Влияние техногенного загрязнения на динамику роста и мезоструктуру листьев Betula czerepanovii Orlova (Мурманская область)
")
Автор: Василевская Наталья Владимировна, Лукина Юлия Михайловна
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Биология
Статья в выпуске: 8 (121), 2011 года.
Бесплатный доступ
Тяжелые металлы, береза черепанова, рост, лист, мезоструктура
Короткий адрес: https://sciup.org/14750038
IDR: 14750038
Текст статьи Влияние техногенного загрязнения на динамику роста и мезоструктуру листьев Betula czerepanovii Orlova (Мурманская область)
Техногенное загрязнение среды лимитирует продукционный процесс, что приводит к нарушению метаболизма растений, торможению их роста и развития, снижению продуктивности и продолжительности вегетации. Для оценки антропогенного воздействия с 60-х годов ХХ века начали использовать методы фитоиндикации [10], [14]. Накоплен большой фактический материал мониторинга техногенной среды с использованием различных критериев, таких как интенсивность физиологических процессов, уровень развития морфологических и анатомических структур растений [9], [10], [23], [25], [34].
Мурманская область является одним из центров цветной металлургии на Северо-Западе России. Вокруг медно-никелевых комбинатов «Североникель» и «Печенганикель» проводится регулярный мониторинг состояния наземных экосистем, растительных популяций и сообществ, их продукционных характеристик [17], [18], [20], [21], [32], [36]. В то же время работы по изучению антропогенного воздействия на фотосинтетический аппарат листа, в частности его анатомическую структуру, немногочисленны и выполнены в основном на хвойных [8], [29], [30].
Цель исследования – анализ влияния техногенного загрязнения на динамику роста и анатомическую структуру листьев березы Черепанова ( Betula czerepanovii Orlova) в окрестностях комбината «Североникель».
МАТЕРИАЛЫ И МЕТОДЫ
Исследование роста и развития листьев Betula czerepanovii проводили в полевой сезон 2006 года в окрестностях медно-никелевого комбина-
та «Североникель» (г. Мончегорск). Основными загрязняющими веществами при переработке сырья являются диоксид серы и пыль, содержащая тяжелые металлы (Ni, Cu, Co). В 2006 году валовые выбросы загрязняющих веществ в атмосферу превысили 46 492 т. Выбросы сернистого ангидрида (SO2) в 2006 году составили 40,3 т, никеля – 456,5 т, меди – 599,6 т [7]. Техногенная пустыня, где доля погибших деревьев составляет более 80 %, протянулась на 10 км в южном направлении от комбината и на 15 км – в северном [32]. Исследования проводили на пробных площадях, расположенных по градиенту загрязнения в юго-западном направлении от комбината. Экспериментальные площади заложены в растительных сообществах, находящихся на разных стадиях деградации: техногенная пустошь (3 км – ПП1), техногенное редколесье (8 км – ПП2), дефолиирующие леса (20 км – ПП3, 30 км – ПП4), фоновая зона (60 км – ПП5). На каждой площадке маркировали по 10 деревьев B. czerepanovii, близких по габитусу, с диаметром ствола 10–15 см, высотой 3–5 м. Исследования динамики роста листьев проводили в течение июня – июля 2006 года по стандартной методике [12]. Замеры листьев (длины и ширины) проводили каждые 10 суток. Для проведения анатомических исследований в начале августа 2006 года с маркированных деревьев на каждой площадке собрано и зафиксировано по 20 листьев. В качестве фиксатора использован FAA [2]. Изучение мезоструктуры листьев B. cze-repanovii проводили на поперечных срезах под световым микроскопом Биомед С-2 с помощью окулярмикрометра. Исследованы следующие показатели: число слоев, длина и ширина клеток палисадной и губчатой ткани, толщина палисадного и губчатого мезофилла и листовой пластинки. Объем выборки составил 100 измерений по каждому параметру. Микрофотосъемку проводили при помощи цифровой фотокамеры «Canon Ixus 800 IS» на микроскопе Биомед С-2.
Концентрацию тяжелых металлов (медь, никель) в листьях B. czerepanovii определяли методом атомно-адсорбционной спектрофотометрии в лаборатории наземных экосистем Института проблем промышленной экологии Севера КНЦ РАН. В качестве контроля концентрации тяжелых металлов в листьях березы использованы данные по содержанию никеля и меди, полученные на фоновой площади в пос. Алакуртти (250 км от г. Мончегорска), которая соответствует критериям, принятым в международной практике [24] (табл. 1). Для статистической обработки данных использован пакет программ Microsoft Exсel-2000.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Содержание тяжелых металлов в листьях. В результате проведения анализа на содержание в листьях B. czerepanovii ионов никеля и меди показано, что концентрация тяжелых металлов значительно увеличивается по градиенту загрязнения (табл. 1).
Таблица 1
Содержание меди и никеля в листьях Betula czerepanovii в окрестностях комбината «Североникель» (2006 год)
|
№ пробной площади |
Ni (мг/кг) |
Cu (мг/кг) |
|
ПП1 |
191,61 |
39,393 |
|
ПП2 |
169,33 |
50,156 |
|
ПП3 |
79,581 |
28,603 |
|
ПП4 |
45,626 |
19,00 |
|
ПП5 |
26,887 |
12,911 |
|
Контроль |
4,51 |
4,98 |
Максимальное содержание ионов никеля в листьях березы выявлено в зонах техногенного редколесья (ПП2) и техногенной пустоши (ПП1), где превышение составляет 38 и 42 раза соответственно (табл. 1). Концентрация меди в фотосинтезирующих органах на данных площадках значительно меньше и превышает контрольные значения в 8 и 10 раз. Полученные данные согласуются с результатами других авторов [1], [4], [17], [18], [19], [27], проводивших химический анализ листьев различных видов растений, которые подвергались воздействию техногенного загрязнения комбината «Североникель», и выявивших в них многократное увеличение содержания ионов никеля и меди.
Рост и развитие листьев. В ходе исследований установлено, что высокие концентрации ни- келя и меди в листьях B. czerepanovii влияют на процессы роста и развития. Так, на пробных площадках в фоновой зоне (ПП5) и зоне дефоли-ирующих лесов (ПП3, ПП4) развертывание листьев началось на 5 суток ранее (22.05.2006), чем у деревьев, произрастающих на площадках в окрестностях комбината (ПП2, ПП1). Нарушение динамики развития на ранних этапах онтогенеза листа свидетельствует о токсичном воздействии тяжелых металлов на процессы дифференцировки меристематических клеток. В условиях техногенной нагрузки выявлено ингибирование апикального (рис. 1) и маргинального роста листовых пластинок B. czerepanovii (рис. 2). Если в зоне дефолиирующих лесов (ПП4) показатели длины листа незначительно отличаются от фоновых значений (ПП5), то в окрестностях комбината эффект ингибирования усиливается на экспериментальных площадках ПП3, ПП2, ПП1 (рис. 1), особенно в первые дни вегетации. Длина листовых пластинок в зонах техногенного редколесья (ПП2) и пустоши (ПП1) в начале их развертывания (31.05.2006) на 40 % меньше, чем в фоновой зоне (ПП5). Через 20 дней после начала вегетации эти различия на всех площадках снижаются до 20–30 % и сохраняются в течение дальнейшего периода роста листьев.
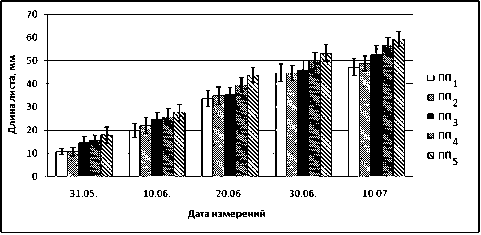
Рис. 1. Влияние техногенного загрязнения на апикальный рост (длину листовой пластинки) B. czerepanovii в окрестностях комбината «Североникель» (мм) в вегетационный сезон 2006 года
Установлено, что высокая концентрация ионов никеля и меди в листьях B. czerepanovii вызывает большее ингибирование маргинального роста листовых пластинок, чем апикального, особенно на ранних этапах онтогенеза (рис. 2). В начале развертывания ширина листа в зоне техногенного редколесья (ПП2) составляет 40 % (31.05.2006) и 20 % (10.06.2006) от фоновых значений, на техногенной пустоши (ПП1) этот показатель равен 45 и 38 % соответственно.
Как показывают исследования многих авторов [6], [22], [23], [31], фотосинтетический аппарат очень чувствителен к повышению содержания тяжелых металлов в окружающей среде, что проявляется в нарушении многих параметров его функционирования. Согласно нашим данным, ингибирование апикального и маргинального роста листьев B. czerepanovii в окрестностях комбината «Североникель» прежде всего обусловлено высокими концентрациями ионов никеля и меди. Полученные результаты подтверждают исследования других авторов [11], [16], [23], [28], [35], [37], [38], которые также показали, что повышение концентрации тяжелых металлов в среде приводит к значительному уменьшению длины, ширины и площади листовой пластинки, что является одной из причин снижения интенсивности процесса фотосинтеза.
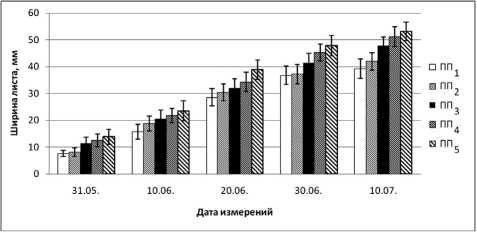
Рис. 2. Влияние техногенного загрязнения на маргинальный рост (ширину листовой пластинки) B. czerepanovii в окрестностях комбината «Североникель» (мм) в вегетационный сезон 2006 года
Особый интерес представляет высокий уровень ингибирования роста листа в начале его онтогенеза. Этот эффект может быть вызван токсичным воздействием ионов никеля, которые подавляют дифференциацию клеток меристем, функционирующих в первые 10 дней развития листа, то есть в период начала формирования листовой пластинки [26]. При этом, как показали наши данные, наиболее выражено ингибирование маргинального роста, что, по-видимому, объясняется более токсичным воздействием тяжелых металлов на деление клеток маргинальных меристем, чем апикальных. По мнению Б. Р. Васильева, длина и ширина листовой пластинки изменяются независимо друг от друга, что свидетельствует об отсутствии сопряженности между апикальным и маргинальным ростом зачатка листа, а также о слабой выраженности диффузного роста в процессе развития листа древесных растений [3].
Мезоструктура листа. В результате исследований анатомической структуры листьев B. czerepanovii выявлено, что в условиях фоновой зоны лист имеет крупноклеточную эпидерму с сильно утолщенной наружной клеточной оболочкой (рис. 3). Отчетливо выражена дифференциация мезофилла на палисадную и губчатую ткани. Палисадный мезофилл состоит из одного ряда вытянутых узких клеток. Очень рыхлая, с крупными межклетниками губчатая ткань состоит из 5–6 рядов клеток изодиаметри-ческой формы. Клетки нижней эпидермы сильно различаются между собой по форме и размеру, их клеточные стенки несколько утолщены.
В листьях B. czerepanovii, произрастающей в условиях техногенной нагрузки, число слоев палисадной ткани варьирует по градиенту загряз- нения от 1 до 2. Индекс палисадности мезофилла листьев на всех экспериментальных площадках, кроме техногенной пустоши, варьирует от 0,6 до 0,7 (табл. 2), тогда как на площадке ПП1 этот показатель имеет максимальное значение – 0,9.
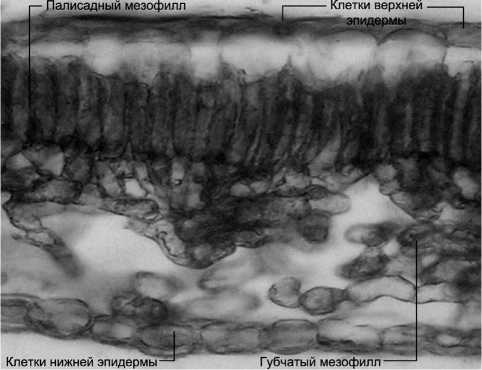
Рис. 3. Анатомическая структура листа B. czerepanovii на техногенной пустоши в окрестностях комбината «Североникель»
Палисадные клетки листа значительно удли- няются по мере увеличения концентрации тяжелых металлов (в зоне дефолиирующих лесов (ПП3, ПП4) – на 45 %, техногенного редколесья (ПП2) – на 60 %, техногенной пустоши (ПП1) – на 100 %) и уменьшаются в диаметре. Соотношение
длины палисадной клетки к ее ширине возрастает по градиенту загрязнения от 1,9 (ПП5, ПП4) до 3 (ПП3), 4,2 (ПП2) и 6,6 (ПП1). Толщина палисадного мезофилла листьев также увеличивается за счет роста клеток в длину (табл. 2).
Число слоев губчатой ткани листа B. cze-repanovii , произрастающей в зоне техногенного воздействия комбината, варьирует в пределах ошибки от 5 до 6 (табл. 2). В то же время изменяются количественные параметры клеток губчатого мезофилла. Длина губчатых клеток листьев березы в зоне дефолиирующих лесов увеличивается на 48 % (ПП3), техногенного редколесья (ПП2) – на 38 %, техногенной пустоши (ПП1) – на 55 % по сравнению с фоновыми значениями (табл. 2). Достоверно уменьшается диаметр клеток губчатого мезофилла листьев березы, находящихся в непосредственной близости к комбинату (ПП2, ПП1). За счет удлинения губчатых клеток увеличивается толщина губчатого мезофилла по градиенту загрязнения (табл. 2).
По мере возрастания техногенной нагрузки и увеличения концентрации ионов никеля и меди увеличивается толщина листовой пластинки. Если в зоне дефолиирующих лесов (ПП4) этот показатель изменяется незначительно, то в зонах техногенного редколесья (ПП2) и пустоши (ПП1) происходит утолщение листа на 65 % по сравнению с фоновой зоной.
Таблица 2
Влияние аэротехногенного загрязнения на анатомическую структуру листьев Betula czerepanovii в окрестностях комбината «Североникель» (2006 год)
|
Параметры (n = 50) |
ПП1 |
ПП2 |
ПП3 |
ПП4 |
ПП5 |
|
Толщина листовой пластинки (мкм) |
25,89 ± 1,69 |
25,63 ± 1,52 |
23,54 ± 1,43 |
18,36 ± 1,09 |
16,93 ± 1,74 |
|
Толщина палисадного мезофилла (мкм) |
9,79 ± 1,40 |
7,73 ± 1,52 |
7,10 ± 1,06 |
5,58 ± 0,96 |
5,89 ± 0,76 |
|
Число слоев палисадного мезофилла |
1,76 ± 0,34 |
1,39 ± 0,31 |
1,45 ± 0,36 |
1,32 ± 0,27 |
1,29 ± 0,23 |
|
Толщина губчатого мезофилла (мкм) |
11,15 ± 1,64 |
10,96 ± 1,62 |
10,28 ± 1,06 |
8,91 ± 1,02 |
8,67 ± 1,62 |
|
Число слоев губчатого мезофилла |
5,03 ± 0,84 |
5,57 ± 0,76 |
5,96 ± 0,58 |
5,24 ± 0,65 |
5,57 ± 0,82 |
|
Длина палисадной клетки (мкм) |
4,59 ± 0,86 |
3,76 ± 0,65 |
2,93 ± 0,64 |
2,20 ± 0,52 |
2,27 ± 0,47 |
|
Диаметр палисадной клетки (мкм) |
0,70 ± 0,2 |
0,90 ± 0,36 |
0,95 ± 0,22 |
1,13 ± 0,35 |
1,22 ± 0,46 |
|
Длина губчатой клетки (мкм) |
2,33 ± 0,34 |
2,07 ± 0,46 |
2,22 ± 0,46 |
1,89 ± 0,49 |
1,51 ± 0,38 |
|
Диаметр губчатой клетки (мкм) |
0,92 ± 0,33 |
1,18 ± 0,29 |
1,33 ± 0,36 |
1,35 ± 0,28 |
1,42 ± 0,39 |
|
Индекс палисад-ности |
0,9 |
0,7 |
0,7 |
0,6 |
0,7 |
|
ДПМ/ ШПМ* |
6.56 |
4,2 |
3 |
1,9 |
1,9 |
*Соотношение длины клетки палисадного мезофилла к ширине клетки.
Тяжелые металлы, в частности никель, вызывают анатомические изменения органов растений [26]. Существуют различные представления о том, какие именно ткани листа подвержены наибольшему воздействию. По мнению одних авторов, это палисадная паренхима [11], по мнению других – губчатая [13]. Согласно полученным нами данным, ионы никеля и меди вызывают более значительную модификацию палисадных клеток листьев B. czerepanovii , выражающуюся в их удлинении, уменьшении диаметра, увеличении индекса палисадности, усилении периклинальных делений. В меньшей степени изменяется величина губчатых клеток. Аналогичные данные получены И. В. Серегиным и А. Д. Кожевниковой [26] в лабораторных условиях на видах Triticum aestivum , Brassica oleracea .
Значительное утолщение листовых пластинок B. czerepanovii в окрестностях комбината
«Североникель» может быть вызвано интегральным воздействием тяжелых металлов. Имеются данные, что ионы меди обусловливают 5 % увеличение средней суммарной толщины листьев [15]. При этом изменения как в апикальной, так и в срединной и базальной частях происходят за счет увеличения толщины столбчатой и губчатой паренхимы. Значительный рост листа в толщину при уменьшении площади поверхности выявлен также у Vaccinium myrtillus в окрестностях завода по переработке твердых бытовых отходов г. Мурманска [33].
Увеличение размеров клеток мезофилла и толщины листа, по-видимому, является неспецифической реакцией B. czerepanovii на техногенное загрязнение среды и, в частности, высокую концентрацию ионов никеля и меди. Аналогичные данные получены Ю. В. Гамалеем [5] для рода Betula в экстремальных условиях произрастания, что, по нашему мнению, свидетельствует о неспецифической реакции ассимиляционного аппарата берез на действие стрессовых факторов.
ЗАКЛЮЧЕНИЕ
Техногенное загрязнение комбината «Северо-никель», в частности высокое содержание ионов никеля и меди в листьях B. czerepanovii , приводит к ингибированию их апикального и маргинального роста, особенно в первые дни вегетации. Установлено, что маргинальные меристемы листа более подвержены воздействию тяжелых металлов, чем апикальные. Тяжелые металлы вызывают более значительную модификацию клеток палисадного мезофилла листьев B. czerepanovii , чем губчатого, что выражается в их удлинении, уменьшении диаметра, увеличении индекса палисадности и соотношения длины палисадной клетки к ее ширине, усилении периклинальных делений. В зонах техногенного редколесья и пустоши длина клеток палисадного мезофилла листьев B. czerepanovii увеличивается на 60 и 100 % соответственно, при этом уменьшается их диаметр. По мере возрастания техногенной нагрузки листовая пластинка B. сzerepanovii при общем снижении фотосинтетической поверхности значительно утолщается в большей степени в связи с увеличением объема палисадного мезофилла. Возможно, что в условиях стресса достигается компенсационный эффект за счет увеличения внутренней фотосинтетической поверхности мезофилла листа.
БЛАГОДАРНОСТЬ
Авторы выражают благодарность сотрудникам лаборатории наземных экосистем Института проблем промышленной экологии Севера КНЦ РАН за проведение анализа листьев B. czerepanovii на содержание тяжелых металлов методом атомно-адсорбционной спектрофотометрии.
Список литературы Влияние техногенного загрязнения на динамику роста и мезоструктуру листьев Betula czerepanovii Orlova (Мурманская область)
- Баркан В. Ш. Загрязнение почвы никелем и медью от промышленного источника металлургических пылей//Экологические проблемы северных регионов и пути их решения. Апатиты: Изд-во КНЦ РАН, 2008. С. 46-50.
- Барыкина Р. П. Справочник по ботанической микротехнике. Основы и методы. М.: Изд-во МГУ, 2004. 312 с.
- Васильев Б. Р. Строение листа древесных растений различных климатических зон. Л.: Изд-во ЛГУ, 1988. 208 с.
- Влияние промышленного атмосферного загрязнения на сосновые леса Кольского полуострова/Под ред. Б. Н. Норина и др. Л.: Изд-во БИН АН СССР, 1990. 195 с.
- Гамалей Ю. В. Транспортная система сосудистых растений. СПб.: Изд-во СПбГУ, 2004. 422 с.
- Гетко Н. В. Растения в техногенной среде: структура и функции ассимиляционного аппарата. Минск: Наука и техника, 1989. 208 с.
- Ежегодник Кольской горно-металлургической компании. М., 2006. 47 с.
- Ефимова М. А. Биоморфологические особенности Vaccinium myrtillus L. и V. vitis-idaea L. в естественных и антропогенно нарушенных лесных сообществах Кольского полуострова: Дис. … канд. биол. наук. СПб., 2007.
- Жиров В. В. Адаптации и возрастная изменчивость растений на Севере. Ч. 1/2. Апатиты: Изд-во КНЦ РАН, 2001. 355 с.
- Жиров В. К., Голубеева Е. И., Говорова А. Ф., Хаитбаев А. Х. Структурно-функциональные изменения растительности в условиях техногенного загрязнения на Крайнем Севере. М.: Наука, 2007. 162 с.
- Илькун Г. М. Загрязнители атмосферы и растения. Киев: Наукова думка, 1978. 246 с.
- Кищенко И. Т. Методика изучения сезонного роста древесных растений//Методы полевых и лабораторных исследований растений и растительного покрова. Петрозаводск: Изд-во ПетрГУ, 2001. С. 178-184.
- Красинский Н. П. Дымоустойчивость растений и дымоустойчивые ассортименты. М.: Наука, 1950. С. 9-109.
- Крюч ков В. В., Кондратович И. И., Андреев Г. Н. Красная Книга экосистем Кольского Севера. Апатиты: Изд-во КНЦ РАН, 1988. 102 с.
- Кулагин А. А., Шагиева Ю. А. Древесные растения и биологическая консервация промышленных загрязнителей. М.: Наука, 2005. 190 с.
- Лайдинен Г. Ф., Таланова В. В., Титов А. Ф., К азнина Н. М. Влияние свинца на рост и развитие Setaria viridis (L.) Beauv.//Растительные ресурсы. 2004. Т. 40. Вып. 3. С. 53-59.
- Лукина Н. В., Никонов В. В. Биогеохимические циклы в лесах Севера в условиях аэротехногенного загрязнения. Апатиты: Изд-во КНЦ РАН, 1996. Ч. 1-2. С. 36-47.
- Лукина Н. В., Никонов В. В. Питательный режим лесов северной тайги: Природные и техногенные аспекты. Апатиты: Изд-во КНЦ РАН, 1998. 316 с.
- Лянгузова И. В., Чертов О. Г. Химический состав растений при атмосферном и почвенном загрязнении//Лесные экосистемы и атмосферное загрязнение. Л.: Наука, 1990. С. 75-86.
- Мазная Е. А. Сравнительная характеристика ценопопуляций Vaccinium myrtillus L. в условиях длительного атмосферного загрязнения (Кольский полуостров)//Растительные ресурсы. 2003. Т. 39. Вып. 4. С. 36-47.
- Мазная Е. А., Лянгузова И. В. Параметры ценопопуляций и накопление тяжелых металлов Vaccinium myrtillus L. и V. vitis-idaea L. (Ericaceae) при разном уровне техногенной нагрузки//Растительные ресурсы. 2006. Вып. 1. С. 16-27.
- Неверова О. А., Цандекова О. Л. Фотосинтетическая способность древесных растений как индикатор суммарного загрязнения атмосферного воздуха городской среды//Сибирский экологический журнал. 2010. Вып. 2. С. 193-196.
- Николаевский В. С. Биологические основы газоустойчивости растений. Новосибирск: Наука, 1979. 280 с.
- Отчет о НИР ИППЭС КНЦ РАН. Восстановление территорий, нарушенных антропогенным воздействием в окрестностях г. Мончегорска. Апатиты: Изд-во КНЦ РАН, 2003.
- Сергейчик С. А. Устойчивость древесных растений к техногенной среде. Минск: Наука и техника, 1994. 280 с.
- Серегин И. В., Кожевникова А. Д. Физиологическая роль никеля и его токсическое действие на высшие растения//Физиология растений. 2006. Т. 53. С. 285-308.
- Сухарева Т. А. Многолетняя динамика содержания тяжелых металлов в листьях доминирующих растений бореальных лесов в условиях атмосферного загрязнения//Экологические проблемы северных регионов и пути их решения. Апатиты: Изд-во КНЦ РАН, 2008. С. 156-160.
- Таланова В. В., Титов А. Ф., Боева Н. П. Влияние свинца и кадмия на проростки ячменя//Физиология и биохимия культурных растений. 2001. Т. 33. № 1. С. 33-34.
- Федорков А. Л. Адаптация хвойных к стрессовым условиям Крайнего Севера. Екатеринбург: Изд-во УрО РАН, 1999. 97 с.
- Федорков А. Л. Изменчивость признаков анатомического строения хвои сосны и ее устойчивость к техногенному и климатическому стрессу//Экология. 2002. № 1. С. 70-72.
- Фролов А. К. Окружающая среда крупного города и жизнь растений в нем. СПб.: Наука, 1998. 328 с.
- Черненькова Т. В. Особенности прорастания семян сосны и ели при разной загрязненности почв тяжелыми металлами//Влияние промышленных предприятий на окружающую среду. М.: Наука, 1987. С. 168-182.
- Яковлев А. П. Мониторинг роста и развития Vaccinium myrtillus L. в окрестностях завода по термической обработке твердых бытовых отходов г. Мурманска//Естественнонаучные проблемы Арктического региона. Мурманск, 2007.
- Ярмишко В. Т. Сосна обыкновенная и ее сообщества в условиях атмосферного загрязнения на Европейском Севере: Автореф. дис. … д-ра биол. наук. СПб., 1994.
- Kosobrukhov A., Knyazeva I., Mudrik V. Plantago major plants responses to increase content of lead in soil: Growth and photosynthesis//Plant growth Regul. 2004. Vol. 42. P. 145-151.
- Kozlov M. V., Zve reva E. L. Reproduction of mountain birch along a strong pollution gradient near Monchegorsk, Northwestern Russia//Environmental Pollution. 2004. Vol. 132. P. 443-151.
- Sandalio L. M., Dalurzo H. C., Gomes M., Romero -Puertas M. C., del Rio L. A. Cadmium-induced changes in the growth and oxidative metabolism of pea plants//J. Exp. Bot. 2001. Vol. 52. № 364. P. 2115-2126.
- Vassilev A., Berova M., Zlatev Z. Influence of Cd2+ on growth, chlorophyll content, and water relations in young barley plants//Biol. Plant. 1998. Vol. 41. № 4. P. 601-606.
