Влияние техногенного засоления на некоторые биохимические показатели растений, произрастающих в зоне солеотвалов

Автор: Кусакина Маргарита Григорьевна, Еремченко Ольга Зиновьевна, Четина Оксана Александровна
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Ботаника
Статья в выпуске: 1, 2013 года.
Бесплатный доступ
Представлены результаты исследований содержания сахаров, водорастворимых белков и пролина у некоторых видов растений, произрастающих в условиях техногенного засоления. Выявлены биохимические особенности, обеспечивающие их устойчивость к действию техногенных солей.
Техногенное засоление, адаптация, метаболизм, сахара, водорастворимые белки, пролин
Короткий адрес: https://sciup.org/147204614
IDR: 147204614 | УДК: 581.1.632.1
Influence of technogenic salinization on some biochemical parameters of plants grown in salt dumping area
Results of researches of the content of sugars, water-soluble proteins and proline at some species of the plants growing in the conditions of technogenic salinization are presented. The biochemical features providing their stability to effect of technogenic salts are revealed.
Текст научной статьи Влияние техногенного засоления на некоторые биохимические показатели растений, произрастающих в зоне солеотвалов
В результате антропогенной деятельности наблюдается значительное увеличение техногенной нагрузки на окружающую среду. На территории Верхнекамского месторождения солей производится добыча сырья. Спецификой калийного производства является накопление значительного количества отходов, представленных в основном хлоридами натрия, калия и магния. Галитовые отходы производства являются источником загрязнения окружающей среды. На техногенно засоленных почвах происходит смена растительных сообществ на более устойчивые, что приводит к трансформации зональных ландшафтов в целом [Еремченко, Четина, 2009].
В адаптации растений к различным природным и антропогенным факторам значительная роль принадлежит углеводному обмену. Усиление гидролиза полимерных форм углеводов, в том числе низкомолекулярных олигосахаридов, является неспецифической ответной реакцией на неблагоприятные воздействия различной природы – холодовой, тепловой, осмотический, солевой стрессы и др. [Колупаев, Трунова, 1994; Франко, Мело, 2000].
Аккумуляция растворимых углеводов, являющихся осмотическими веществами, обеспечивает внутреннюю регуляцию водного потенциала и способствует увеличению поглощения воды корнями растений, что является важным в условиях засоления [Кафи, Стюарт, Борланд, 2003; Веселов, Маркова, Кудоярова, 2007]. Сахара оказывают антиде-натурационное действие на белково-липидный комплекс мембран. Они также являются перехват- чиками активных форм кислорода (АФК) и тормозят процессы свободнорадикального окисления биологических молекул при развитии окислительного стресса, индуцированного действием неблагоприятных факторов среды [Mitter, 2002; Дерябин и др., 2007; Карпец, Колупаев, 2009].
Высказывается мнение, что помимо протекторного и антиоксидантного действия, некоторые углеводы могут выполнять сигнальную роль [Rolland, Sheen, 2005].
Формирование устойчивости связано с активностью белоксинтезирующей системы. Белки являются главной мишенью действия экстремальных факторов, и в это же время они служат инструментом репарации и защиты от повреждений [Чудинова, Суворов, Механошина, 2001].
При засолении может изменяться как количественное содержание белков, так и соотношение их компонентного состава. При этом наиболее подвержены изменениям водорастворимые белки, которым принадлежит важная защитная роль в клетках растений в условиях различных стрессов.
К низкомолекулярным метаболитам, накапливающимся в растениях и способствующим их выживанию в неблагоприятных условиях среды относится аминокислота пролин. Как отмечают Н.Л. Радюкина и др. [2007], повышение содержания свободного пролина в условиях засоления является результатом NaCl-зависимой экспрессии генов, кодирующих ферменты его биосинтеза: пирролин-5-карбоксилат-синтетазы и пирролин-5-карбоксилатредуктазы. Пролин имеет важное значение для поддержания клеточного метаболизма, проявляя при этом осмо-
регуляторный, стресс-протекторный и сигнальный эффекты, а также участвует в детоксикации супер-оксид-радикала, снижая образование АФК [Кузнецов, Шевякова, 1999; Hare, Cress, Van Staden, 1999].
Целью нашей работы явилось изучение роли некоторых органических соединений в адаптации дикорастущих видов растений к действию техногенного засоления.
Объекты и методы исследования
В качестве объектов исследования были взяты следующие виды растений: вейник наземный ( Calamagrostis epigeios (L.) Roth), бодяк полевой ( Cirsium arvense (L.) Scop.), молокан татарский ( Lactuca tatarica (L.) C.A. Mey.), произрастающие как в зоне устойчивого засоления (на расстоянии 1– 5 м от солеотвалов), так и в зоне неустойчивого засоления (30–50 м от солеотвалов) – контрольный вариант, а также триостренник приморский ( Triglochin maritimum L.), встречающийся только в зоне устойчивого засоления. Пробы для анализов брали в июне 2011 г. в районе солеотвалов г. Соликамска.
Определение количества растворимых сахаров проводили методом Хагедорн-Иенсена [Ермаков и др., 1987]. Содержание водорастворимых белков определяли спектрофотометрическим методом при двух длинах волн – 230 и 260 нм, определение пролина проводили по методу L.S. Bates, R.P. Waldren, G.D. Teare [1973]. Повторность опытов трехкратная. Полученные результаты статистически обработаны. Определен показатель достоверности между контрольными и опытными вариантами при 95% уровне вероятности.
Результаты и их обсуждение
Существенную роль в защите растений от действия неблагоприятных факторов среды и, в частности, засоления, играют такие низкомолекулярные соединения, как растворимые углеводы. Недостаточное содержание этих основных метаболитов может тормозить процессы роста в условиях засоления.
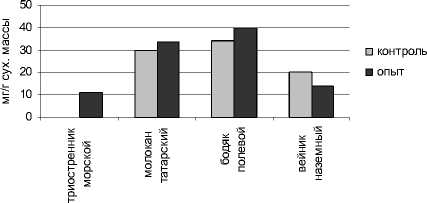
Результаты определения суммарного количества моносахаров (глюкозы и фруктозы) показали, что техногенное засоление оказало неоднозначное влияние на содержание этих соединений в надземной части и корнях исследуемых растений (рис. 1).
У таких видов, как бодяк полевой и вейник наземный в зоне устойчивого техногенного засоления в надземных органах произошло статистически достоверное повышение содержания моносахаров по сравнению с зоной неустойчивого засоления. Накапливающиеся при солевом стрессе углеводы используются для осмотической адаптации [Кафи, Стюарт, Борланд, 2003]. Повышение уровня моносахаров, являющихся одними из низкомо- лекулярных протекторов, можно рассматривать как неспецифический ответ этих видов, способствующий их адаптации к условиям техногенного засоления.
надземные органы
корни
1,6
1,2
0,8
0,4
□ контроль
□ опыт
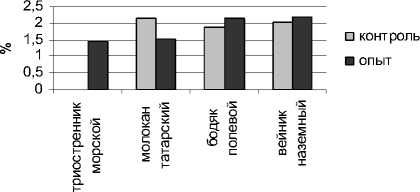
Рис. 1 . Содержание моносахаров в органах исследуемых растений
Накопление углеводов в растениях может быть ответной реакцией на избыток одновалентных ионов натрия и хлора в вакуолях растений при засолении. Органические соединения, накапливающиеся в цитоплазме, функционируют как осмотики, которые, в отличие от одновалентных ионов, не оказывают вредного влияния на ферменты и мембраны [Кафи, Стюарт, Борланд, 2003].
У молокана татарского в зоне повышенной концентрации солей отмечено снижение уровня моносахаров в надземных органах. Понижение уровня этих метаболитов в условиях техногенного засоления, по-видимому, может свидетельствовать о расходовании этих соединений как источника энергии и пластических веществ на биосинтез ин-дуцибельных белков.
Как отмечают А.Н. Дерябин и др. [2007], сахара тормозят процессы свободнорадикального окисления и перехватывают гидроксильные радикалы, образующиеся при воздействии различных абиотических стрессов, и тем самым снижают токсичность ОН-генерирующией системы Фентон.
В связи с этим выявленное снижение содержания сахаров у молокана татарского, вероятно, также может быть связано с расходом этих соединений на обезвреживание активных форм кислорода.
Триостренник приморский, встречающийся только вблизи солеотвалов, характеризовался более низким уровнем растворимых сахаров в над- земной части и большим содержанием в корнях по сравнению с другими исследуемыми видами.
Усиление фактора засоленности вызвало повышение содержания моносахаридов в корнях гликофита вейника наземного и галофита молокана татарского и не оказало существенного влияния на величину данного показателя у бодяка полевого.
Синтез белка является одним из клеточных процессов, крайне чувствительных к осмотическому стрессу, особенно к солевому. Присутствие растворимых белков в цитоплазме способствует осморегуляции [Франко, Мело, 2000].
Исследование содержания водорастворимых белков в органах растений показало, что высокий уровень техногенного засоления неоднозначно повлиял на содержание водорастворимых белков в листьях растений. У такого вида, как вейник наземный произошло снижение содержания этих белков по сравнению с контролем – зоной неустойчивого засоления. Очевидно, это связано с тем, что в условиях действия техногенных солей процессы распада белка преобладали над их синтезом. Торможение синтеза белка может быть связано как с лимитированием этого процесса со стороны обеспеченности энергией и активированными аминокислотами, так и со снижением функциональной активности нуклеиновых кислот [Удовенко, 1977] (рис. 2).
надземные органы
корни
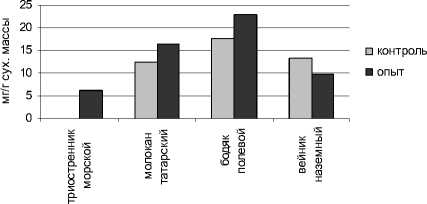
Рис. 2 . Содержание водорастворимых белков в органах исследуемых растений
Под влиянием устойчивого засоления произошло увеличение количества водорастворимых белков в листьях молокана татарского и бодяка полевого, по сравнению с зоной неустойчивого засоления. Возрастание количества альбуминов имеет важное адаптивное значение. Они повышают устойчивость протоплазмы, кроме того, благодаря гидрофильным свойствам этих белков, увеличивается водоудерживающая способность клетки, что так необходимо при засолении, когда уменьшается степень оводненности тканей. Водорастворимые белки, аминокислоты, органические кислоты могут связывать засоляющие ионы, благодаря чему они переходят в метаболически неактивное состояние [Строгонов, 1973].
Водорастворимые белки, составляя основу ферментативного аппарата клетки, играют особую роль в адаптации метаболизма к засоляющим ионам путем изменений в изоферментных спектрах.
Накопление водорастворимых белков при повышенной концентрации техногенных солей у молокана татарского и бодяка полевого, очевидно, может рассматриваться как часть протекторной системы у этих растений. Таким образом, увеличивая количество водорастворимых белков, клетки растений предохраняют коллоиды протоплазмы от токсического влияния ионов солей.
Что касается природных галофитов, то триостренник морской, найденный только в зоне устойчивого засоления, характеризовался низким содержанием водорастворимых белков. Очевидно, приспособление этого вида к действию техногенных солей осуществляется за счет других механизмов, в частности, накопления засоляющих ионов в цитоплазме и такой аминокислоты, как пролин.
Одним из защитных механизмов, который поддерживает водный статус клеток в условиях засоления, является аккумуляция низкомолекулярных органических соединений, таких, как пролин. Свободный пролин и пролин в составе белковой молекулы – обязательные компоненты растительной клетки.
Аккумуляция пролина направлена на поддержание в клетках водного гомеостаза, а также на протекторный эффект. В основе протекторного действия пролина лежат гидрофильные и гидрофобные взаимодействия с водной фазой и макромолекулами цитоплазмы, мембран и клеточных органелл, стабилизирующие их конформацию [Кре-славский и др., 2012].
Определение содержания свободного пролина показало, что в условиях устойчивого техногенного засоления произошло повышение величины данного показателя в органах всех исследуемых растений (рис. 3).
Более высоким содержанием пролина в надземных органах характеризовался галофит триостренник морской. В адаптации другого исследуемого галофита – молокана татарского, пролин также имеет существенное значение. Накопление пролина может происходить в результате ингибирования его распада, усиления синтеза, уменьшенного использования для синтеза белка, а также гидролиза белков [Сунн, Лен, Тян, 2005].
При накоплении пролина в условиях осмотического стресса, окисление его избытка подавлено и сохраняется высокий уровень свободного пролина. Напротив, при снятии водного или солевого стресса окисление пролина активируется. Это окисление рассматривается как регуляторный механизм, в процессе которого избыток пролина используется как энергетический субстрат, а также субстрат для образования глутамата и простых сахаров. Последние активно используются в синтезе крахмала, своеобразного депо сахаров, которые в условиях солевого стресса, наряду с пролином, могут выполнять осморегуляторные функции. Таким образом, в условиях засоления пролин выступает еще и в качестве энергетического субстрата [Шевякова и др., 1994; Франко, Мело, 2000; Dajic, 2006].
надземные органы
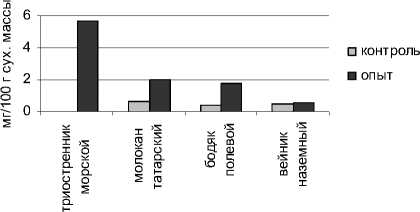
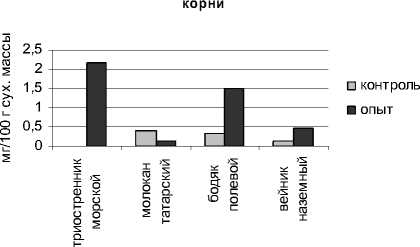
Рис. 3 . Содержание пролина в органах исследуемых растений
Нами установлена взаимосвязь между содержанием пролина и сахаров. Триостренник приморский и бодяк полевой, накапливая значительное количество пролина, характеризовались также довольно высоким уровнем моносахаров. Накопление низкомолекулярных протекторов происходит как за счет повышения активности ферментов, которые обеспечивают их синтез, так и в результате активации катаболического потока [Карпец, Колупаев, 2009].
Проведенные нами исследования показали, что изученные растения характеризовались дифференцированным накоплением свободного пролина в органах. В листьях уровень пролина у всех видов был выше, чем в корнях.
На основании проведенных экспериментов нами установлена взаимосвязь между аккумуляцией засоляющих ионов хлора и накоплением пролина. Триостренник приморский и бодяк полевой, характеризующиеся высоким содержанием в органах ионов свободного хлора, отличались повышенным уровнем пролина.
Выводы
-
1. Повышение уровня моносахаров у бодяка полевого и вейника наземного в условиях устойчивого техногенного засоления имеет важное адаптивное значение, свидетельствуя о наличии у растений энергетических и пластических ресурсов.
-
2. Одним из показателей перестройки обмена веществ в условиях устойчивого засоления явилось увеличение содержания пролина у всех исследуемых растений, что подтверждает его важную роль в преодолении растениями стрессовой ситуации.
-
3. В основе высокой солеустойчивости галофита триостренника приморского лежит его способность к аккумуляции засоляющих ионов и накоплению пролина, а молокана татарского – в накоплении водорастворимых белков.
Список литературы Влияние техногенного засоления на некоторые биохимические показатели растений, произрастающих в зоне солеотвалов
- Веселое Д.С., Маркова И.В., Кудоярова Г.Р. Реакция растений на засоление и формирование устойчивости//Успехи совр. биол. 2007. Т. 127, вып. 5. С. 482-493.
- Дерябин А.Н. и др. Влияние сахаров на развитие окислительного стресса, вызванного гипотермией (на примере растений картофеля, экспрессирующих ген инвертазы дрожжей//Физиол. раст. 2007. Т. 54, № 1. С. 39-46.
- Еремченко О.З., Четина О.А. Адаптация растений к техногенному засолению в таежно-лесной зоне Прикамья//Экология в высшей школе: синтез науки и образования: материалы Всерос. науч.-практ. конф. Челябинск, 2009. Ч. 1. С. 28-32.
- Ермаков А.И. и др. Методы биохимического исследования растений. Л.: Агропромиздат, 1987. 430 с.
- Карпец Ю.В., Колупаее Ю.Е. Ответ растения на гипертермию: молекулярно-клеточные процессы//Вестник Харьков. нац. агр. ун-та. Сер. биол. 2009. Вып. 1. С. 19-39.
- Кафи М.С., Стюарт В.С., Борланд А.М. Содержание углеводов и пролина в листьях, корнях и апексах сортов пшеницы, устойчивых и чувствительных к засолению//Физиол. раст. 2003. Т. 50, № 20. С. 174-182.
- Колупаее Е.В., Трунова Т.И. Активность инвертазы и содержание углеводов в колеоптилях пшеницы при гипертермическом и солевом стрессах//Физиол. раст. 1994. Т. 41, № 4. С. 552-557.
- Креславский В.Д. и др. Сигнальная роль активных форм кислорода при стрессе у растений//Физиол. раст. 2012. Т. 59, № 2. С. 163-178.
- Кузнецов Вл.В., Шевякова Н.И. Пролин при стрессе: биологическая роль, метаболизм, регуляция//Физиол. раст. 1999. Т. 46, вып. 2. С. 321-336.
- Радюкина Н.Л. и др. Сравнительный анализ функционирования защитных систем у представителей галофитной и гликофитной флоры в условиях прогрессирующего засоления//Физиол. раст. 2007. Т. 54, № 6. С. 902-912.
- Строгонов Б.П. Метаболизм растений в условиях засоления М.: Наука, 1973. 380 с.
- Сун С.К., Лен Е.Б.,Э Тян К.Р. Метаболизм пролина и перекрестная устойчивость к засолению и температурному стрессу у прорастающих семян пшеницы//Физиол. раст. 2005. Т. 52, вып. 6. С. 897-904.
- Удовенко Г.В. Солеустойчивость культурных растений. Л.: Колос, 1977. 315 с.
- Франко О.Л., Мело Ф.Р. Осмопротекторы: ответ растений на осмотический стресс//Физиол. раст. 2000. Т. 47, вып. 1. С. 152-159.
- Чудинова Л.А., Суворов В.И., Механошина Л.И. Влияние гипертермии на устойчивость кукурузы к последующему засолению//Вестник Пермского университета. Серия Биология. 2001. Вып. 4. С. 85-90.
- Шевякова Н.И. и др. Стрессорный ответ клеток Nicotiana sylvestris L. на засоление и высокую температуру. 1. Аккумуляция пролина, полиаминов, бетаинов и сахаров//Физиол. раст. 1994. Т. 41, № 4. С. 558-565.
- Bates L.S., Waldren R.P., Teare G.D. Rapid determination of free proline for water stress//Plant Soil. 1973. Vol. 39. P. 205-207.
- Dajic Z. Salt stress//Phisiology and Molecular Biology of Stress Tolerance in plants/eds Madhava Rao K.V., Raghavendra A.S., Janardhan Reddy K. Dordrecht: Kluwer, 2006. P. 41-101.
- Hare P.D., Cress W.A., Van Staden J. Dissecting the roles of osmolyte accumulation during stress//Plant, Cell and Environ. 1998. № 6. P. 535-553.
- Mitter R. Oxidative stress, antioxidants and stress tolerance//Trends Plant Sci. 2002. Vol. 7. P. 405410.
- Rolland F., Sheen J. Sugar sensing and signaling networks in plant//Biochemical Society Transaction. 2005. Vol. 33. P. 269-271.
