Влияние стресса от засухи на структурные и функциональные характеристики Triticum spelta L.

Автор: Косакивска И.В., Васюк В.А., Бабенко Л.М., Войтенко Л.В.
Журнал: Журнал стресс-физиологии и биохимии @jspb
Статья в выпуске: 1 т.14, 2018 года.
Бесплатный доступ
Изучено влияние почвенной засухи на рост, микроструктуру поверхности листьев и состав фотосинтетических пигментов растений Triticum spelta L. 14-дневного возраста. Было показано, что корневая система, длина которой уменьшилась на 19%, а масса - на 48%, оказалась более чувствительной к воздействию 4-дневной почвенной засухи. На 23-й день после реабилитации разница между биометрическими параметрами контрольных и опытных образцов уменьшилась, но полного восстановления растений не произошло. Микроструктурный анализ амфистоматической листовой пластинки выявил одинаковое количество устьиц с одинаковыми размерами устьиц устья на адаксиальной и абаксиальной поверхности. После почвенной засухи наблюдалось некоторое увеличение плотности воска, устьица на обеих поверхностях листа оставалась закрытой. Пигментный комплекс 14-дневных растений ответил снижением количества хлорофилла и каротиноидов. На 23-й день после реабилитации наблюдалось дальнейшее снижение уровня фотосинтетических пигментов. Изменения соотношения содержания пигментов после стресса и особенностей микроструктуры поверхности листьев соответствовали засухоустойчивости T. spelta. Сохранение довольно высоких параметров содержания зеленого пигмента сразу после обезвоживания на 18-й день соответствовало данным биометрических исследований, демонстрирующих стабильный рост растения над наземной частью.
Короткий адрес: https://sciup.org/143165192
IDR: 143165192
Drought stress effects on Triticum spelta L. structural and functional characteristics
Effects of a soil drought on the growth, microstructure of leaf surface and photosynthetic pigments composition of Triticum spelta L. 14-days-old plants have been studied. It was shown that the root system whose length diminished by 19% and weight - by 48% turned out to be more sensitive to the impact of a 4-day soil drought. On the 23d day after rehabilitation the difference between biometric parameters of control and experiment samples reduced, but a complete recovery of plants did not occur. A microstructural analysis of the amphistomatic leaf lamina revealed the same number of stomata with similar sizes of stomatal pores on the adaxial and abaxial surface. Following the soil drought, some increase in wax density was observed, stomata on the both leaf surfaces remained closed. The pigment complex of 14-days-old plants responded by decreasing the quantity of chlorophyll and carotenoids. On the 23th day after rehabilitation a further decrease in the photosynthetic pigments level was observed. The changes in the ratio of pigments content after stress and peculiarities of microstructure of leaf surface corresponded with drought tolerance of T. spelta. The retention of rather high parameters of the green pigment content immediately after dehydration on the 18th day corresponded to biometric study data demonstrating a stable growth in the plant over ground part.
Текст научной статьи Влияние стресса от засухи на структурные и функциональные характеристики Triticum spelta L.
Пшеница принадлежит к главным продовольственным зерновым культурам мира. Важным элементом, определяющим ее урожайность, является создание устойчивых к биотическим и абиотическим стрессам сортов. Все виды пшеницы по морфологическим признакам разделяют на голозерные (настоящие) и пленчатые или полбяные (Жуковский, 1971). Гексаплоидный вид Triticum spelta L. принадлежит к пленчатым пшеницам, содержит 42 хромосомы и генетически близок к мягкой пшенице Triticum aestivum L. (Господаренко и др ., 2016). Как отдельный биологический вид T. spelta возникла на Ближнем Востоке в регионе «Плодородного полумесяца» вследствие спонтанного изменения в геноме Aegilops squarrosa (Господаренко и др ., 2016; Бабенко и др ., 2017). Спельта неприхотлива к условиям выращивания, имеет высокий коэффициент кущения, отличается скороспелостью и стекловидностью зерна, которое не осыпается и не повреждается вредителями, устойчива к переувлажнению, обладает холодо- и зимостойкостью (Дорофеев и др ., 1987). Зерно T. spelta содержит до 25% белка, что вдвое выше чем у T. aestivum , 18 незаменимых аминокислот, богата ненасыщенными жирными кислотами и клетчаткой (Шелепов и др. , 2004; Горн, 2008). Высокие пищевые качества и адаптированность к малозатратному органическому земледелию привлекают внимание к T. spelta во многих странах Европы (Твердохлеб, Богуславский, 2012).
Определяющими факторами, влияющими на процессы роста, развития и урожайность аграрных культур, являются температура и водный режим. Ранее нами были исследованы структурнофункциональные характеристики новых перспективных сортов озимой пшеницы при действии температурных стрессов (Babenko et al. , 2014; Kosakivska et al. , 2014; 2015). Целью настоящей работы было изучение влияния почвенной засухи на рост, микроструктуру поверхности листа и спектр фотосинтетических пигментов дикого сородича озимой пшеницы – T. spelta .
MATERIALS AND METHODS
Опыты проводили с 14-дневными растениями T. spelta сорта Франкенкорн. Сорт среднерослый, устойчив к полеганию, морозо- и засухоустойчивый, экологически пластичный (Schmitz, 2006). Семена были получены из коллекции Национального центра генетических ресурсов растений Украины (г. Харьков).
Откалиброванные семена стерилизовали в 80% растворе этилового спирта, промывали дистиллированной водой и замачивали на 3 часа. Затем проращивали в кюветах на смоченной водой фильтровальной бумаге в термостате при температуре +240C в течение 21 ч. Проклюнувшиеся семена высаживали в сосуды емкостью 2 л. В качестве субстрата использовали прокаленный речной песок. Растения выращивали при температуре 20/17°С (день/ночь), интенсивности освещения 690 мкмоль/(м2·с), фотопериод составлял 16/8 ч. (день/ночь), относительная влажность воздуха 65 + 5%, влажность субстрата поддерживали на уровне 60% от полной влагоемкости. Полив проводили ежедневно раствором Кнопа из расчета по 50 мл на сосуд. Почвенную засуху создавали, прекращая полив 14дневных растений на четыре дня до увядания листьев и снижения влагоемкости субстрата вдвое. В фазу 2-3 листьев (на 18 день) полив восстанавливали, рост растений происходил в нормальных условиях (фаза 2-4 листьев) до 23 дня. Длину и вес надземной части и корней определяли на 14, 18 и 23 день. Микроструктуру листовой поверхности исследовали на электронном микроскопе JEOL JSM-6060 LA (Япония). Подготовку образцов для анализа проводили согласно (Щербатюк и др., 2015). Размеры структур на микрофотографиях определяли, используя программу UTHSCSA Image Tool 3.0. Фотосинтетические пигменты экстрагировали 80% ацетоном и определяли согласно (Wellburn, 1987). Опыты проводили в двух биологических и трех аналитических повторах. Для каждого биологического повтора отбирали по 40 растений.
Статистическую обработку результатов проводили, используя t-тест Стьюдента, статистически достоверной считали разницу при р≤ 0.05.
RESULTS AND DISCUSSION
Засухоустойчивость – сложный полигенный признак, который определяется адаптивными, генетически обусловленными свойствами растения (Fischer, 2011). Показано, что скорость потери воды зависит от анатомического строения, интенсивности метаболических процессов и возраста органа растения (Alves, Setter, 2004). При водном дефиците из-за нарушения системы регуляции вода активно поступает в листья, тогда как корневая система обезвоживается. Однако из-за отмирания корневых волосков функциональная активность корневой системы после реабилитации полностью не восстанавливается (Das et al. , 2015)
Нами было показано, что почвенная засуха, вызванная прекращением полива в течение 4 дней, замедляла рост растений T. spelta. Более чувствительной к недостатку влаги оказалась корневая система, длина которой уменьшилась на 19%, а вес – на 48%, тогда как изменения в надземной части были менее выраженными. В период реабилитации после возобновления полива разница между биометрическими показателями контроля и опыта уменьшилась, однако полного восстановления у растений, которые подверглись воздействию засухи, не произошло (рис. 1).
Листовая амфистоматическая пластинка T. spelta имеет ярко-зеленую окраску. На адаксиальной и абаксиальной эпидерме расположены трихомы двух типов: «хуки» и длинные игловидные трихомы (рис. 2А, Б). Эпидерма покрыта слоем воска, представленным двумя видами кристаллов: пластинками и тубусами (рис. 2В, Г). Тубусы расположены в основном в зоне устьиц (рис. 2Д). К первичным функциям кутикулярных восков относятся защита от чрезмерной инсоляции и уменьшение испарения при водном дефиците (Yeats, Rose, 2013).
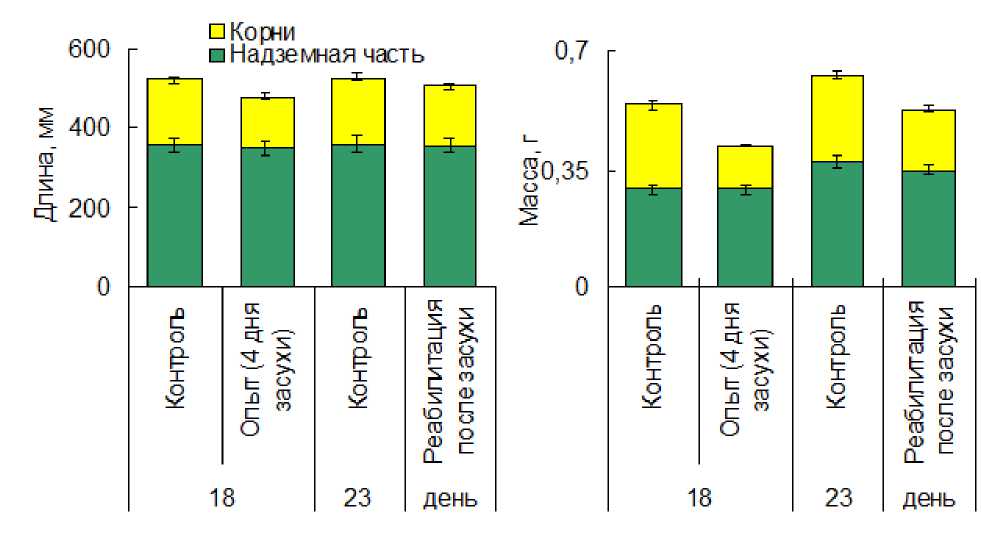
Figure 1. Влияние почвенной засухи на биометрические показатели Triticum spelta.
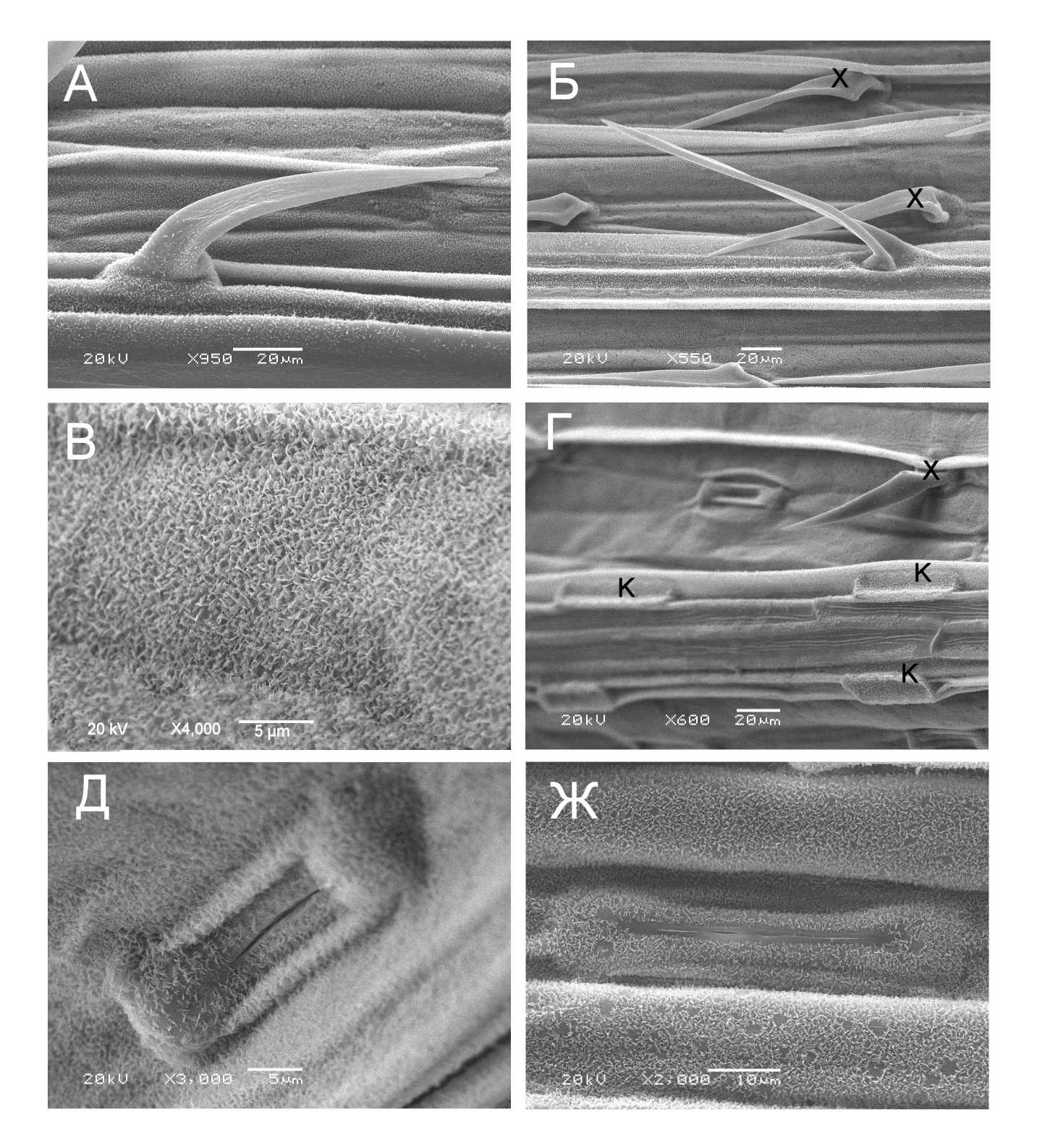
Figure 2. Микроструктура эпидермиса листа Triticum spelta в контрольных условиях (А–Д) и в условиях почвенной засухи (Ж). А – внешний вид трихомы типа «хук»; Б – внешний вид игловидной трихомы.; В – слой восковых пластинок на адаксиальной поверхности листа; Г – общий вид адаксиальной поверхности листа; Д – устьице в контроле; Ж – устьице в условиях почвенной засухи. К – кремниевые клетки; Х – «хуки»
Table 1. Влияние почвенной засухи на соотношение фотосинтетических пигментов Triticum spelta
|
Образец |
а/b |
a+b/ каротиноиды |
|
Контроль, 18 день |
2,89 |
4,86 |
|
Опыт, 18 день |
3,23 |
5,00 |
|
Контроль, 23 день |
2,81 |
5,18 |
|
Реабилитация после засухи, 23 день |
3,83 |
6,19 |
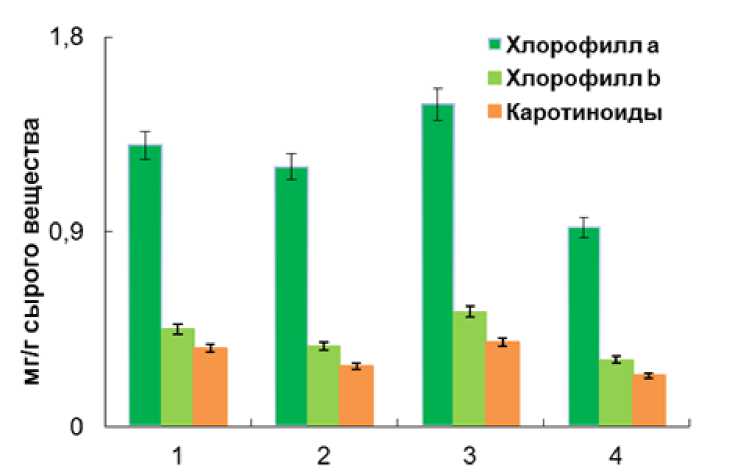
Figure 3. Влияние почвенной засухи на содержание фотосинтетических пигментов у растений Triticum spelta . 1 – 18-дневные растения контроль; 2 – 18-дневные растения после 4-дневной почвенной засухи; 3 – 23-дневные растения контроль; 4 – восстановление после почвенной засухи, 23 день.
Эпидермальная ткань состоит преимущественно из длинных извилистых кремниевых клеток (рис. 2Г). Устьица парацитного типа расположены на обеих поверхностях листовой пластинки на одном уровне с основными эпидермальными клетками и ориентированы вдоль проводящих пучков (рис. 2Г). Количество устьиц в межреберной зоне на 1 мм2 составляет 64,44 ± 7,19 шт., Площадь устьица равна 42,23 ± 2,14мкм2, длина устричной щели – 10,91 ± 0,64 мкм. В условиях почвенной засухи характерные признаки эпидермиса листовой пластинки T. spelta сохранялись. Устьица на обеих поверхностях листовой пластинки оставались закрытыми (рис. 2Ж).
Успешность адаптации в значительной степени зависит от оптимального функционирования ассимиляционного аппарата, одним из показателей состояния которого является содержание фотосинтетических пигментов (Андрианова, Тарчевский, 2000). Соотношение хлорофиллов а/b рассматривается как один из признаков фотосинтетической активности, а при стрессах используется в качестве маркера устойчивости (Косаковская и др., 2014; Babenko et al., 2014)
В условиях почвенной засухи у растений T. spelta зафиксировано незначительное уменьшение количества хлорофиллов a и b. Однако после восстановления на 23 день произошло дальнейшее снижение уровня хлорофиллов а (в 1,7 раза) и b (в 1.3 раза), а также существенное уменьшение содержания каротиноидов (рис. 3). Соотношения между хлорофиллами а и b после почвенной засухи и в период реабилитации возросло. Также у опытных растений на 23 день значительно увеличилось соотношение суммы хлорофиллов к каротиноидам (табл.).
В работах других авторов сообщалось, что засуха негативно влияла на содержание хлорофиллов у разных видов растений (Guerfel et al. , 2009; Mafakheri et al. , 2010; Zhang, Kirkham, 2016). Так, установлено значительное снижение количества пигментов у шести сортов T. aestivum (Nyachiro, 2001). Уровень хлорофилла а во флаговом листе пшеницы используется в качестве маркера при скрининге на устойчивость к засухе (Arjenaki et al. , 2012). Полученные нами данные продемонстрировали, что пигментный комплекс активно реагировал на засуху, а отмеченные изменения в соотношениях между пигментами указывают на устойчивость спельты к почвенной засухе. Сохранение достаточно высоких показателей в содержании зеленых пигментов после обезвоживания соответствовало данным биометрических исследований, продемонстрировавшим стабильность ростовых процессов в надземной части растений.
CONCLUSION
В современном производстве пшеницы четко обозначились тенденции, сосредоточенные на возрождении, селекции и внедрении в производство забытых региональных зерновых культур, так называемых «античных злаков», обладающих ценными хозяйственными свойствами. Одним из таких злаков является T. spelta. В результате проведенного исследования нами было установлено, что более чувствительной к воздействию засухи была корневая система растений T. spelta, длина которой уменьшилась на 19%, а вес – на 48%. В период реабилитации на 23 день разница между биометрическими показателями контроля и опыта уменьшилась, однако полного восстановления у растений не произошло. Микроструктурный анализ амфистоматической листовой пластинки выявил одинаковое количество устьиц с близкими значениями размеров устьичных щелей на адаксиальной и абаксиальной поверхностях. После почвенной засухи наблюдалось увеличение плотности воска, устьица на обеих поверхностях листовой пластинки оставались закрытыми.
Пигментный комплекс 14-дневных растений реагировал на засуху уменьшением количества хлорофиллов и каротиноидов. В период реабилитации на 23 день зафиксировано дальнейшее снижение уровня фотосинтетических пигментов. Сохранение достаточно высоких показателей в содержании зеленых пигментов сразу после обезвоживания на 18 день соответствовало данным биометрических исследований, продемонстрировавшим стабильность ростовых процессов в надземной части растений. Изменения в соотношении между пигментами, зафиксированные после обезвоживания, а также особенности микроструктуры листовой поверхности коррелируют с устойчивостью спельты к почвенной засухе.
Список литературы Влияние стресса от засухи на структурные и функциональные характеристики Triticum spelta L.
- Андрианова Ю.Е., Тарчевский И.А. (2000) Хлорофилл и продуктивность растений. М.: Наука, 135 с
- Бабенко Л.М., Рожков Р.В., Парiй Я.Ф., Парiй М.Ф., Водка М.В. Косакiвська I.В. (2017) Triticum dicoccum (Schrank) Schuebl.: походження, бiологiчна характеристика й перспективи використання в селекцiї та сiльському господарствi. Вiсник Харкiв. нацiон. агр. ун-ту., 2 (41), с. 92-102
- Горн Е. (2008) Лучше чем пшеница, но… Фермерське господарство, 4 (372), 21 с
- Господаренко Г.М., Костогриз П.В., Любич В.В., Парiй М.Ф., Полторецький I.О. (2016) Пшениця спельта. К.: Стiк груп Україна, 300 с
- Дорофеев В.Ф., Удачин Р.А., Семенова Л.В. (1987) Пшеницы мира. Л.:ВО Агропромиздат, 560 с
- Жуковский П.М. (1971) Культурные растения и их сородичи. Ленинград: Колос, 752 с
- Косакiвська I.В., Бабенко Л.М., Скатерна Т.Д., Устинова А.Ю. (2014) Вплив гiпо-i гiпертермiї на активнiсть лiпоксигенази, вмiст пiгментiв i розчинних бiлкiв у проростках Triticum aestivum L. сорту Ятрань 60. Физиол. раст. и генетика, 46(3), с. 212-220
- Твердохлiб О.В., Богуславський Р.Л. (2012) Видове рiзноманiття пшеницi, напрямки i перспективи його використання. Зб. наук. праць Уманськ. нац. ун-ту садiвництва, Вып. 80, ч. 1, с. 37-47
- Шелепов В.В., Маласай В.М., Пензев А.Ф. и др. (2004) Морфология, биология, хозяйственная ценность пшеницы. Мироновка, 524 с
- Щербатюк M.M., Бриков В.O., Мартин Г.Г. (2015) Пiдготовка зразкiв рослинних тканин для електронної мiкроскопiї (теоретичнi та практичнi аспекти). К.: Талком, 152 с
- Alves A.C., Setter T.L. (2004) Response of cassava leaf area expansion to water deficit: cell proliferation, cell expansion and delayed development. Annals of Botany, 94, р. 605-613
- Arjenaki G., Jabbari R., Morshedi A. (2012) Evaluation of drought stress on relative water content, chlorophyll content and mineral elements of wheat (Triticum aestivum L.) varieties. Int. J. Agricult. Crop Sci., 4 (11), р 726-729
- Babenko L., Kosakivska I., Akimov Yu., Klymchuk D., Skaternya T. (2014) Effect of temperature stresses on pigment spectrum, lipoxygenase activity and cell ultrastructure of winter wheat seedlings. Genetics and plant physiology, 4 (1-2), р. 117-125
- Das R., Bhagawati K., Boro A., Medhi T., Medhi B., Bhanisanar K. (2015) Relative performance of plant cultivars under respective water deficit adaptation strategies: A case study. Current World Environment, 10, р. 683-690
- Fischer R.A. (2011) Wheat Physiology: A review of recent developments. Crop and Pasture Science, 62(2), р. 95-114
- Guerfel M., Baccouri O., Boujnah D., Chaibi W., Zarrouk M. (2009) Impacts of water stress on gas exchange, water relations, chlorophyll content and leaf structure in the two main Tunisian olive (Olea europaea L.) cultivars. Sci. Horticult., 119, р. 257-263
- Hsiao T.C., Xu L. (2000) Sensitivity of growth of roots versus leaves to water stress: biophysical analysis and relation to water transport. J. of Exp. Bot. 51(350), р. 1595-1616
- Kosakivska I.V., Voytenko L.V., Likhnyovskiy R.V., Ustinova A.Y. (2014) Effect of temperature on accumulation of abscisic acid and indole-3-acetic acid in Triticum aestivum L. seedlings. Genetics and Plant Physiol. 4(3-4), р. 201-208
- Kosakivska I.V., Voytenko L.V., Likhnyovskiy R.V. (2015) Effect of temperature on Triticum aestivum L. seedlings growth and phytohormone balance. J. of Stress Physiology & Biochemistry, 11(4), р. 91-99
- Mafakheri A., Siosemardeh A., Bahramnejad B., Struik P.C., Sohrabi Y. (2010) Effect of drought stress on yield, proline and chlorophyll contents in three chickpea cultivars. Austr. J. Croup Sci. 4(8), р. 580-585
- Schmitz K. (2006) Dinkel -ein Getreide mit Zukunft. Backmittelinstitut aktuell.,-Sonderausgabe, P. 1-8
- Wellburn A. (1994) The spectral determination of chlorophyll a and chlorophyll b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol., 144, р. 307-313
- Yeats T.H., Rose J.K. (2013) The formation and function of plant cuticles. Plant Physiol. 163, р. 5-20
- Zhang J., Kirkham M.B. (2016) Antioxidant response to drought in sunflower and sorghum seedlings. New Phytol. 132, р. 361-373
