Влияние температуры на эндемичных и неэндемичных турбеллярий

Автор: Ключевская А.А., Стом Д.И.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Биология и экология
Статья в выпуске: 2 т.8, 2006 года.
Бесплатный доступ
Изучено влияние температурного фактора на эндемичные и неэндемичные виды планарий озера Байкал и окружающих его водоемов (Baikalobia guttata, B. variegata, Armilla livanovi и Phagocata sibirica) в условиях лабораторного эксперимента. Установлено, что термопреферендум и темпе- ратурные оптимумы скоростей передвижения испытанных видов планарий, коррелируют с пока- зателями температур зон их обитания.
Короткий адрес: https://sciup.org/148197837
IDR: 148197837 | УДК: 574.5
Influence of temperature on endemic and Siberian fauna turbellaria
Influence of the temperature factor on endemics and non-endemics planarian (Baikalobia guttata, B. variegata, Armilla livanovi and Phagocata sibirica) of lake Baikal and the reservoirs of its basin (Baikalobia guttata, B. variegata, Armilla livanovi and Phagocata sibirica) was investigated. Experiments have been carried out in the laboratory. Thermopreference and temperature optimum of speeds Turbellaria correlated with the parameters of temperatures of the zones of their inhabitation.
Текст научной статьи Влияние температуры на эндемичных и неэндемичных турбеллярий
Турбеллярии составляют многочисленный компонент большинства биоценозов Байкала. По данным О.А.Тимошкина в Байкале уже сегодня насчитывается около 150 видов планарий [1]. Но экология байкальских тур-беллярий исследована недостаточно, известно лишь, что основная часть червей обитает в литоральной зоне озера. В связи с этим изучали влияние температурного фактора на байкальских и общесибирских видов планарий в условиях лабораторного эксперимента.
Планарий содержали в темноте, при температурах, близких к местам их отлова (около 8-100С) в аквариумах объемом 2-3 литра с байкальской водой. Воду в аквариумах меняли ежедневно. Для опытов отбирали половозрелых (имеющих половое отверстие), неповрежденных планарий. Размеры P. sibirica в среднем составляли 15 мм : 3 мм, Baicalobia guttata 10 мм : 2 мм. Если дополнительно не оговорено, то сходные условия стремились поддерживать и при постановке опытов. В эксперименты брали планарий, адаптированных в течении суток к аквариумным условиям.
Для оценки влияния температуры на скорость передвижения планарий, их помещали в кювету, выложенную фильтровальной бумагой, с водой требуемой температуры (диапазон от 5 до 300С). Движение червей инициировали, освещая их естественным светом 5000 – 10000 люкс, заставляя уходить турбел-лярий в темноту. Часть кюветы закрывали плотной темной тканью, не пропускающей свет. Рядом с передним концом тела плана-рии, на фильтровальной бумаге, карандашом ставили точку, обозначающую начало движения. Длину пройденного пути измеряли линейкой, время его прохождения засекали секундомером. Каждую особь использовали в экспериментах только один раз.
Температурный оптимум, т.е. область предпочитаемых температур находили, применяя термоградиентную установку по общепринятым процедурам [3]. Эксперименты по оценке термопреференции проводили в сделанной из металла установке (длина 2 метра, ширина
25 см, высота бортика 10 см). Необходимого температурного градиента достигали за счет того, что один конец садка подогревали с помощью соответствующего устройства, а противоположный охлаждали, используя холодильный агрегат [4]. Вся площадь садка была разделена на участки, в которых постоянно измеряли температуру. Перепад температур в них составлял 1 – 1,5 0С. В зависимости от необходимости создавали температурный градиент от 0 до 10 °С или от 3 до 25 °С. Только после того, как градиент температур стабилизировался, гидробионтов по 30 особей помещали в садок. Первые определения распределения беспозвоночных по температурным зонам проводили спустя 15 минут от момента их запуска. Последующий подсчет количества объектов в разных зонах осуществляли каждые 15 минут в течение 2–3 часов.
В экспериментах по определению термоустойчивости использовали чашки Петри. Термоустойчивость определяли при +20, +25, +30, +35 0С, помещая планарий в воду, предварительно доведенную до этих значений. Критерием гибели служило полное подавление двигательной активности, отсутствие какой-либо реакции на прикосновение кисточкой и частичное нарушение целостности покровов, проявляющееся во фрагментации тканей.
Представленные в работе данные являются средними из 5-10 биологических и 4-х аналитических повторностях. Для каждой аналитической повторности брали не менее 10 экземпляров червей. Статистическую обработку проводили с использованием пакета программ STATISTIKA 5.0 и MICROSOFT EXCEL. Выводы сделаны при вероятности безошибочного прогноза P ≥ 0,90.
Результаты исследования и их обсуждение
При определении влияния температуры на скорость передвижения изучаемых видов планарий, выявили, что максимальную скорость передвижения речная Ph. sibirica развивала при температуре 5 оС, и она составляла 2,7±0,4 мм/сек (табл. 1). При дальнейшем увеличении температуры у этого вида наблюдали понижение скорости. Самые низкие значения скоростей передвижения получены при температуре 30 оС. B. guttata и B. variegata имели самую минимальную скорость передвижения при температуре 5 оС – 0,7±0,3 мм/ сек и 0,3±0,04 мм/сек соответственно. Повышение температуры сопровождалось постепенным увеличением их скоростей движения и достигало пика при 20 0С у B. variegata, (скорость = 0,9±0,2 мм/сек) и при 25?С у B. guttata (скорость = 1,9±0,3 мм/сек).
Ph. sibirica и B. guttata вплоть до 25 оС показывали фактически 100 % выживаемость в течение 60 минут экспозиции (табл. 2). При повышении температуры до 30 оС двигательная активность планарий постепенно снижалась, турбеллярии начинали выделять белую слизь и на 60-й минуте при высоких температурах практически все особи Ph. sibirica и B. guttata погибали.
Исследование распределения планарий в термоградиентной установке выявило наличие ярко выраженного оптимума температурной преференции. На основании полученных данных построены графики, отражающие распределение Ph.sibirica и B.guttata в термоградиентной установке, которые показаны на рис. Наиболее четким температурным максимумом обладала неэндемичная Ph.sibirica, предпочитавшая зону от 0-5 оС, где концентрировалось до 77 % особей. Ph.sibirica, обитающая в ключе, предпочитала температуры от 0- 2 оС, причем часть планарий оказывалась вмерзшей в лед (приблизительно 56 %). По окончании эксперимента вмерзшие планарии оттаивали без видимых нарушений. Байкальский эндемик B.
Таблица 1. Скорость движения (мм/сек) планарий при различных температурах
|
Температура, 0С |
5 |
10 |
15 |
20 |
25 |
30 |
|
Ph. sibirica |
2,7±0,8 |
- |
1,4±0,3 |
1,4±0,7 |
0,8±0,3 |
0,2±0,03 |
|
B. guttata |
0,7±0,3 |
0,8±0,3 |
1,4±0,7 |
0,8±0,3 |
1,9±0,3 |
- |
|
B. variegata |
0,3±0,04 |
0,5±0,05 |
0,8±0.04 |
0,9±0,2 |
0,3±0,01 |
- |
Таблица 2. Численность Ph. sibirica и B.guttata (в процентах к общему числу особей), выживших в течение экспериментов при температурах 25 оС и 30 оС
|
Время, мин |
Ph. sibirica |
B.guttata |
||
|
25 оС |
30 оС |
25 оС |
30 оС |
|
|
10 |
100 |
100 |
100 |
100 |
|
20 |
100 |
100 |
100 |
100 |
|
30 |
100 |
80±5 |
100 |
76±3 |
|
40 |
100 |
80±5 |
100 |
44±4 |
|
50 |
100 |
48±4 |
100 |
28±4 |
|
60 |
96±1 |
8±7 |
100 |
4±6 |
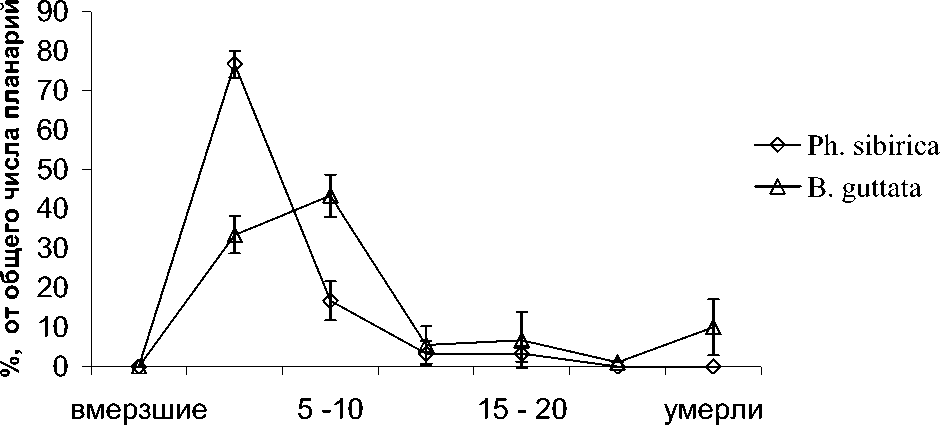
температурные диапазоны, градусы
Рис. Распределение планарий в термоградиентной установке, в % от общего числа guttata выбирал более высокие температуры – 5-10 оС, где сосредотачивалось до 45 % особей. Другой байкальский эндемичный вид, A. livanovi, предпочитал более низкие температуры –2-4 оС – (25%), что можно, предположительно, объяснить тем, что он обитает на больших глубинах (от 3 до 35 метров) [1], чем B. guttata, и там колебания температуры выражены менее незначительно.
Рассмотрение величин термопреференции и оптимума двигательной активности изученных планарий позволило их расположить в следующий ряд в порядке их возрастания Ph. sibirica, < B. variegata, < A. livanovi, < B. guttata. Следовательно, видам турбеллярий с более высокими значениями термопреференции соответствовали и боль- шие значения температурного оптимума скоростей перемещения. Самые низкие величины термопреференции и температурного оптимума скоростей перемещения были установлены у Ph. sibirica. По-видимому, это обусловлено с тем, что этот вид обитает в горных холодных речках и ручьях, где температура очень слабо поднимается, даже летом [5]. Более высокие значения этих величин у B. guttata– можно объяснить, в частности, тем, что данный вид – обитатель верхней литорали Байкала, где летом происходит существенное прогревание воды [6]. Более низкие значения скоростей у B. variegata, возможно, связаны с обитанием на несколько больших глубинах, где волновые явления менее выражены и турбеллярии меньше вынуждены сопротивляться перемещениям воды.
Таким образом, термопреферендум и температурные оптимумы скоростей передвижения испытанных видов планарий, коррелировали с показателями температур зон их обитания.
Авторы считают своим приятным долгом выразить искреннюю признательность д.б.н. О. А. Тимошкину за внимание к работе и ценные советы.
Работа выполнена при финансовой поддержке РФФИ (грант 04-04-48945).
Список литературы Влияние температуры на эндемичных и неэндемичных турбеллярий
- Тимошкин О.А. Аннотированный список фауны озера Байкал и его водосборного бассейна. Новосибирск: Наука, 2001. Т. 1: Озеро Байкал, кн. 1.
- Порфирьева Н.А. Фауна планарий озера Байкал. Казань, 1973.
- Elliot W.R., Mitchell W. Temperature preference responses of some aquatic, caveadapted crustaceans from Central Texas notheastern Mexico//Int. J. Speliol. 5. 2. 1973.
- Timofeyev M. A., Shatilina J. M., Stom D. I. Attitude to temperature factor of some endemic amphipods from Lake Baikal and Holarctic Gammarus lacustris Sars, 1863: A comparative experimental study Arthropoda Selecta10 (2).
- Дыганова Р.Я, Порфирьева Н.А. Планарии азиатской части СССР (Морфология, систематика, распространение). Казань: изд-во Казан. ун-та, 1990.
- Шимараев М.Н. Общая схема взаимодействия обменных процессов в Байкале//Водные ресурсы Байкальского региона: проблемы формирования и использования на рубеже тысячелетий: Матер. науч.-практ. конф. 1998.
