Влияние температурной предобработки на конститутивную и индуцибельную устойчивость проростков ржи к хлоридному засолению

Автор: Чудинова Л.А., Юсупов Д.Р.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Экология
Статья в выпуске: 2, 2021 года.
Бесплатный доступ
Изучали интенсивность роста проростков ржи, а также динамику содержания в побегах растворимых белков и пролина в процессе их адаптации к резкому (300 мМ NaCl одноразово, время экспозиции 9 сут.) и постепенному (100 мМ NaCl, затем по 100 мМ NaCl через 2 сут. до конечной концентрации 400 мМ) засолению хлоридом натрия в условиях наличия или отсутствия теплового закаливания (+40°С, 3 ч.). Установленная динамика содержания пролина и растворимых белков в побегах позволяет предполагать, что формирование устойчивости к засолению определяется высоким конститутивным уровнем пролина, а также стресс-индуцибельным синтезом пролина и водорастворимых белков. Тепловая предобработка проростков в большей степени стимулировала их конститутивную устойчивость. Обнаруженные метаболические изменения, очевидно, имеют отношение к одному из возможных механизмов протекторного действия теплового закаливания к последующему засолению.
Проростки ржи, засоление хлоридом натрия, тепловое закаливание, растворимые белки, пролин
Короткий адрес: https://sciup.org/147235450
IDR: 147235450 | УДК: 581.1:632.122.1 | DOI: 10.17072/1994-9952-2021-2-134-140
Influence of temperature processing on the constitutive and inducible resistance of rye seedlings to chloride salinization
We studied the growth rate of rye seedlings, as well as the dynamics of the content of soluble proteins and proline in the shoots during their adaptation to sharp (300 mM NaCl once, exposure time 9 days) and gradual (100 mM NaCl, then 100 mM NaCl after 2 days to the final concentration of 400 mM) salinity with sodium chloride in the presence or absence of thermal hardening (+40°C, 3 h). The established dynamics of the content of proline and soluble proteins in the shoots suggests that the formation of resistance to salinity is determined by the high constitutive level of proline, as well as the stress-inducible synthesis of proline and water-soluble proteins. Thermal pretreatment of the seedlings stimulated their constitutive stability to a greater extent. The detected metabolic changes are obviously related to one of the possible mechanisms of the protective effect of thermal hardening on subsequent salinization.
Текст научной статьи Влияние температурной предобработки на конститутивную и индуцибельную устойчивость проростков ржи к хлоридному засолению
Актуальность проблемы солеустойчивости растений определяется тем, что засоленные почвы снижают как продуктивность, так и биоразнообразие фитоценозов [Zhu J.-K., 2001; Чиркова, 2002]. В связи с этим вопрос о солеустойчивости растений, а также механизмы адаптации на молекулярном и молекулярно-генетическом уровне в настоящее время активно изучаются [Колупаев, Карпец, 2010].
Известно, что солеустойчивость растений определяется функционированием конститутивных и индуцибельных систем. Оба компонента устойчивости генетически детерминированы и направлены на сохранение жизнеспособности организма в экстремальных условиях [Радюкина и др., 2007].
Принципиальное различие этих компонентов заключается в том, что конститутивные механизмы постоянно находятся в функционально активном состоянии, а индуцибельные – в норме отсутствуют и появляются лишь в ответ на действие повреждающего фактора. О конститутивной устойчивости растений свидетельствует их способность выживать в условиях шокового действия фактора высокой интенсивности, тогда как об индуцибель-ной устойчивости говорит способность постепенно адаптироваться к длительному действию стрессора возрастающей интенсивности [Радюкина и др., 2007].
Механизмы формирования стресс-индуцибель-ных защитных систем в условиях солевого стресса на сегодняшний день остаются недостаточно изученными. В этом плане особый интерес представляет кросс-адаптация (перекрестная адаптации), т.е. явление, когда действие одного неблагоприятного фактора среды приводит к повышению устойчивости растений к другим факторам. Оно известно довольно давно, но до сих пор его молекулярнобиохимические механизмы остаются до конца не выясненными.
В ряде исследований отмечается, что кратковременный тепловой шок оказывает протекторный эффект к последующим тепловому и холодовому воздействиям, а также водному дефициту [Титов, Таланова, 2009]. В отношении комбинированного действия на растения закаливающих температур и засоления данных в литературе недостаточно.
Имеются основания считать, что в процессе перекрестной адаптации участвуют разнообразные низкомолекулярные соединения (пролин, органические кислоты, растворимые сахара, полиамины и др.), большинство из которых проявляет полифункциональность и выполняет протекторные функции [Карпец, Колупаев, 2009].
В связи с этим цель данной работы – исследование способности проростков ржи адаптироваться к засолению за счет функционирования конститутивных и стресс-индуцируемых защитных механизмов, а также влияние на эти процессы теплового закаливания.
Были поставлены следующие задачи: оценить влияние засоления NaCl (резкого и постепенного), а также предварительного теплового закаливания на ростовые процессы проростков ржи; изучить динамику водорастворимых белков и пролина в проростках ржи в контрольных и опытных вариантах.
Объект и методы исследований
Семена ржи посевной ( Secale cereale L . ) стерилизовали 20 мин. в 1‒4%-ном растворе перманганата калия, затем выращивали в условиях водной культуры в пластиковых контейнерах (субстрат – керамзит с добавлением агроперлита) при комнатной температуре (20–23ºС) в условиях естественного освещения. Восьмидневные проростки подвергали следующим стрессовым воздействия:
-
1 серия – а) резкое засоление: 300 мМ NaCl одноразово, время экспозиции 9 сут. и б) постепенное засоление: 100 мМ NaCl, затем по 100 мМ NaCl через 2 сут. до конечной концентрации 400 мМ;
-
2 серия – температурная предобработка +40ºС в течение 3 ч. до засоления (резкого или постепенного).
Физиологические и биохимические показатели анализировали после 2-; 4-; 6- и 9-суточной экспозиции проростков в засоленной среде в первой половине дня. Для характеристики интенсивности роста определяли длину побегов (30-кратная повторность). Содержание суммы водорастворимых белков определяли по методу с амидо-черным 10 В в сыром материале [Большой …, 2012], количество пролина – методом L.S. [Bates, Waldren, Teatre, 1973] в растительном материале, фиксированном паром в течение 5 мин. и досушенном при +70ºС.
Повторность биохимических анализов – трехкратная. Результаты обрабатывали статистически с использованием однофакторного дисперсионного анализа в программе Stadia. Определяли достоверность различий между некоторыми вариантами по наименьшей существенной разнице (d > НСР 0.05 ).
Результаты и обсуждение
При оценке устойчивости сортов и видов растений к различным экологическим факторам среды наиболее широко используются методы, в основе которых лежит реакция ростовых процессов, так как в большинстве случаев при стрессах различной природы наблюдается угнетение роста. По отклонению величины измеряемого показателя от контроля можно судить о силе воздействия фактора на растение и о степени его устойчивости [Титов, Таланова, 2009].
Как показали наши исследования (рис. 1, А), в условиях засолениях наблюдалась общая тенденция уменьшения линейных размеров проростков ржи. При этом амплитуда зависела от характера засоления (резкого или постепенного). Так, при применении шокового засоления длина побега уменьшалась на 21% от контроля, а в условиях стресса, вызванного постепенным засолением – на 14% от контроля.
Отсюда можно предположить, что у проростков ржи при солевом стрессе, вызванном NaCl, в большей степени подавляется конститутивная устойчивость. Это согласуется с выводом некоторых авторов о том, что конститутивные протекторные системы лежат в основе солеустойчивости галофитов, тогда как устойчивость гликофитов основана главным образом на функционировании стресс-индуцируемых механизмов [Кузнецов, 2001].
Тепловое закаливание снижало рост-ингибирующий эффект засоления в отношении длины побегов, причем в большей степени в условиях резкого засоления (рис. 1, Б). По-видимому, тепловое закаливание повышает как конститутивную, так и индуцибельную солеустойчивость проростков ржи, причем более существенно – конститутивную.
Это согласуется с выводом ряда авторов о том, что адаптивные процессы, индуцированные в растении при первом воздействии, вызывают увеличение устойчивости ко второму, что обусловлено развитием преимущественно неспецифических ре- акций (кросс-адаптация) [Титов, Таланова, 2009;
Чудинова, Суворов, 2011].
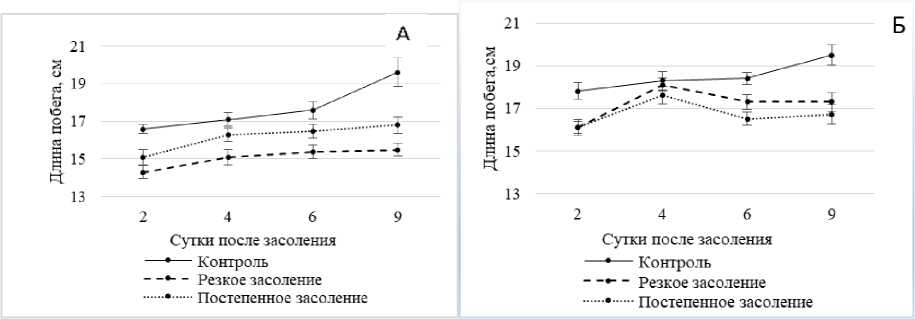
Рис. 1 . Изменение интенсивности роста побегов ржи в условиях хлоридного засоления без температурной предобработки (А) и после температурной предобработки (Б).
Здесь и далее: резкое засоление – 300 мМ NaCl одноразово; постепенное засоление – 100 мМ NaCl,затем по 100 мМ NaCl через 2 сут. до конечной концентрации 400 мМ; температурная предобработка +40ºС в течение 3 ч. до засоления
Таким образом, наши данные и анализ литературы однозначно свидетельствуют о том, что у растений, наряду со специализированными адаптивными механизмами, существуют и общие механизмы устойчивости к засолению и неблагоприятным температурам. Это позволяет растению избегать огромных энергетических затрат, связанных с необходимостью формирования специализиро- ванных механизмов адаптации в ответ на любое отклонение условий обитания организма от нормальных. Ряд исследователей устойчивость к засолению и высокой температуре связывают с накоплением одних и тех же стрессовых белков [Ruy et al., 1995], этилена, пролина и путресцина [Кузнецов, 2001].
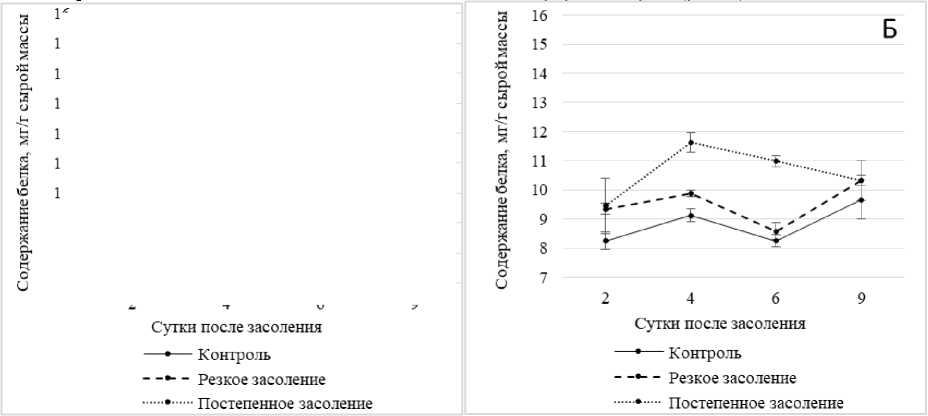
Рис. 2. Динамика содержания белка в побегах ржи в условиях хлоридного засоления без температурной предобработки (А) и после температурной предобработки (Б)
Полученные нами результаты показали, что у проростков ржи в ответ на действие резкого и постепенного хлоридного засоления уже через 2 сут.
Маркером изменения метаболизма клеток в целом часто рассматривают белковый синтез, поскольку он является центральным звеном метаболизма и очень чувствителен к внешним условиям роста растений [Шевякова и др., 2013]. Об этом, в частности, говорит тот факт, что подавление белкового синтеза с помощью тех или иных ингибиторов транскрипции или трансляции препятствует росту устойчивости, а стимуляция работы белок-синтезирующего аппарата, наоборот, положительно сказывается на устойчивости [Карпец, Колупаев, 2009].
В связи с этим нами изучалось влияние засоления на содержание водорастворимых белков в побегах проростках ржи (рис. 2).
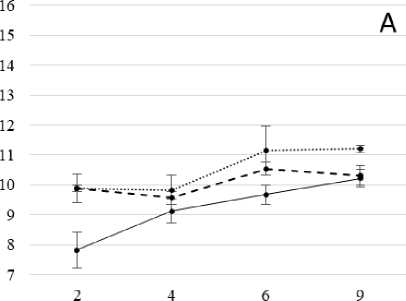
наблюдался значительный рост содержания белка (на 27% выше контрольного), который сохранялся при увеличении продолжительности солевого стресса до 9 сут. (рис. 2, А). Причем, в большей степени белок накапливался в условиях постепенного засоления, что говорит о преобладании стресс-индуцированного синтеза белка.
Под влиянием тепловой предобработки (рис. 2, Б) был отмечен более сильный рост содержания белка в условиях стресса, вызванного постепенным засолением по сравнению с вариантами без теплового закаливания. Это, очевидно, имеет отношение к одному из возможных механизмов протекторного действия теплового закаливания. В вариантах с резким засолением стимулирующее влияние тепловой предобработки на синтез белков было существенно слабее. Эффекты теплового закаливания растений, как правило, сопровождаются повышением термостабильности белков (в частности, ферментов) и, как результат, определенных функций клеток [Титов, Таланова, 2009].
На основании полученных результатов можно сделать предположение, что формирование конститутивной и индуцибельной устойчивости проростков ржи к хлоридному засолению связано с активированием синтеза водорастворимых белков, причем в большей степени за счет стресс-
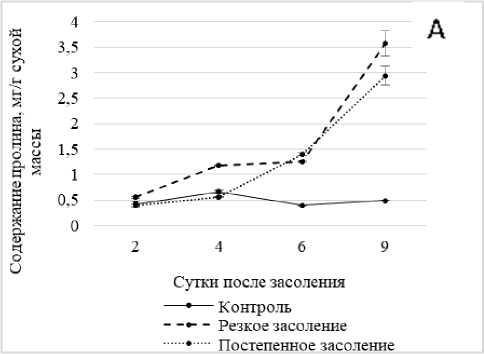
Рис. 3 . Динамика содержания пролина в побегах ржи в условиях хлоридного засоления без температурной предобработки (А) и после температурной предобработки (Б)
Полученные результаты показали значительный рост концентрации пролина в условиях засоления по мере увеличения времени экспозиции проростков ржи в солевом растворе. Так, при 9-суточном солевом шоке содержание пролина превысило контроль более чем в 7, а в условиях постепенного засоления – в 6 раз (рис. 3, А).
Предварительное тепловое закаливание изменило динамику протекания этого процесса в том плане, что рост концентрации пролина при увеличении времени экспозиции на солевом растворе был постепенным и менее значительным – не более чем в 2.6 раза относительно контроля (рис. 3, Б).
Резюмируя эту часть наших исследований, можно предположить, что у данного вида имеет место как индуцибельный, так и конститутивный синтез проли- индуцированного синтеза, который носит адаптивный характер.
По мнению многих авторов, аминокислота пролин, содержание которой в растениях увеличивается в несколько раз в условиях засухи, засоления, действия высоких и низких температур и других повреждающих факторов, может служить индикатором стресса [Hare, Cress, Van Staden, 2000; Кузнецов, 2001; Чиркова, 2002]. Среди стресс-протекторных функций пролина указываются функции химического шаперона, антиоксиданта, регулятора экспрессии стресс-регулируемых генов, источника углерода, азота и восстановительных эквивалентов, регулятора внутриклеточного рН-стата и т.д. [Колупаев, Вайнер, Ястреб, 2014; Ефимова и др., 2018].
Однако, несмотря на многолетнее изучение физиологических функций «стрессового» пролина, многие вопросы, связанные с его участием в адаптации растений к неблагоприятным условиям, остаются невыясненными.
В связи с этим в нашем эксперименте было определено количество пролина в побегах проростков ржи (рис. 3).
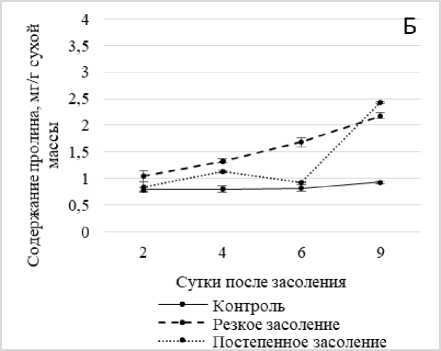
на с преобладанием последнего. Сходная динамика пролина была отмечена в литературе [Kant еt al, 2006; Ахмад, Джалил, Шарма, 2010; Чудинова, Суворов, 2011; Шевякова и др., 2013]. Делается предположение, что регуляция ферментов синтеза пролина осуществляется не только на транскрипционном, но и на пост-транскрипционном уровнях [Радюкина и др., 2007].
В отношении динамики пролина при тепловом закаливании мнения авторов расходятся. В одних работах указывается, что кратковременный тепловой шок стимулирует способность растений аккумулировать пролин в условиях последующих почвенной засухи или засоления. В других говорится, что тепловой шок сам по себе не индуцирует синтез пролина, а лишь делает растения компетент- ными ответить на действие последующих стрессоров [Кузнецов, 2001].
В нашем эксперименте в вариантах с предварительным тепловым закаливанием, в целом, уровень пролина оставался выше контрольного. Однако рост его концентрации при увеличении времени экспозиции на солевом растворе был постепенным и менее значительным. Причиной этого может быть создание дефицита глутамата, необходимого для биосинтеза пролина [Радюкина и др., 2007], поскольку происходит интенсивный расход этой аминокислоты на синтез стрессовых белков, который стимулируется тепловым закаливанием. Можно предположить также, что тепловое закаливание изменяет вклад различных биологических функций пролина, а также других низкомолекулярных протекторов в формирование толерантности растений в зависимости от стадии адаптационного процесса, так называемое «разделение труда» между разными низкомолекулярными протекторами. Это согласуется с данными литературы [Чудинова, Суворов, 2011; Колупаев, Вайнер, Ястреб, 2014].
Заключение
Резюмируя полученные нами данные, следует отметить:
-
- солевой шок оказывает достоверно более сильное ингибирующее влияние на интенсивность роста проростков ржи, чем постепенное засоление. Отсюда можно предположить, что у данного вида гликофитов наиболее активны стресс-
- индуцибельные механизмы устойчивости, а не конститутивные;
-
- тепловая предобработка (+40ºС в течение 3 ч.) проростков ржи изменяет их ростовую реакцию на последующее хлоридное засоление, способствуя смягчению неблагоприятного действия как резкого, так и постепенного засоления. При этом в большей степени стимулируется конститутивная устойчивость;
-
- динамика пролина и растворимого белка в побегах проростков ржи в норме и при солевом стрессе позволяет предполагать, что формирование устойчивости к засолению определяется высоким конститутивным уровнем пролина, а также стресс-индуцибельным синтезом пролина и водорастворимых белков. Обнаруженные изменения в метаболизме пролина и растворимых белков, очевидно, имеют отношение к одному из возможных механизмов протекторного действия теплового закаливания к последующему засолению.
Список литературы Влияние температурной предобработки на конститутивную и индуцибельную устойчивость проростков ржи к хлоридному засолению
- Ахмад П., Джалил К.А., Шарма С. Влияние солевого стресса на систему антиоксидантной защиты, перекисное окисление липидов, ферменты метаболизма пролина и биохимическую активность у двух генотипов шелковицы // Физиология растений. 2010. Т. 57, № 4. С. 547-555.
- Большой практикум «Биохимия»: лабораторные работы / сост. М.Г. Кусакина, В.И. Суворов, Л.А. Чудинова. Пермь, 2012. С. 26-35.
- Ефимова М.В. и др. Физиологические механизмы устойчивости растений Solanum tuberosum L. к хлоридному засолению // Физиология растений. 2018. Т. 65, № 3. С. 196-206.
- Карпец Ю.В., Колупаев Ю.Е. Ответ растений на гипертермию: молекулярно-клеточные аспекты // Вестник Харьковского национального аграрного университета. Серия: Биология. 2009. Вып. 1 (16). С. 19-38.
- Колупаев Ю.Е., Вайнер А.А., Ястреб Т.О. Пролин: физиологические функции и регуляция содержания в растениях в стрессовых условиях // Вестник Харьковского национального аграрного университета. Серия: Биология. 2014. № 2. С. 6-22.
- Колупаев Ю.Е., Карпец Ю.В. Формирование адаптивных реакций растений на действие абиотических стрессоров. Киев: Основа, 2010. 352 с.
- Кузнецов Вл.В. Общие системы устойчивости и трансдукция стрессорного сигнала при адаптации растений к абиотическим факторам // Вестник Нижегородского университета им. Н.И. Лобачевского. Серия: Биология. 2001. Б/н. С. 64-68.
- Радюкина Н.Л. и др. Изучение индуцибельных и конститутивных механизмов устойчивости к солевому стрессу у гравилата городского // Физиология растений. 2007. Т. 54, № 5. С. 692698.
- Титов А.Ф., Таланова В.В. Устойчивость растений и фитогормоны. Петрозаводск: Карельский научный центр РАН, 2009. 206 с.
- Чиркова Т.В. Физиологические основы устойчивости растений. СПб.: Изд-во СПбГУ, 2002. 244 с.
- Чудинова Л.А., Суворов В.И. Роль некоторых низкомолекулярных соединений в механизме перекрестной адаптации растений // Вестник Пермского университета. Серия: Биология. 2011. Вып. 1. С. 17-20.
- Шевякова Н.И. и др. Регуляция абсцизовой кислотой содержания полиаминов и пролина в растениях фасоли при солевом стрессе // Физиология растений. 2013. Т. 60, № 1. С. 1-13.
- Bates L.S., Waldren R.P., Teatre I.D. Rapid Determination of Free Proline for Water Stress Studies // Plant and Soil. 1973. Vol. 39. P. 205-207.
- Hare P.D., Cress W.A., Van Staden J. Proline synthesis and degradation: a model system for elucidating stress-related signal transduction // Journal of experimental botany. 1999. Vol. 50, № 333. Р. 413-434.
- Kant S. et al. Evidence that differential gene expression between the halophyte Thellungiella hal-ophila and Arabidopsis thaliana is responsible for higher levels of the compatible osmolyte proline and tight control of Na+ uptake in T. halophila // Plant Cell and Environment. 2006. Vol. 29, № 7. P. 1220-1234.
- Ryu S.B. et al. Induction of cold hardiness by salt stress involved synthesis of cold and abscisic acid-responsive proteins in potato (Solanum commer-sonii Dun.) // Plant and cell physiology. 1995. Vol. 36, № 7. P. 1245-1251.
- Zhu J.K. Plant salt tolerance // Trends Plant Sci. 2001. Vol. 6. P. 66-71.
