Влияние температуры почвы и других факторов среды на фотосинтез сеянцев березы повислой

Автор: Холопцева Екатерина Станиславовна, Дроздов Станислав Николаевич, Сазонова Татьяна Аркадьевна, Хилков Николай Иванович
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Биология
Статья в выпуске: 8 (129) т.1, 2012 года.
Бесплатный доступ
В регулируемых условиях среды в планируемом многофакторном эксперименте изучено влияние температуры почвы (15, 20 и 25 °С) на свето-температурную характеристику двухлетних интактных сеянцев березы повислой. Наибольший потенциальный максимум нетто-фотосинтеза у сеянцев березы повислой получен при температуре почвы 15,0 °С, воздуха 20,6 °С и освещенности 34,6 клк. При двух других температурах почвы потенциальный максимум нетто-фотосинтеза был ниже, в среднем на 10 %, и достигался при более высокой освещенности и низкой температуре воздуха.
Многофакторный планируемый эксперимент, со 2-обмен, нетто-фотосинтез, температура почвы, свето-температурная характеристика
Короткий адрес: https://sciup.org/14750321
IDR: 14750321 | УДК: [58.036:581.08.132]:582.739
Effect of soil temperature on light-thermal characteristics of birch seedlings Betula pendula
The effect of the soil temperatures (15, 20 and 25 °C) on light-thermal characteristics on intact two-year birch seedlings was studied in the controlled environment in a pre-planned multifactor experiment. The greatest possible maximum of net-photosynthesis in birch seedlings was reached at soil temperature of 15 °C, air temperature of 20,6 °C, and light intensity of 34,6 klux. At two other soil temperatures, the potential maximum of the net-photosynthesis was about 10 % lower and was achieved at a higher level of light intensity and lower level of air temperature.
Текст научной статьи Влияние температуры почвы и других факторов среды на фотосинтез сеянцев березы повислой
По мере освоения северных территорий шло изучение их биологических ресурсов и развитие сельскохозяйственного производства. В конце XIX и начале XX столетия значительный материал об особенностях жизни растений в северных условиях, в том числе вечной мерзлоты, получили экспедиции переселенческого управления Министерства земледелия. Результаты их работы привлекли вначале ботаников, а в дальнейшем физиологов растений к выяснению влияния на растения экологических особенностей Севера, в частности низких температур почвы [ 6; 18-40 ] . В России первые опыты по изучению влияния пониженных температур почвы на растения проведены в 1900 году в Санкт-Петербурге в Лесном институте. В дальнейшем исследования влияния температуры почвы на жизнедеятельность растений нашли отражение во многих трудах [ 6], [7], [18 ] , в которых основное внимание уделено сельскохозяйственным растениям. Сведения о реакции древесных пород малочисленны и носят в основном описательный характер [ 6; 79-136], [9; 48 ] , в то время как возрастание роли искусственного лесовозобновления диктует необходимость всестороннего изучения экофизиологической характеристики сеянцев перспективных лесных культур.
В послеледниковый период в процессе исторического развития растительного покрова широкое распространение получили березовые формации. К настоящему времени представители рода Березы ( Betula L. ) занимают обширный ареал в умеренных и арктических зонах Европы и Северной Америки [ 3 ] . В Российской Федерации ими занято более половины всей площади, находящейся под лиственными древесными породами. В европейской части России наиболее часто встречаются береза повислая ( Betula pendula Roth. ) и береза пушистая ( Betula pubescens Ehrh .).
По занимаемой площади береза стоит на третьем месте после лиственницы и сосны, являясь одной из основных лесообразующих пород. Ценность березы определяется ее пластичностью, неприхотливостью, способностью расти и возобновляться в различных лесорастительных условиях. Она дает высококачественное сырье для различных отраслей промышленности, причем все ее части – почки, ветки, листья, береста, сок, и особенно древесина, с давних пор широко используются человеком. Помимо этого она имеет огромное санитарно-гигиеническое значение. Береза привлекает все большее внимание лесоводов. Среди видов берез имеется много форм, выделенных преимущественно по габитусу ствола, цвету и строению коры, рисунку древесины [ 13;
46 ] . Очень ценной является форма березы повис -лой с узорчатой древесиной. Целенаправленное изучение биологии берез началось в первой половине XX столетия. Но, несмотря на значительное внимание к ней как в России, так и за рубежом [ 3], [11 ] , многие вопросы остались недостаточно изучены, особенно касающиеся ее экологии на ранних этапах развития, что имеет большое значение при выращивании посадочного материла. Практически не исследовано влияние температуры почвы на сеянцы березы [ 15 ] . В то же время возрастание роли экологических показателей в решении конкретных практических вопросов требует их перевода из качественного описания в количественное. Последнее стало возможно в результате развития фитотроники и вычислительной техники, разработки методики проведения многофакторных планируемых экспериментов на базе системной идеологии и моделирования [ 8 ] . При этом, учитывая большое влияние фактора времени, в связи с ростом и развитием растений для оценки их реакции на действие внешней среды в активном эксперименте наиболее удобным интегральным показателем является первичный процесс продуктивности – СО2-обмен, быстро реагирующий на изменения условий среды и доступный для регистрации без контакта с растением, дистанционно и непрерывно [ 12 ] .
Целью данного исследования было изучение влияния температуры почвы на экофизиологиче-скую характеристику сеянцев березы повислой. В задачи работы входило проведение планируемого многофакторного эксперимента и определение уровня свето-температурных условий внешней среды, которые обеспечивают достижение потенциального максимума и зоны оптимума нетто-фотосинтеза интактных растений при естественном содержании в воздухе СО2.
МАТЕРИАЛЫ И МЕТОДЫ
Исследования проводили на территории Агробиологической станции Института биологии КарНЦ РАН. Объектами изучения являлись 2-летние сеянцы березы повислой. В качестве посевного материала использовали сертифицированные семена березы повислой от контролируемого опыления фирмы Forelia OY (Finland). В первый год исследования семена проращивали на фильтровальной бумаге в чашках Петри, пикировали в стаканчики с песком, затем подросшие растения пересаживали в грунт в условия теплицы. На второй год растения переносили на грядки. По завершении второго года развития растения выкапывали из грунта и помещали в пластиковые сосуды, по одной штуке в каждый, сохраняя в целости корневую систему, и переносили под светоустановку с люминесцентными лампами, где выдерживали в течение недели для адаптации растений к факторостатным условиям выращивания (температура воздуха – 18–
20 °С ночь/день, освещенность – 10–12 клк, длина светового периода – 16 часов).
Далее по 3 сосуда с растениями помещали в установку с регулируемыми условиями среды, позволяющую обеспечивать пределы освещенности 0–40 клк, температуру воздуха от 5 до 40 °С и почвы от 15 до 30 °С [ 8 ] , где в течение 3 дней (ежедневно изменяя температуру почвы – 15, 20 и 25 ° С) исследовали их СО2-обмен в условиях двухфакторного эксперимента [ 4 ] (табл. 1) в двукратной повторности.
Таблица 1
План постановки эксперимента по изучению зависимости нетто-фотосинтеза растений березы повислой от переменных факторов среды
|
№ |
1-й день |
2-й день |
3-й день |
Тв, ºС |
Е, клк |
|
Тп1 |
Тп2 |
Тп3 |
|||
|
1 |
15 |
20 |
25 |
12 |
15 |
|
2 |
15 |
20 |
25 |
12 |
25 |
|
3 |
15 |
20 |
25 |
20 |
15 |
|
4 |
15 |
20 |
25 |
20 |
25 |
|
5 |
15 |
20 |
25 |
20 |
40 |
|
6 |
15 |
20 |
25 |
28 |
25 |
|
7 |
15 |
20 |
25 |
28 |
40 |
Примечание. Тп1, Тп2, Тп3 – температура почвы в опытах (°С), Тв – температура воздуха, Е – интенсивность освещенности по плану эксперимента.
При определении концентрации СО2 использовали оптико-акустический инфракрасный газоанализатор Infrаlуt-IV (Германия, фирма Junkаlоr), включенный по дифференциальной схеме. Интенсивность газообмена определяли при концентрации СО2 в воздухе, близкой к естественной. После 40-минутной экспозиции на каждой ступени плана интенсивность нетто-фотосинтеза сеянцев рассчитывали по разности концентраций СО2 на входе и выходе из ассимиляционной камеры установки и скорости поступления наружного воздуха и пересчитывали на единицу сухой массы целых (интактных) растений.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Обработка полученных данных методом множественного регрессионного анализа позволила получить ряд уравнений, отражающих зависимость интенсивности нетто-фотосинтеза растений березы от факторов среды – света и температуры воздуха при трех уровнях температуры почвы.
NP = b0 + b1E +b2T + b3ET + b4E2 + b5T2, где NP – интенсивность видимого фотосинтеза, мг СО2/г ч; Е – освещенность, клк; Т – температура воздуха, °С; b0–b5 – коэффициенты, определенные при математической обработке экспериментальных данных. Статистическая оценка уравнений показала высокую степень их достоверности: коэффициенты множественной детерминации R2 > 0,80, критерии Фишера
F = 3,15–5,04, что во всех вариантах опыта больше F табличного при уровне значимости a = 0,05.
В естественных условиях сочетание факторов внешней среды, поддерживающее максимальные значения фотосинтеза, бывает очень неустойчиво. Поэтому наилучшими условиями произрастания любого вида являются те, которые обеспечивают достижение достаточно высоких значений оптимума, – температура и освещенность, позволяющие получить более 90 % от потенциального максимума интенсивности нетто-фотосинтеза. Конкретные границы оптимума зависят от вида и фазы развития растений и сопутствующих условий внешней среды.
Поэтому при анализе уравнений связи мы определяли не только максимальные значения интенсивности видимого фотосинтеза растений, но и области его оптимума (90 % от максимума) и интенсивность света и температуры, обеспечивающие их достижение.
Как показывает анализ полученных уравнений (табл. 2), судя по относительно небольшой разнице потенциального максимума нетто-фотосинтеза (в пределах 10 %) при естественном содержании в воздухе СО2 исследуемые температуры почвы практически входят в зону оптимальных температур почвы для данного экотипа березы. Наибольшего максимума нетто-фотосинтеза интактные растения двухлетних сеянцев березы повислой достигают при температуре почвы 15 °С и определенных температуре воздуха и освещенности – 20,6 °С и 34,6 клк. При более высокой температуре почвы (20–25 °С) максимум нетто-фотосинтеза тех же сеянцев березы ниже, чем при температуре почвы 15 °С и достигается при более низкой температуре воздуха и более высокой освещенности. Снижение видимого фотосинтеза при повышении температуры почвы, возможно, происходит в результате возрастания дыхания корневой системы [ 14 ] , но при этом оптимум нетто-фотосинтеза достигается в более широком световом и температурном диапазоне. Ранее в работе финских исследователей [ 15 ] было показано, что низкие температуры почвы (5–10 °С) отрицательно сказываются на росте трехлетних сеянцев березы повислой, снижая ее фотосинтез, сухой вес листьев и изменяя ряд физиологических показателей. Такие явления, вероятно, связаны с тем, что локальное охлаждение корневой системы вызывает закаливающий эффект во всем растении, как это было показано на примере пшеницы [ 2 ] .
Изучение влияния температуры воздуха на видимый фотосинтез сеянцев березы при двух уровнях температуры почвы и разной освещенности – из района потенциального максимума и областей предполагаемого оптимума – свидетельствует о значительном воздействии всех исследуемых факторов среды на видимый фотосинтез растений березы. При температуре по- чвы 15 °С наиболее сильное влияние на нетто-фотосинтез сеянцев оказывает температура воздуха, о чем можно судить по углу наклона кривых и их сближенности при всех рассматриваемых уровнях освещенности. При этом, во всем диапазоне исследованных температур воздуха повышенная освещенность – 45 клк – при температуре почвы 15 °С снижает эффективность фотосинтеза, что свидетельствует об изменении характера влияния этой интенсивности света на фотосинтетический аппарат. Скорее всего, происходит ее выход за пределы зоны светового оптимума в данном сочетании с уровнями напряженности сопутствующих факторов внешней среды, как и в случае интенсивности света 40 клк при повышенной температуре воздуха. В литературе имеются данные, что при увеличении освещенности вначале происходит перестроение хлоропластов с изменением их угла направленности к световому потоку [19]. При дальнейшем усилении интенсивности света возможно образование синглетного кислорода, что ведет к саморазрушению клетки [5], [16], [17].
При температуре почвы 25 °С сила влияния температуры воздуха на видимый фотосинтез значительно снижается и возрастает необходимость в усилении освещенности до 45–50 клк. При более высоком или пониженном уровне освещенности нетто-фотосинтез сеянцев снижается.
Таблица 2
Потенциальный максимум и оптимум нетто-фотосинтеза сеянцев березы повислой и свето-температурные условия среды, обеспечивающие их достижение при трех уровнях температуры почвы
|
Тп |
Интенсивность нетто-фотосинтеза (NP) |
Условия максимума |
Условия оптимума |
|||
|
max |
оpt |
E |
Т |
E |
Т |
|
|
°С |
мг СО2/г ч |
мг СО2/г ч |
клк |
°С |
клк |
°С |
|
15 |
4,3 |
3,9 |
34,6 |
20,6 |
26,7–42,5 |
16,3–25,0 |
|
20 |
3,8 |
3,4 |
63,8 |
18,7 |
44,6–83,0 |
11,2–24,3 |
|
25 |
3,9 |
3,5 |
81,5 |
13,0 |
>39 |
5,6–29,6 |
Примечание. Тп – температура почвы, Е – интенсивность освещенности, Т – температура воздуха.
Графики по влиянию освещенности на нетто-фотосинтез сеянцев березы, построенные по экспериментальной модели (рис. 2А, Б), имеют в основном куполообразную форму, что не соответствует данным, приведенным в литературе [1; 128-132], полученным при изучении растений в природных условиях или в однофакторных экспериментах. По мнению авторов, при определенной интенсивности света кривые видимого фотосинтеза выходят на плато в результате достижения уровня светового насыщения. Этому утверждению противоречат данные о влиянии избыточной освещенности на появление синглетного кислорода и его разрушительного действия на фотосинтетический аппарат в результате развития окислительного стресса [10]. По нашему мнению, выход световой кривой нетто-фотосинтеза на плато является результатом не светового насыщения, а действия какого-либо лимитирующего фактора: в природе это, вероятнее всего, недостаток влаги, а в эксперименте – температуры.
Анализ световых кривых подтверждает взаимовлияние на нетто-фотосинтез исследуемых факторов внешней среды, в том числе температуры почвы. При этом у световых кривых более рельефно выделяется зона оптимума температуры воздуха в зависимости от температуры почвы. При температуре почвы 15 ° С, воздуха -12 и 28 ° С из зоны оптимума переходят в соседние зоны холодового и теплового закаливания (см. рисунок). При 25 ° С в зону холодового закаливания попадают температуры ниже 12 ° С. Но если при температуре почвы 15 ° С экспериментальные точки попадают в центр зоны светового оптимума, то при температуре почвы 25 ° С необходим более высокий уровень света, и поэтому экспериментальные точки не покрывают всей зоны, что частично компенсируется возможностями модели.
ЗАКЛЮЧЕНИЕ
Таким образом, проведенные исследования показывают значительное влияние температуры почвы на сеянцы березы повислой и ее взаимосвязь с ведущими факторами внешней среды. Потенциального максимума нетто-фотосинтеза растения достигают при температуре почвы 15,0 °С, воздуха - 20,6 °С, освещенности 34,6 клк и естественном содержании в воздухе СО2. При этом сеянцы березы имеют широкий диапазон оптимума по всем рассмотренным факторам внешней среды, обеспечивающий 90 % уровень видимого фотосинтеза при температуре воздуха 16,7-25,0 °С, освещенности 26,7-42,5 клк, кото- рый значительно расширяется в зоне оптимальных значений фотосинтеза (табл. 2).
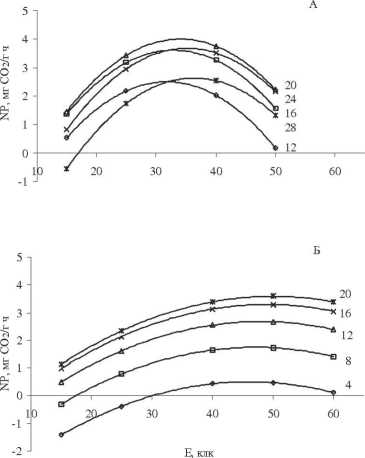
Влияние интенсивности света на видимый фотосинтез двухлетних сеянцев березы повислой в зависимости от температуры почвы и воздуха: А – при температуре почвы 15 °С и воздуха: 12 °С, 16 °С, 20 °С, 24 °С, 28 °С; Б – при температуре почвы 25 °С и воздуха: 4 °С, 8 °С, 12 °С, 16 °С, 20 °С
Для ускоренного получения сеянцев березы целесообразно, особенно в северных условиях ее выращивания, оборудовать рассадные теплицы возможным подогревом почвы в холодный период вегетации. Этого можно достигнуть закладкой по всей площади теплицы на глубине 30 см параллельно через 25–30 см оцинкованной стальной проволоки диаметром 3–4 мм. Ее концы присоединяют к клеммам трансформатора с регулируемым напряжением порядка 10–15 вольт при контроле уровня температуры почвенными термометрами.
Список литературы Влияние температуры почвы и других факторов среды на фотосинтез сеянцев березы повислой
- Березина Н. А., Афанасьева Н. Б. Экология растений. М.: Академия, 2009. 400 с.
- Венжик Ю. В., Титов А. Ф., Таланова В. В., Назаркина Е. А. Влияние охлаждения корней на устойчивость клеток листьев и активность фотосинтетического аппарата пшеницы//Доклады РАН. 2009. Т. 27. № 3. С. 314-316.
- Ветчинникова Л. В. Береза. Вопросы изменчивости. М.: Наука, 2004. 199 с.
- Голикова Г. И., Панченко Л. А., Фридман М. З. Каталог планов второго порядка. Вып. 47. Ч. 1. М.: МГУ, 1974. 387 с.
- Головко Т. К., Дымова О. В., Яцко Я. Н., Захожий И. Г., Далькэ И. В., Табаленкова Г. И. Механизмы адаптации фотосинтетического аппарата к стрессовым воздействиям//Материалы Всеросс. науч. конф. «Устойчивость организмов к неблагоприятным факторам внешней среды». Иркутск, 2009. С. 89-92.
- Дадыкин В. П. Особенности поведения растений на холодных почвах. М.: Изд-во АН СССР, 1952. 276 с.
- Коровин А. И. Роль температуры в минеральном питании растений. Л.: Гидрометиздат, 1972. 282 с.
- Ку рец В. К., Попов Э. Г. Статистическое моделирование системы растение -среда. Л.: Наука, 1991. 152 с.
- Ку усела К. Динамика бореальных хвойных лесов. Хельсинки: Repola, 1991. 210 с.
- Мокроносов А. Т., Гавриленко В. Ф. Фотосинтез. Физиолого-экологические и биохимические аспекты. М.: МГУ, 1992. 240 с.
- Новицкая Л. Л. Карельская береза: механизмы роста и развития структурных аномалий. Петрозаводск, 2008. 143 с.
- Суворова Г. Г. Фотосинтез хвойных деревьев в условиях Сибири. Новосибирск: Гео, 2009. 195 с.
- Тренин В. В. Ценные формы деревьев. Петрозаводск: Карелия, 1999. 88 с.
- Цельникер Ю. Л. Дыхание корней и их роль в углеродном балансе древесных//Лесоведение. 2005. № 6. С. 11-18.
- Aphalo P. J., Lahti M., Lehto T., Repo T., Rummukainen A., Mannerkoski H., Finer L. Responses of silver saplines to low soil temperature//University of Joensuu. Faculty of Forestry. 2006. Vol. 40. № 3. P. 429-442.
- Bennett J. Protein phosphorilation in green plant chloroplasts//Ann. Rev. Plant Rhysiol. 1991. Vol. 42. P. 281-311.
- Powles S. B. Photoinhibition of photosynthesis induced divisible//Ann. Rev. Plant Physiology. 1984. Vol. 35. P. 15-44.
- Went D. J. Plant growth under controlled conditions. Thermoperiodicity in growth and a fruiting of the tomato//Am. J. Bot. 1944. Vol. 31. P. 135-150.
- Witiak S. M., Williams W. E., Sorton H. B. Chloroplast pigments in the field//Plant, Cell and Environ. 2003. 26. № 12. P. 2005-2014.
