GISS |
Модель E GISS |
||||
|
2050 г. |
2100 г. |
2150 г. |
2200 г. |
||
|
t янв |
–13,9 |
–13,2 |
–9,2 |
–6,2 |
–3,2 |
|
t июля |
20,1 |
21,3 |
23,0 |
23,5 |
24,4 |
|
r хп |
261 |
296 |
340 |
350 |
360 |
|
r тп |
368 |
362 |
407 |
435 |
460 |
|
r год |
629 |
658 |
747 |
785 |
820 |
|
Е о |
659 |
775 |
970 |
1035 |
1159 |
|
К увл |
0,95 |
0,85 |
0,77 |
0,76 |
0,71 |
|
Зональная принадлежность |
Типичная лесостепь, у ее северной границы |
Типичная и южная лесостепь |
Южная граница лесостепи |
Северная степь |
|
Б. Согласно модели HadCM3
|
Климатические параметры *) |
Базовый период |
Прогнозируемые сроки |
|||
|
2050 г. |
2100 г. |
2150 г. |
2200 г. |
||
|
t янв |
–13,9 |
–12,6 |
–8,1 |
–6,5 |
–8,0 |
|
t июля |
20,1 |
22,5 |
24,3 |
25,6 |
27,2 |
|
r хп |
261 |
284 |
307 |
365 |
290 |
|
r тп |
368 |
361 |
315 |
317 |
289 |
|
r год |
629 |
645 |
622 |
682 |
579 |
|
Е о |
659 |
910 |
1115 |
1340 |
1609 |
|
К увл |
0,95 |
0,71 |
0,56 |
0,51 |
0,36 |
|
Зональная принадлежность |
Типичная лесостепь, у ее северной границы |
Северная степь |
Средняя степь |
Граница между средней и южной степью |
Южная степь |
Примечание . *) Средние температуры: t янв – января, t июля – июля; количество осадков: r хп – холодного периода, r тп – теплого периода, r год – годовое; Е о – годовая испаряемость; К увл – годовой коэффициент атмосферного увлажнения Высоцкого-Иванова.
Год проведения ландшафтно-экологических съемок в Жигулях вошел в самое теплое пятилетие (1995–1999 гг.) за весь период инструментальных наблюдений (Израэль, 2001), что подтверждаются летними температурами и расчетными значениями запасов продуктивной влаги в почве. Средняя температура июля была на 1,5o выше базовой нормы. Она не только превысила термический уровень 2050 г., прогнозируемый по модели GISS (см. табл. 2), но и приблизилась к тому уровню данного срока (22,0o для Жигулевских гор, по ст. Сосновый Солонец), который дает более экстремальная модель HadCM3.
Несмотря на повышенное годовое количество осадков и более низкие зимние температуры, способствовавшие уменьшению потерь снегоза-пасов при оттепелях и повышению весенней влагозарядки почвы, значительный рост летних температур оказал решающее отрицательное влияние на водный режим почвы в данный вегетационный период. Коэффициент увлажнения за 1995–1996 гидрологический год по ст. Сосновый Солонец оказался равным 0,86, что согласно модели GISS, должно составить многолетнюю норму только после 2050 г. (см. табл. 2). Лесные экосистемы Жигулевского массива оказались в гидротермическом режиме южной лесостепи, т.е. произошло их функциональное смещение к югу на целую подзону (см. выше). В результате июльские запасы продуктивной влаги в верхнем слое почвы 0–20 см не превысили 67% от базовой нормы (15 мм), а во всей метровой толще снизились почти в два раза (с 72 мм до 46 мм). Таким образом, полученные в результате съемки структурно-функциональные характеристики лесных биогеоценозов Жигулевских гор уже отображают начавшееся глобальное потепление.
МЕТОДЫ ПРОГНОЗНО-ЭКОЛОГИЧЕСКИХ РАСЧЕТОВ
Весь алгоритм прогнозных расчетов состоял из следующих этапов.
-
I. Выявление (по данным 120 агрометстанций Волжского бассейна и его окружения) зональнорегиональных связей июльских запасов продуктивной почвенной влаги открытых сельскохозяйственных полей с основными климатическим параметрами – средними температурами января ( t янв ) и июля ( t июля ), и годовым количеством осадков ( r год ), а также суммами осадков холодного и теплого периодов. Данный этап расчетов был необходим, поскольку только для агрометстанций можно получить массовый многолетний материал по запасам влаги в почве. Указанные параметры фигурируют в исходных прогнозноклиматических моделях.
Для слоев почвы 0–20 см, 0–50 см и 0–100 см были получены следующие уравнения множественной регрессии:
(W-20) = 78,7 + 0,1212∙ t янв – 3,85∙ t июля +
+ 0,02324∙ r год ;
R = 0,86; R2 = 0,74;
(W-50) = 166,9 + 0,0394∙ t янв – 8,66∙ t июля +
+ 0,06537∙ r год ;
R = 0,86; R2 = 0,74;
(W-100) = 426,7 + 0,1633∙ t янв – 20,12∙ t июля +
+ 0,07998∙ r год ;
R = 0,86; R2 = 0,74.
Как видим, зимнее потепление и рост годовых осадков дают некоторую прибавку июльских влагозапасов в почве, а летнее – весьма сильное их сокращение (за счет усиления испарения), причем на порядок большее. В южной лесостепи и северной степи изменение весенне-летнего влагосодержания почвы на 10 мм может быть вызвано либо сдвигом зимних температур на 7,5–8,0o, либо изменением годовой суммы осадков на 75–80 мм, либо ростом или снижением летних температур всего на 0,9o. Превалирующая роль летних температур очевидна, и хотя зимнее потепление вместе с увеличение годовой суммы осадков способствуют увеличению влагозапасов в почве, это приращение с лихвой перекрывается их убылью в весенне-летний период из-за гораздо более эффективного роста температур. Даже если принять летнее потепление в 2–3 раза меньшее, чем зимнее (что предполагается некоторыми климатическими моделями), процесс аридизации почвы неизбежен.
Для прогнозных расчетов июльского влагосо-держания почвы агроэкосистем Жигулевского массива были использованы полученные нами экспоненциальные связи параметров (W-20) , (W-50) и (W-100) со среднеиюльской температурой (с учетом преобладающего здесь легко-и среднесуглинистого почвообразующего субстрата):
(W-20) = exp [9,033 – 0,3266∙ t июля ];
R = 0,92; R2 = 0,84;
(W-50) = exp [9,723 – 0,3179∙ t июля ];
R = 0,89; R2 = 0,79;
(W-100) = exp [10,43 – 0,3153∙ t июля ];
R = 0,90; R2 = 0,81.
Следует оговориться, что расчеты по этим зависимостям дают несколько большее летнее иссушение почвы, чем использование приведенных выше уравнений множественной регрессии, поскольку не учитывается компенсирующее влияние t янв и r год .
-
II. Установление переходных зависимостей (для трех указанных параметров летнего влаго-содержания почвы) от сельскохозяйственных полей агрометстанций W(поле) к лесопокрытым площадям W(лес) в соответствии с зональнопровинциальными условиями региона. Для Жигулевского массива были использованы данные многолетних наблюдений Нижнедевицкой воднобалансовой станции (Воронежская область), расположенной так же в подзоне южной лесостепи. Линейное уравнение связи имеет вид (для любого срока весенне-летнего периода и для любого слоя почвы):
W(лес) = 0, 9912∙ W(поле) + 7,2;
R = 0,98; R2 = 0,97.
Несмотря на хорошую тесноту связи, точность самой корреляции (информационный шум) остается невысокой – в пределах 15–20 мм. Соответственно таким же должен быть, очевидно, и порог чувствительности лесных экосистем к изменениям влагосодержания почвы, вызванным климатическим сигналами.
-
III. Прогнозные расчеты изменения летних запасов продуктивной почвенной влаги в различных топоэкосистемах для пяти прогностических сроков: 2050 г., 2100 г., 2050 г. и 2200 г., – по каждой из двух глобальных климатических моделей. Начальное летнее влагосодержание в метровом слое почвы в жигулевских лесных и лесостепных экосистемах, как правило, не превышает соответственно 150 мм и 100 мм, т. е. остается ниже так называемой критической влажности, согласно (Будыко, 1971). В этих условиях испарение почвенной влаги определяется не только испаряемостью, но и самим количеством почвенной влаги. Исходя из этого, нами принято, что при одном и том же повышении температуры запасы продуктивной влаги в различных биогеоценозах уменьшаются пропорционально их начальному (для каждого прогнозного шага-интервала) влагосо-держанию. Чем оно выше, тем больше абсолютные потери влаги в почве к концу данного шага. Одинаковым для всех биогеоценозов остается только относительное уменьшение влагозапасов.
-
IV. Локальный ландшафтно-экологический прогноз: оценка вероятностей и скоростей функциональных сдвигов в системе биогеоценозов, согласно установленному тренду климатогенных изменений июльских запасов почвенной влаги. Построение графо-аналитических моделей вероятностей ландшафтно-экологических переходов между объектами. Под переходом понимается такая смена функционального, а затем и структурного состояния данного объекта, при которой объект принимает (с той или иной вероятностью) черты других объектов-прообразов благодаря тому, что его экологическая (в данном случае гид-роэдафическая) ниша при своем изменении все более пересекается с нишами этих объектов.
В экологическом прогнозировании ключевое значение имеют оценки состояний экосистем и их наиболее вероятных динамических тенденций через призму конкурентных отношений их экологических ниш. В основе этого подхода лежат известные представления об экологической нише объекта в виде некоторой области распределения его состояний в данном экологическом пространстве (Одум, 1975). Вероятностный ландшафтно-экологический прогноз осуществлялся с помощью операций с современными и предполагаемыми гидротермическими нишами экосистем (Коломыц, 2003, 2008). Для этого каждая такая ниша была представлена некоторым «размытым» (дескриптивным) множеством в виде вектора-строки (или вектора-столбца). Составляющими вектора служили нормированное частные коэффициенты связи (Пузаченко, Скулкин, 1981), которые рассматривались как «весовые» коэффи- циенты. Каждый из этих коэффициентов является определенным эквивалентом вероятности (встречаемости) того или иного объекта при данной градации геофизического параметра. Вероятность эта тем выше, чем больше частный коэффициент связи.
Сама методика прогноза уже излагалась нами ранее достаточно подробно и неоднократно (Ко-ломыц и др., 1995; Коломыц, 2003, 2008). Отметим лишь, что предикторами прогнозных расчетов в данном случае служат ожидаемые на каждый прогнозный срок значения влагозапасов в слоях почвы 0–20 см, 0–50 см и 0–100 см, с соответствующими «весовыми» коэффициентами, полученными из частных коэффициентов связей биогеоценозов с гидроэдафотопами (см. выше). Кроме того, учитывается вероятность «остаточных» переходов того или иного объекта в другие прообразы-объекты, отсутствующие в данной операционной системе (выборке), т. е. во вневы-борочные объекты. Для семиаридной плеяды лесных экосистем Жигулевского массива эти переходы оказываются доминирующими и даже единственно возможными.
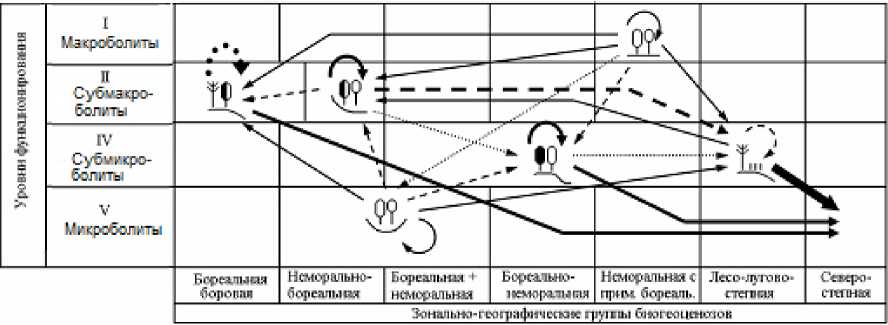
В данном сообщении мы ограничимся этапом получения для каждого прогнозного шага квадратных матриц вероятностей переходов групп биогеоценозов друг в друга ( Р ij ), а также вероятностей стабилизации каждой из них ( Р ii , диагональные элементы матриц). Параметр Р ii рассматривается как мера устойчивости биогеоценозов к данному гидротермическому сигналу. По этим матрицам построены орграфы вероятностей переходов (рис. 3 и 4), описывающие общую картину реакции топоэкосистем на глобальные изменения климата.
На временно́й траектории функциональной трансформации биогеоценозов рост интенсивности ландшафтно-экологических переходов сопровождается усилением избирательности этого процесса. С каждым новым прогнозным шагом функциональный сдвиг каждой лесной экосистемы становится не только более сильным (по отношению к базовому состоянию), но и более однозначным. Это хорошо видно по изменению самой густоты сети переходов. Так, по моделям GISS/E GISS в Жигулевском массиве на первом шаге трансформации топоэкосистем (1996–2050 гг.) насчитывается 17 случаев еще преимущественно слабых межгрупповых переходов, с вероятностью не более 30–40%, то к финишу прогнозного периода (2200 г.) их число сокращается в 1,5 раза, а сама вероятность преобразований повышается до 70–90%. Еще более отчетливо эту закономерность демонстрирует модель HadCM3 (см. рис. 4).
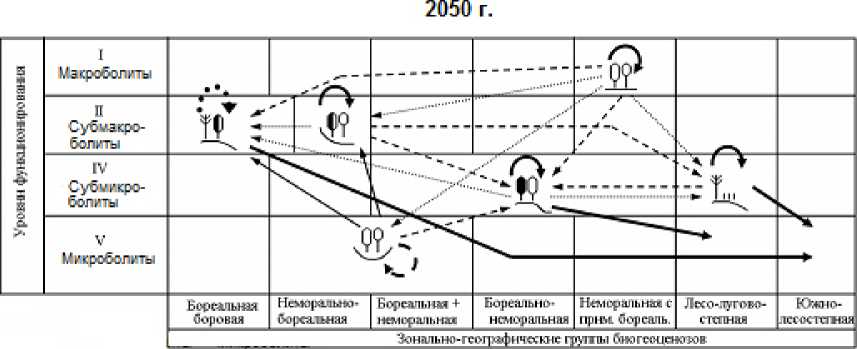
2100 г.
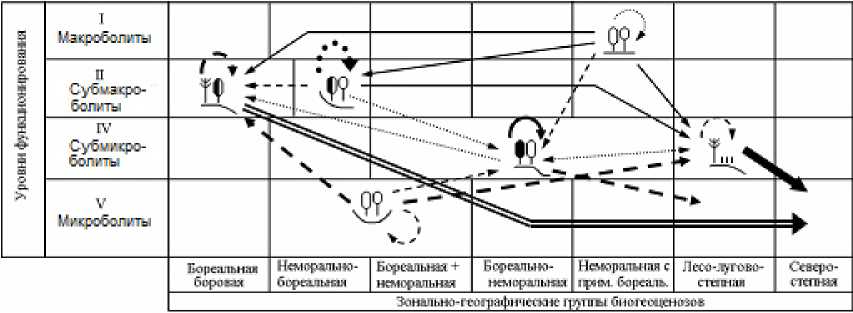
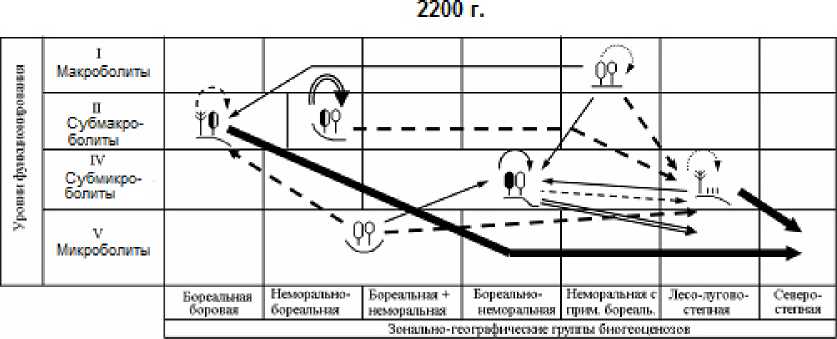
Рис. 3. Орграфы функциональных ландшафтно-экологических переходов, согласно прогнозно-климатической модели GISS, между группами биогеоценозов Жигулевского массива (см. табл. 1), имеющих различную зональную принадлежность и находящихся на соответствующих уровнях базового функционирования.
Вероятности переходов (для рис. 3 и 4): 1 – 0,10 и менее; 2 – 0,11–0,20; 3 – 0,21–0,30; 4 – 0,31–0,40; 5 – 0,41–0,50; 6 – 0,51–0,60; 7 – 0,61–0,70; 8 – 0,70–0,80; 9 – 0,81–0,90; 10 – 0,91–1,00.
Fig. 3. The orgraphs of functional landscape-ecological transitions, according to the GISS predictive-climatic model, between groups of biogeocoenoses of the Zhigulevsky massif (see Table 1), which have different zonal affiliation and are at the corresponding levels of basic functioning. Transition probabilities (for Fig. 3 and 4): 1– 0,10 and less; 2– 0,11–0,20; 3 – 0,21–0,30; 4 – 0,31–0,40;
-
5 – 0,41–0,50; 6 – 0,51–0,60; 7 – 0,61–0,70; 8 – 0,70–0,80; 9 – 0,81–0,90; 10 – 0,91–1,00.
2050 г.
2100 г.
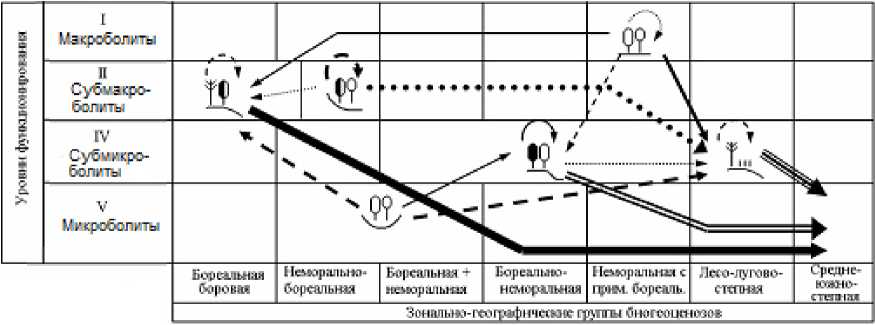
2200 г.
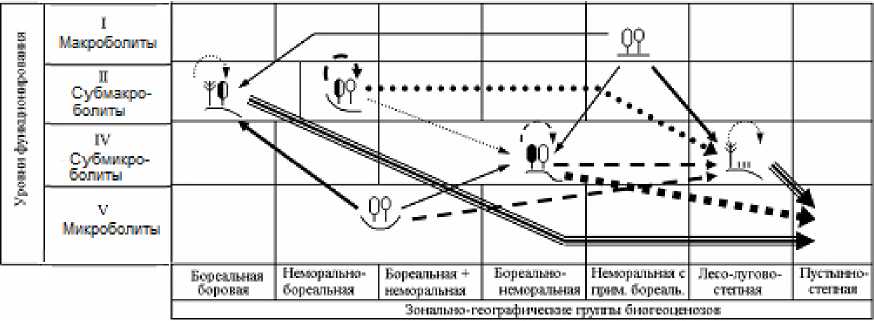
Рис. 4. Орграфы функциональных ландшафтно-экологических переходов, согласно прогнозно-климатической модели HadCM3, между группами биогеоценозов Жигулевского массива (см. табл. 1), имеющих различную зональную принадлежность и находящихся на соответствующих уровнях базового функционирования.
Условные обозначения см. на рис. 3.
Fig. 4. Digraphs of functional landscape-ecological transitions, according to the forecast-climatic model HadCM3, between groups of biogeocoenoses of the Zhigulevsky massif (see Table 1), which have different zonal affiliation and are at the corresponding levels of basic functioning.
For legend see Fig. 3.
ОБЩИЕ ЧЕРТЫ ЭКОСИСТЕМНЫХ ПРЕОБРАЗОВАНИЙ
При более «мягком» климатическом прогнозе, который дает модель GISS, начальные этапы функциональных перестроек топоэкосистем отличаются своего рода высоким «перемешиванием» сети переходов. Во-первых, одна и та же фитоце-нотическая группа лесов начинает приобретать облик сразу нескольких других групп, что отмечается на орграфах переходов определенным множеством выходных стрелок. Во-вторых, нередки случаи парных сочетаний фациальных групп с почти одинаковой взаимной функциональной трансформацией (это показано двухсторонне направленными стрелками). Однако со временем избирательность переходов заметно возрастает, и почти каждая группа лесных экосистем приобретает одно предпочтительное направление своих перестроек, что делает смену функциональных состояний топоэкосистем более детерминированной, а сам экологический прогноз гораздо более определенным.
По модели HadCM3, которая отличается, как уже говорилось, от ГИСовской модели повышенной экстремальностью, предполагается уже с первых шагов более резкая и, вместе с тем, более однозначная (с разреженной сетью переходов) трансформация лесных биогеоценозов. Имеющееся множество фациальных групп все более отчетливо «поляризуется» на две альтернативные категории: 1) доминанты как прообразы будущих состояний большинства топоэкосистем и 2) редуценты, с их деградирующими и исчезающими состояниями. Соответственно изрежива-ются и сами орграфы переходов.
ИЗМЕНЕНИЕ СОСТОЯНИЙ ЛЕСНЫХ ЭКОСИСТЕМ
Наиболее разнообразные изменения должны претерпеть топоэкосистемы семигумидной плеяды, охватывающей два относительно увлажненных «полюса» ландшафтных сопряжений: с одной стороны, мезоморфные липняки жигулевского плато (плакоры), а с другой, – мезо-гидроморфные лиственные леса днищ малых глубоко врезанных долин, а также примыкающие к ним дубо-липняки и осинники нижних частей склонов (рис. 4). К 2200 г. мера P ii их функциональной стабилизации составит от 28–43% по первой прогнозной модели до 15–38% – по второй (см. табл. 3 и рис. 2 и 3).
Максимально широкий спектр трансформации будет свойствен теневым широколиственным лесам жигулевского плато (4–5 выходных стрелок на орграфах), и это произойдет уже на первых двух шагах прогнозного периода (к 2050 и
2100 гг.). При этом если по модели GISS предполагается несколько одновременных вариантов переходов их в ксеромезофитные лесные сообщества горных склонов (группы фаций 2, 3 и 5) с вероятностью каждого варианта до 20–30%, то более экстремальная модель HadCM3 дает прямо-таки катастрофическое олуговение и остепнение плакорных липняков – 40–50-процентную трансформацию их в ксерофитные лесо-лугово-лесные комплексы (группу 1), распространенные ныне только на крутых солнцепечных склонах низкого-рья. На более влажных местообитаниях плато эти экосистемы будут поглощаться также (с вероятностью более 25%) трансэлювиальными мезо-ксерофитными сосняками сложными и сосновошироколиственными лесами (группой 2). Уже на первом прогнозном этапе плакорные липняки на 85% потеряют свою функциональную устойчивость, а в дальнейшем мера их стабилизации сведется к нулю. После 2100 г. процесс их аридиза-ции, по-видимому, несколько замедлится. Наряду с продолжающимся остепнением начнется их столь же значительная трансформация (более чем на 40%) в ксеромезофитные дубняки и осинники, занимающих ныне теневые и нейтральные склоны.
Долинные осиново-липовые фитоценозы на протяжении всего прогнозного периода будут переходить в состояние более сухих широколиственных лесов соседних крутых склонов (с вероятностью 30–40% на каждом шаге), а также приводораздельных местообитаний (на 20–30%).
В конечном итоге к 2150 г. лесные фитоценозы семигумидной плеяды Жигулевских гор практически перестанут существовать как самостоятельные функциональные единства (мера их P ii ≤ 0,1). Начнется весьма интенсивный распад этих лесных сообществ. Особенно крупные территориальные масштабы этот процесс примет в липняках жигулевского плато, занимающих в настоящее время довольно обширные пространства (рис. 5). С наиболее высокого, макроболит-ного, уровня функционирования, с оптимальными показателями продуктивности, эти леса перейдут сразу на гораздо более низкий субмикро-болитный уровень, что будет сопровождаться резким падением их продуктивности – с 300– 350 т/га до 50–70 т/га.
Прообразами их будущих функциональных состояний станут три фациальные группы: 1) лесолугово-степные комплексы солнцепечных склонов; 2) водораздельные сосново-широколиственные леса; 3) дубняки и осинники крутых нейтрально-теневых склонов. На своих южных окраинах массивы плакорных липняков будут поглощаться луговой степью, а на северных форпостах, где начинается глубокое эрозионное расчленение рельефа, – даже сухостепной растительностью.
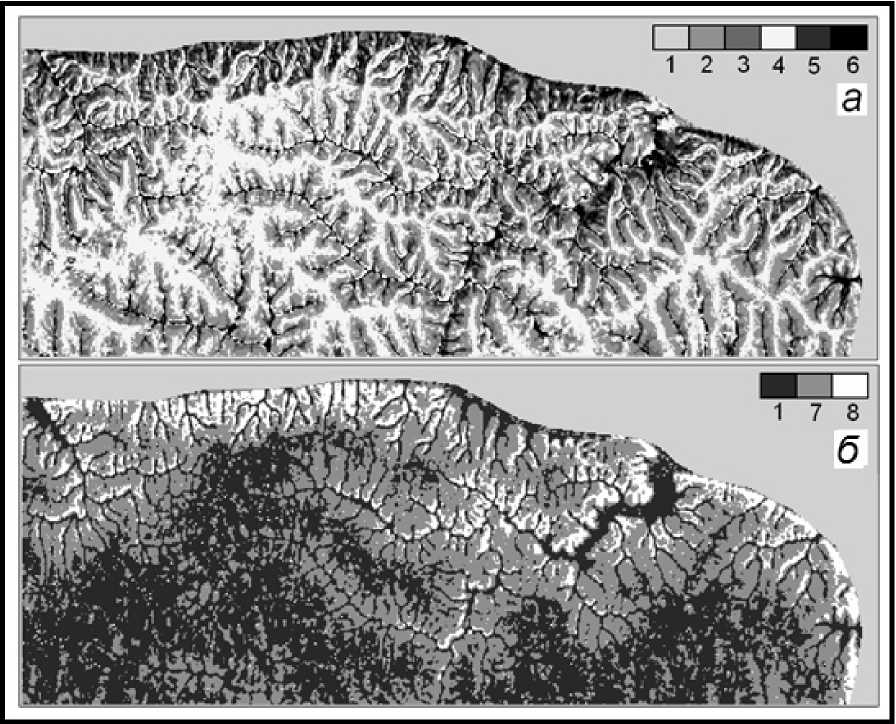
Рис. 5. Распределение фитоценозов в конце базового периода (а) и на прогнозный срок 2100 г. (б), согласно климатической модели HadCM3.
Карта рассчитана и составлена Л.С. Шарой.
Группы базовых фитоценозов: 1 – вязо-липняки и кленово-липовые осинники (элювиальные);
2 – редкостойные остепненные сосняки; 3 – сосняки сложные и сосново-широколиственные леса, 4 – дубняки и осинники; 5 – дубово-вязо-липовые леса и производные от них; 6 – липняки, кленовники и осинники; прогнозируемая растительность: 7 – южностепная; 8 – пустынно-степная.
Fig. 5. Distribution of phytocoenoses at the end of the base period ( a ) and for the forecast period of 2100 ( b ), according to the HadCM3 climate model.
The map was calculated and compiled by L.S. Sharaya.
Groups of basic phytocoenoses: 1 – viscous-lime and maple-linden aspen forests (eluvial); 2 – sparse steppe pine forests; 3 – compound pine forests and pine-broadleaf forests, 4 – oak and aspen forests;
5 – oak-elm-linden forests and their derivatives; 6 – lime, maple and aspen forests.
Predicted vegetation: 7 – southern steppe; 8 – desert-steppe.
На первых двух шагах (1996–2050 гг. и 2050– 2100 гг.) в лесо-лугово-степные комплексы заметно будут трансформироваться также дубо-липняки и осинники нижних частей и подножий теневых и нейтральных склонов (к 2100 г. по моделям GISS и HadCM3 соответственно на 25% и 52%). Одновременно до некоторой степени (на 10–20%) они приобретут облик сосновошироколиственных лесов. Однако в дальнейшем устойчивость дубняков возрастет, и они сохранят изменившийся вначале характер функционирования вплоть до конца ХХII столетия. В целом трансаккумулятивная мезоморфная группа лесных биогеоценозов окажется наиболее устойчивой во всем лесном покрове Жигулевского массива (их Pii = 0,69–0,91).
Гораздо более резкой трансформации подвергнутся лежащие выше по теневым и нейтральным склонам трансэлювиальные неморальнолесные фитоценозы (группа фаций 3).
Уже к 2050 г. почти на 50% они перейдут в функциональное состояние лесо-лугово-степных или даже северостепных формаций (соответственно по моделям GISS и HadCM3), а к концу прогнозного периода степень их преобразований в эти сообщества достигнет 70–80%.
Климатический сигнал наиболее сильно скажется на состоянии топоэкосистем семиаридной плеяды – лесо-лугово-степных комплексов солнцепечных склонов и водораздельных сосновошироколиственных лесов (групп фаций 1 и 2). Аридизация проявилась в структуре экологических ниш указанных биогеоценозов уже в конце базового периода, когда проводилась ландшафтная съемка (см. выше). Экологические доминанты термоэдафических ниш оказались сдвинутыми к максимальной температуре, а ниш гидро-эдафических – к минимальному влагосодержа-нию почвы. Очевидно, эти топоэкосистемы уже подверглись процессам климатической аридиза-ции и существенно утратили способность устойчиво функционировать при дальнейшем повышении летних температур и сокращении годовых осадков.
Климатическое остепнение фитоценозов семиаридной плеяды пойдет по нарастающей, причем согласно модели HadCM3 с заметно большей скоростью, нежели по модели GISS (см. рис. 3 и 4). Уже к 2050 г. остепненные редкостойные сосняки солнцепечных склонов сохранят уровень своего базового функционирования на 19% в первом случае и на 46% – во втором. В конце второго шага (2050–2100 гг.) на 80–90% они перейдут в состояние зональных групп северных или средних степей. В течение XXII столетия мера их Pii снизится до 0,081 и 0,043 соответственно. Согласно хэдловской модели, они на 96% превратятся функционально в опустыненные степи (см. рис. 4). Столь же интенсивному остепнению, а в дальнейшем и опустыниванию подвергнутся водораздельные сосново-широколиственные леса, несколько слабее – трансэлювиальные дубняки.
ЗАКЛЮЧЕНИЕ
Проведенный опыт изучения механизмов отображения глобальных биосферных процессов в структурно-функциональных характеристиках природных экосистем топологической размерности демонстрирует эффективность применения методов биогеоценологии к решению системных задач эколого-географического прогнозирования. Намечается один из эффективных путей разработки методов локального биоэкологического и геосистемного мониторинга в свете глобальных климатических изменений.
Работа выполнена при финансовой поддержке Российского фонда фундаментальных исследований, проект № 18-05- 00023-а.
Список литературы Влияние современного глобального потепления на лесные экосистемы Жигулевского массива (прогнозное эмпирико-статистическое моделирование)
- Албриттон Д.Л., Баркер Т., Башмаков И. и др. Изменение климата. 2001 г. Обобщенный доклад МГЭИК / Под ред. Р.Т. Уотсона. Geneva: World Meteorological Organization, 2003. 220 p.
- Андреев В.Л. Классификационные построения в экологии и систематике. М.: Наука, 1980. 142 c.
- Будыко М.И. Климат и жизнь. Л.: Гидрометеоиздат, 1971. 472 c.
- Высоцкий Г.Н. Избранные труды. М.: Сельхозгиз, 1960. 435 с.
- Израэль Ю.А., ред. Состояние и комплексныймониторинг природной среды и климата. Пределы изменений. М.: Наука, 2001. 242 с.
- Коломыц Э.Г. Региональная модель глобальных изменений природной среды. М.: Наука, 2003. 371 с.
- Коломыц Э.Г. Локальные механизмы глобальных изменений природных экосистем. М.: Наука, 2008. 427 с.
- Коломыц Э.Г., Розенберг Г.С., Колкутин В.И. и др. Экология ландшафтов Волжского бассейна в системе глобальных изменений климата (прогнозный Атлас-монография). Нижний Новгород: Интер-Волга, 1995. 165 с.
- Коломыц Э.Г., Юнина В.П., Сидоренко М.В. Воротников В.П. Экосистемы хвойного леса на зональной границе. Организация, устойчивость, антропогенная динамика. Н. Новгород: ИЭВБ РАН, 1993. 346 с.
- Меняющийся мир: географический подход к изучению. М.: Прогресс, 1991. 391 с.
- Одум Ю. Экология. В 2-х т. Пер. с англ. М.: Мир, 1986. Т. 1. 328 с. Т. 2. 376 с.
- Пузаченко Ю.Г., Скулкин В.С. Структура растительности лесной зоны СССР: Системный анализ. М.: Наука, 1981. 275 с.
- Спрыгин И.И. Материалы к познанию растительности Среднего Поволжья // Научное наследство. Т. 11. М.: Наука, 1986. 512 с.
- Сурова Н.А. Факторально-динамический анализ лесных экосистем и его прогнозное значение // Изв. Самар. НЦ РАН. 2015. Т. 17, № 4-1. С. 29-40.
- Швиденко А.З., Щепащенко Д.Г., Кракснер Ф., Онучин А.А. Переход к устойчивому управлению лесами России: теоретико-методические предпосылки // Сибирский лесной журнал. 2017. № 6. С. 3-25.
