Влияние температуры на скорость роста амилолитических штаммов Geomyces pannorum

Автор: Хижняк С.В., Пучкова Ланкина Е.П., Невзоров В.Н., Олейникова Е.Н., Янова М.А.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Биологические науки
Статья в выпуске: 1, 2018 года.
Бесплатный доступ
Наши предыдущие исследования продемон-стрировали перспективность поиска произво-дителей низкотемпературных амилаз в пе-щерных микробных сообществах. В данной статье представлены результаты исследо-вания влияния температур в диапазоне от 9 до 30 °C на скорость роста восьми амило-литических штаммов Geomyces pannorum, вы-деленных из низкотемпературной карстовой пещеры Караульная-2 (Красноярский край). Бы-ло установлено, что все штаммы являются психопротерантами с максимальной темпе-ратурой роста при 30 °C и оптимальными температурами роста в диапазоне от 18,5 до 25,3 °C. Скорость роста при оптимальной температуре колеблется от 4,2 до 17,5 мкм/ч на одну гифу в зависимости от штамма. Не обнаружено корреляции между оптимальным значением температуры и значением скоро-сти роста при оптимальной температуре. Средняя оптимальная температура для вось-ми штаммов составляет 22,2 °C, средняя скорость роста при оптимальной темпера-туре - 10,2 мкм/ч. В большинстве случаев влияние температуры на скорость роста можно адекватно (с коэффициентами детер-минации R2 в диапазоне от 0,987 до 0,999) описать с помощью модели Ratkowsky et al. (1983). Для некоторых штаммов температур-ные кривые соответствуют модели Ратков- ского только вблизи экстремума. Результаты исследования демонстрируют возможность производства грибной амилазы при темпера-туре примерно на 15 градусов ниже темпера-туры, которая используется сейчас для таких продуцентов, как Aspergillus awamori. Это по-зволяет снизить затраты энергии на произ-водство амилазы и замедлить рост посто-ронней мезофильной микрофлоры в случае микробного загрязнения ферментёра.
Низкотемпературные продуценты амилазы, мицелиальные грибы, температура, скорость роста
Короткий адрес: https://sciup.org/140224312
IDR: 140224312 | УДК: 579.6+
Effect of temperature on the growth rate of amylolytic strains of Geomyces pannorum
Our previous researches demonstrated per-spective of search for low-temperature amylase producers in cave microbial communities. In pre-sent work effect of temperatures in the range of +9 to +30 °C on the growth rate of eight amylolytic strains of Geomyces pannorum isolated from low-temperature karst cave Karaulnaya-2 has been studied. All the strains were found to be psychrotolerant with the maximal growth tempera-ture at +30°C and the optimal growth temperatures in the range of +18.5 to +25.3 °C. Growth rate at the optimal temperature varies from 4.2 to 17.5 mi-crometer per hour per one hyphae depending on the strain. No correlation between optimal tempera-ture value and value of growth rate at optimal tem-perature was found. The average optimum temper-ature for eight strains is +22.2 °C and the average growth rate at the optimal temperature is 10.2 mi-crometer per hour. In most cases effect of temper-ature on growth rate can be adequately described by the Ratkowsky et al. (1983) model with the coef-ficients of determination R2 in the range of 0.987 до 0.999. For some strains the temperature curves fits the Ratkowsky model only near extremum. The results of the research demonstrate possibility to produce fungal amylase at temperature about 15 degrees lower then temperature which is used now for such producers as Aspergillus awamori. This allows to reduce energy costs of amylase produc-ing and to slow down growth of extraneous mesophilic microflora in case of microbial contami-nation of fermenter.
Текст научной статьи Влияние температуры на скорость роста амилолитических штаммов Geomyces pannorum
Введение. Амилазы микробного происхождения широко используются в бродильном, крахмало-паточном, текстильном и бумажном производствах, в производстве глюкозы и органических кислот в качестве биодобавок в моющие средства [4]. Показан положительный эффект от использования микробных амилаз для повышения усвояемости крахмалсодержащих кормов в животноводстве [6, 9]. В последние годы наблюдается рост использования амилаз для производства ферментативно-модифицированного крахмала, используемого в продуктах питания в качестве заменителя жира и улучшителя консистенции [3]. По современным оценкам, микробные амилазы занимают около 25 % мирового рынка ферментов [4]. До недавнего времени в промышленности и сельском хозяйстве применялись главным образом терм о ф ильные амилазы c температурными опти-мумами 70–90 °C, однако в последние годы в мире наблюдается стремительно растущий интерес к средне- и низкотемпературным амилазам с оптимумом от 50–60 °C и ниже [5, 8]. В то же время низкотемпературные амилазы отечественного производства представлены на рынке лишь одним препаратом – АмилоЛюкс-А, производитель ООО ПО «Сиббиофарм», штамм-продуцент – мицелиальный гриб Aspergillus awamori . Для расширения номенклатуры низкотемпературных амилолитических препаратов настоятельно необходим поиск новых продуцентов. Поиск продуцентов низкотемпературных ферментов традиционно ведется среди псих-рофильных и психротолерантных микроорганизмов. Ранее нами было показано, что выделяемые из холодных карстовых пещер психро-фильные и психротолерантные грибы р. Geomyces имеют хороший потенциал в качестве возможных продуцентов низкотемпературных амилаз [2].
Цель исследования : изучение влияния температуры на рост пещерных изолятов Geomyces pannorum с точки зрения перспектив их практического использования в производстве низкотемпературных амилолитических ферментов.
Объекты и методы исследования. Объектом исследования служили амилолитические изоляты мицелиальных грибов G. pannorum , выделенные из грунта известняковой карстовой пещеры «Караульная-2» (Красноярский край,
Емельяновский район, Карауленский спелеоучасток Приенисейской складчато-блоковой зоны). Протяженность пещеры составляет 540 м, глубина – 34 м, температура грунта – от +2,8 до +4,0 °С в зависимости от удаленности от входа, относительная влажность воздуха – около 95 %.
Выделение амилолитических изолятов проводили методом рассева из суспензий грунта на среде следующего состава, г/л: крахмал – 10,0; NH4NO3 – 2,0; KH2PO4 – 1,0; MgSO4·7H2O – 0,5; KCl – 0,5; FeSO4 – следовые количества; агар микробиологический – 20,0; рН = 6,5–7,0. Температура инкубации составляла +8 °C. Наличие амилолитической активности проверяли по выявляемой раствором Люголя зоне гидролиза крахмала вокруг колонии (рис. 1).
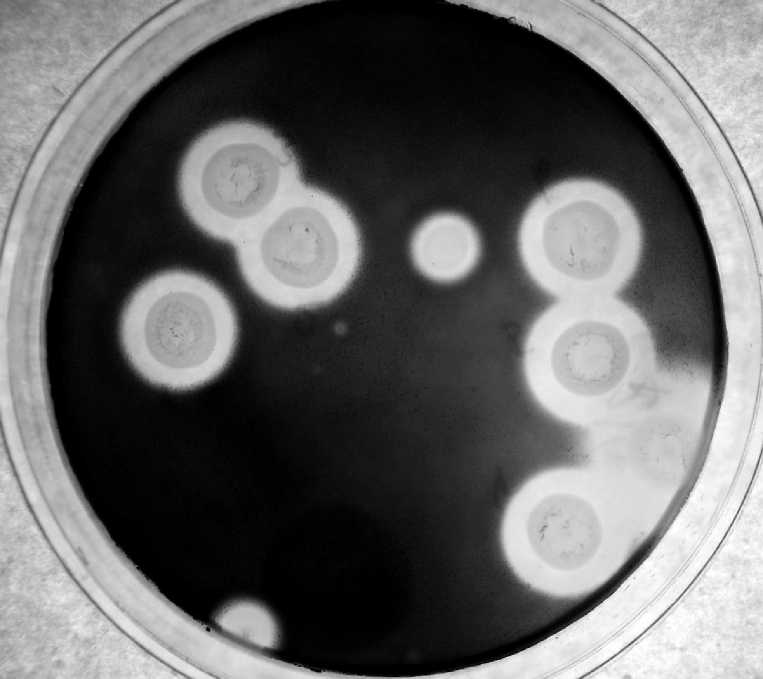
Рис. 1. Зоны гидролиза крахмала вокруг колоний G. pannorum
Видовую идентификацию проводили по культурально-морфологическим признакам по [1].
Влияние температуры на скорость роста штаммов изучали путем высева конидий на ПД-агар (пептон ферментативный – 9,0 г/л; гидролизат казеина ферментативный – 8,0; дрожжевой экстракт – 3,0; NaCl – 5,0; Na2HPO4 – 2,0; агар – 20,0 г/л; pH=7,0–7,2) с последующим параллельным культивированием при различных температурах: +9, +15, +21, +27 и +30 °C. Скорость роста при каждой температуре (мкм/ч) определяли по изменению средней длины про-ростковой гифы за интервал времени. Первое измерение проводили после прорастания конидий, второе – через 12 ч для температуры +9 °C, и через 7–9 ч для остальных температур. Измерение проводили по микрофотографиям с использованием бесплатной программы ImageJ, в каждом случае измеряли по 15–30 гиф. Масштабный коэффициент для пересчета результатов измерений в микрометры определяли с помощью объект-микрометра проходящего света ОМП ГОСТ 7513-55 производства ОАО «ЛО-МО». В качестве теоретической модели зависимости скорости роста от температуры использовали модель Ratkowsky et al. [7].
A = ( b . ( T - T^) ■ (1 - e cT T- > ))2 , (1)
где µ – скорость роста; T – температура; T max и T min – соответственно максимальная и минимальная температуры роста; b и c – коэффициенты.
Коэффициенты в моделях для каждого штамма определяли по экспериментальным данным методом наименьших квадратов с использованием пакета Stat Soft STATISTICA 6.0, модуль «Нелинейная оценка/Пользовательская регрессия». Полученные модели использовали для определения теоретических оптимальных температур роста и скоростей роста при оптимальной температуре. Микроскопические исследования проводили с помощью микроскопа «Микмед 6» (вар. 3), оснащенного цифровой камерой DCM-130E.
Результаты исследования. В общей сложности выделено и изучено 8 амилолитических штаммов, относящихся к виду G. pannorum. Верхний температурный предел роста у всех штаммов близок к +30 °C, температурные опти- мумы варьируют в диапазоне от 18,5 до 25,3 °C при среднем значении 22,2 °C. Скорость роста гифы на ПД-агаре при оптимальной температуре варьирует от 4,2 до 17,5 мкм/час на одну точку роста при среднем значении 10,2 мкм/ч.
Не выявлено связи между температурным оптимумом и скоростью роста штаммов при оптимальной температуре: соответствующий коэффициент корреляции r = 0,205, его статистическая значимость p = 0,627 (рис. 2).
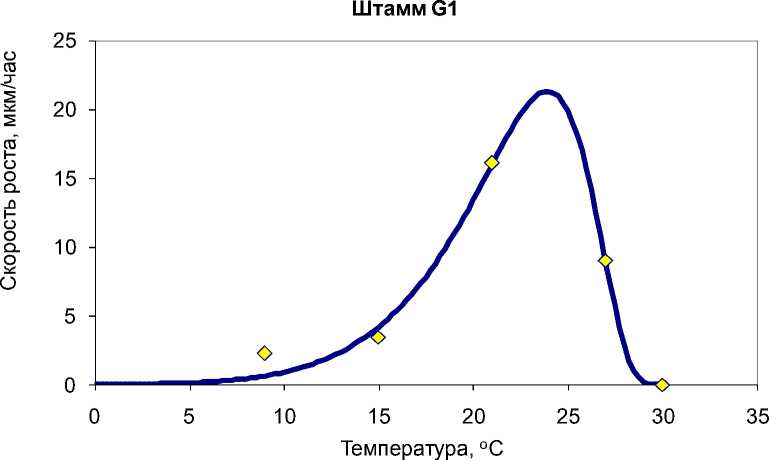
В большинстве случаев зависимость скорости роста от температуры адекватно описывается моделью Ratkowsky et al. с коэффициентами детерминации R2 от 0,987 до 0,999 (рис. 3). Однако для ряда штаммов температурные кривые отличаются от модели Ratkowsky et al. и требуют модификации модели путем возведения уравнения (1) в степень (рис. 4).
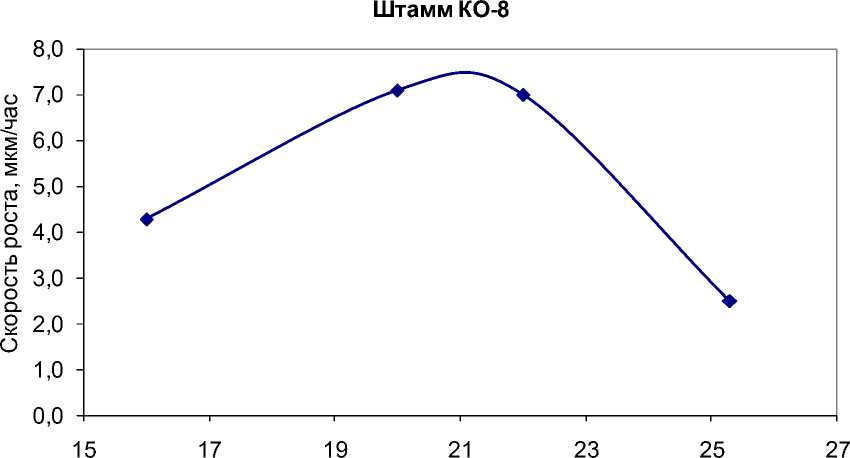
В подобных случаях для нахождения реального температурного оптимума необходимо проводить дополнительные эксперименты вблизи предполагаемого экстремума (рис. 5).
|
on 20 |
||
|
18 о 16 14 12
S 10 8 °" s 6 S. 4 . 4 2 |
♦ |
|
|
0 1 |
7 19 21 23 25 2 Оптимальная температура, ºС |
7 |
Рис. 2. Соотношение температурного оптимума и скорости роста гифы при оптимальной температуре у исследуемых штаммов
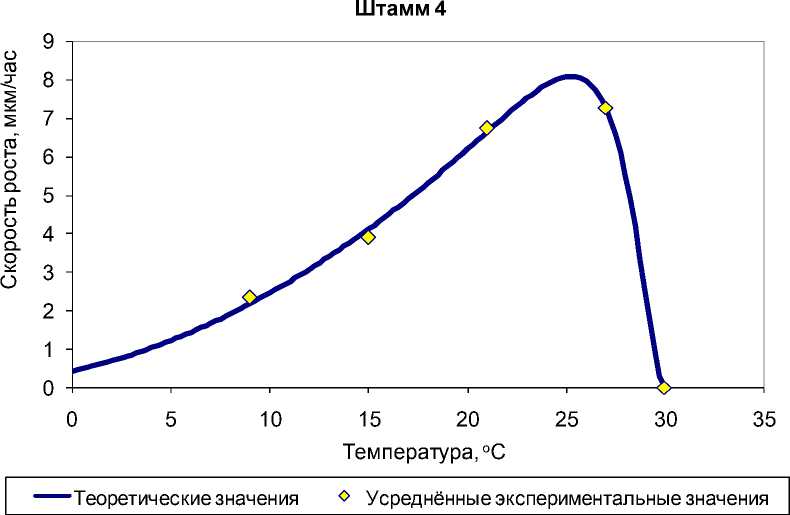
Рис. 3. Экспериментальные и теоретические (рассчитанные по модели Ratkowsky et al.) значения скорости роста G. pannorum в зависимости от температуры на примере штамма 4.
Коэффициент детерминации R2=0,998
^^^мТеоретические значения о Усреднённые экспериментальные значения
Рис. 4. Экспериментальные и теоретические значения скорости роста G. pannorum в зависимости от температуры на примере штамма G1. Для построения теоретической кривой использована 3-я степень уравнения (1). Коэффициент детерминации R2=0,981
Температура, oС
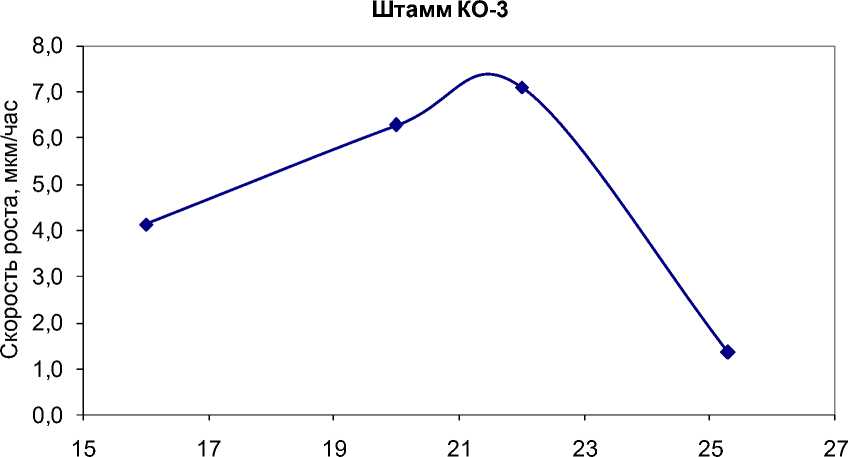
Температура, oС
Рис. 5. Экспериментальные значения скорости роста G. pannorum в зависимости от температуры вблизи температурных оптимумов для штаммов, у которых температурная зависимость отличается от модели Ratkowsky et al.
В большинстве случаев температурные кривые роста имеют асимметричный вид, при котором температурный оптимум лежит вблизи температурного максимума. Данное явление является классическим для микробных культур [7]. Одна- ко для одного штамма температурная кривая близка к параболе, хотя и для этого случая модель Ratkowsky et al. является адекватной (рис. 6).
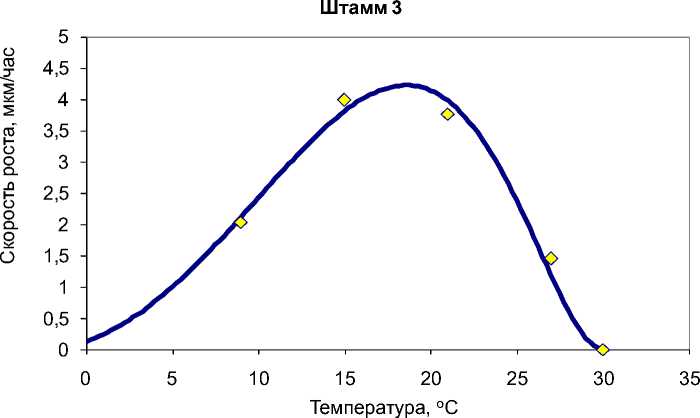
^^^^мТеоретические значения о Усреднённые экспериментальные значения
Рис. 6. Экспериментальные и теоретические (рассчитанные по модели Ratkowsky et al.) значения скорости роста G. pannorum в зависимости от температуры на примере штамма 3.
Коэффициент детерминации R2=0,987[2]
Заключение. Можно констатировать, что все выделенные из пещеры амилолитические штаммы G. pannorum по своим температурным кривым роста могут быть отнесены к психрото-лерантным либо (в связи с размытостью границ между определениями «психрофильный» и «психротолерантный») к умеренно-психро-фильным. Температурные оптимумы лежат в диапазоне, позволяющем осуществлять процесс культивирования для наработки ферментов при температуре 20–22 °C, что примерно на 15 °C ниже температуры, рекомендованной для A. awamori и других промышленных штаммов грибов-продуцентов гидролитических ферментов. Это позволяет существенно снизить энергозатраты за счет отсутствия необходимости подогрева питательной среды.
Список литературы Влияние температуры на скорость роста амилолитических штаммов Geomyces pannorum
- Егорова Л.Н. Почвенные грибы Дальнего Востока: гифомицеты. -Л.: Наука, 1986. -192 с.
- Хижняк С.В., Пампуха В.Т. Микробные сообщества карстовых пещер как потенциальный источник продуцентов низкотемпературных амилаз//Вестн. Омского ГАУ. -2016. -№ 1 (21). -С. 104-110.
- Abbas K.A., Khalil S. K., Anis Shobirin M.H. Modified Starches and Their Usages in Select-ed Food Products: A Review Study//Journal of Agricultural Science. -2010. -№ 2 (2). -P. 90-100.
- De Souza P.M., Oliveira Magalhães P. Appli-cation of microbial α-amylase in industry -A review//Brazilian Journal of Microbiology. -2010. -№ 4. -P. 850-861.
- Kuddus M., Arif Roohi J.M., Ramteke P.W. An overview of cold-active microbial α-amylase: Adaptation strategies and biotechnological po-tentials//Biotechnology. -2011. -№ 10. -P. 246-258.
- Nozière P., Steinberg W., Silberberg M., Morgavi D.P. Amylase addition increases starch ruminal digestion in first-lactation cows fed high and low starch diets//Journal of Dairy Science. -2014. -Vol. 97(4). -P. 2319-2328.
- Ratkowsky D.A., Lowry R.K., McMeekin T.A., Stokes A.N., Chandler R.E. Model for bacterial culture growth rate throughout the entire biokinetic temperature range//J. Bacteriol. -1983. -№ 154. -P. 1222-1226.
- Sarmiento F., Peralta R., Blamey J.M. Cold and Hot Extremozymes: Industrial Relevance and Current Trends//Frontiers in Bioengineering and Biotechnology. -2015. -№ 3(148). -P. 1-15.
- Tricarico J.M., Johnston J.D., Dawson K.A. Dietary supplementation of ruminant diets with an Aspergillus oryzae α-amylase//Animal Feed Science and Technology. -2008. -Vol. 145(1-4). -P. 136-150.
