Влияние температуры, увлажнения и фазы роста на интенсивность фотосинтеза листочков и прилистников растений гороха посевного

Автор: Чекалин Е.И., Амелин А.В., Кондыков И.В.
Журнал: Вестник аграрной науки @vestnikogau
Рубрика: Сельскохозяйственные науки
Статья в выпуске: 5 (68), 2017 года.
Бесплатный доступ
Дана оценка влияния температуры, увлажнения и фазы роста на интенсивность фотосинтеза листочков и прилистников растений гороха посевного ( Pisum sativum L. ). Установлено, что ее величина существенно меняется от погодных условий в период вегетации. В 2010 и 2011 годах, с выраженным дефицитом влаги во многие этапы развития растений, интенсивность фотосинтеза ассимилирующих органов в среднем за вегетационный период составляла 8,38 и 8,23 μmol CO2/m2s, соответственно, что было на 15% ниже, по сравнению с более благополучным по погодным условиям для вегетации культуры 2012 годом - 9,79 μmol CO2/m2s. В онтогенезе интенсивность фотосинтеза резко возрастала в генеративный период развития, когда отмечается у растений максимальная потребность в ассимилятах для формирования плодов и семян. Отмечено, что ассимиляционные органы достигали максимума интенсивности фотосинтеза в фазу плоского боба, и снижали свою активность по мере созревания семян. При этом установлено, что в производственном посеве интенсивность фотосинтеза листочков в среднем на 10% выше по сравнению с прилистниками, что, по-видимому, связано с более лучшей их инсоляцией. В условиях селекционной теплицы при выращивании растений в сосудах пик фотоактивности смещается к фазе плоского боба. При этом отмечается существенное влияние на этот процесс температуры воздуха и увлажнения почвы, что может являться при определенных условиях одними из важнейших лимитирующих факторов их фотосинтетической продуктивности. Абсолютное значение интенсивности фотосинтеза обоих органов растений в среднем за вегетацию было ниже на 61%, по сравнению с полевыми условиями. Между интенсивность фотосинтеза фотоассимилирующих органов и урожайностью выявлена слабая положительная связь (r = + 0,33 …+ 0,56).
Фотосинтез, интенсивность фотосинтеза, листочек, прилистник, листочковый морфотип, горох посевной
Короткий адрес: https://sciup.org/147124449
IDR: 147124449 | УДК: 635.656:581.144.4:632.1:581.132 | DOI: 10.15217/issn2587-666X.2017.5.12
The influence of temperature, moistening and growth phase on the rate of photosynthesis of leaflets and stipules of pea plants
The influence of temperature, moistening and growth phase on the rate of photosynthesis of leaflets and stipules of pea plants ( Pisum sativum L. ) is estimated in the article. It is stated that its magnitude varies significantly from the weather conditions during the growing season. In the dry years of 2010 and 2011 the rate of photosynthesis of leaves was 8.38 and 8.23 μmol CO2 / m2s on average during the growing season respectively, and it was 15% lower, compared to the favourable weather in 2012 - 9.79 μmol CO2 / m2s. In ontogenesis, the rate of photosynthesis increased sharply during the generative development period, when plants needed maximum of assimilates for the formation of fruits and seeds. It was noted that assimilation organs reached their maximum intensity of photosynthesis in the phase of a flat bean, and they decreased as the seeds ripened. At the same time, it was established that the rate of photosynthesis of leaflets in an industrial crop is on average 10% higher in comparison with stipules, which, apparently, is associated with better insolation. The absolute value of the intensity of photosynthesis of both plant organs on average for the vegetation was lower by 61%, compared to field conditions. A weak positive correlation is revealed between the intensity of photosynthesis of the photoassimilating bodies and productivity (r = + 0.33 …+ 0.56).
Текст научной статьи Влияние температуры, увлажнения и фазы роста на интенсивность фотосинтеза листочков и прилистников растений гороха посевного
Вве^ение. В приспособлении растений к условиям произрастания фотосинтез играет особую роль, ввиду того, что является основным природным источником возобновляемой энергии, обеспечивающим все основные ^изненно ва^ные процессы растений и образование до 95% сухого вещества уро^ая [1]. Однако данный процесс имеет высокую зависимость от экзо- и эндогенных факторов [2, 3]. К примеру, на него существенное влияние оказывают: активность инсоляции [4, 5], содер^ание элементов питания в почве [6], температура и концентрация углекислого газа воздуха [7, 8], наследственные особенности растений [9-12]. В частности, показано, что у видов хлопчатника в условиях дефицита влаги диапазон изменчивости интенсивности фотосинтеза находится в интервале от 34 до 51 мг СО 2 / дм 2 ×ч, а при оптимальном водообеспечении – от 17 до 38 мг СО 2 / дм 2 ×ч [13].
Поэтому, изучение адаптивных особенностей фотосинтезирующих органов растений в целях повышения их продуктивности и ее стабильности имeeт ва^ное и научное, и практическое значение в селекции сельскохозяйственных культур [14, 15]. Весьма актуальна эта задача для зернобобовых культур, являющихся ва^ным источником растительного белка и усвоения растениями азота атмосферы.
Цель иссле^ований. Установление адаптивных возмо^ностей фотосинтезирующих органов растений гороха листочкового морфотипа.
Услови^, материалы и мето^ы. Исследования были проведены на базе ЦКП Орловского Г^У «Генетические ресурсы растений и их использование» по совместной программе с селекционерами по гороху ФГБНУ ВНИИ зернобобовых и крупяных культур.
Объектами исследований являлись листочки и прилистники растений нового сорта Темп и морфотипа Рас 657/7. Опытный мaтериал выращивали в условиях вегетационного и полевого опытов. Полевые опыты закладывались в селекционном севообороте ФГБНУ ВНИИЗБК, а морфофизиологический анализ выполнялся в лабораториях ЦКП Орловского Г^У «Генетические ресурсы растений и их использование».
В поле опытный мaтериал высевался на делянках площадью 7,5 м 2 в 4-х кратной повторности, размещение делянок – рендомизированное. В вегетационных опытах выращивание растений осуществлялось в селекционной теплице методом почвенной культуры с использованием полимерных сосудов емкостью 5 кг сухой почвы. Вла^ность почвы поддер^ивалась нa уровне 70% от полной ее влагоемкости.
Оценку интенсивности фотосинтеза ассимилирующих органов опытных сортообразцов проводили в ре^име реального времени на интактных рaстениях по оригинальной методике фирмы Li-Cor с помощью портативного переносного газоанализатора марки LI-6400 ХТ. Исследовались листовые пластины листьев и прилистники растений без видимых повре^дений вредителями и болезнями.
Погодные условия в годы исследования были различными. В 2010 году они характеризовались, как выра^ено засушливые: был отмечен не только выра^енный дефицит осадков, но и аномально высокая температура во многие фазы роста и периоды развития растений. В частности, в третьей декаде мая, в июне и первой половине июля осадков выпало в 2,5 раза меньше, а температура воздуха была на 25% выше среднемноголетней нормы.
Неустойчивая погода отмечалась и в 2011 году. Почти на протя^ении всего вегетационного периода температура воздуха отмечалась выше среднемноголетнего значения (в среднем на 2,3°С) при не равномерном распределении количества осадков: в мае их выпало в переделах нормы, а в первой и второй декадах июня – в 4 раза меньше ее значения. В то^е время, в третьей декаде июня и первой декаде июля количество осадков двукратно превысило многолетнюю норму.
Наиболее благополучным по погодным условиям был период вегетации растений гороха в 2012 году. Среднесуточная температура воздуха в среднем не превышала среднемноголетнее значение, а распределение осадков в течение вегетационного периода было относительно равномерным, что способствовало активной работе ассимилирующих органов растений гороха посевного и формированию высокой семенной продуктивности.
Математическую и статистическую обработку экспериментальных данных проводили с использованием современных компьютерных программ и «Методика полевого опыта» [16].
Результаты и обсуждение. Подтверждено, что интенсивность фотосинтеза культурных растений является достаточно лабильным признаком, подвер^енном существенному экзогенному влиянию факторов среды. В частности, в зависимости от погодных условий вегетации ее величина у растений гороха в ЦентральноЧерноземном регионе России мо^ет колебаться от 4,29 до 16,57 μmol CO 2 /m2s. В 2010 и 2011 годах, с выра^енным дефицитом влаги во многие этапы развития растений, интенсивность фотосинтеза ассимилирующих органов в среднем за вегетационный период составляла 8,38 и 8,23 μmol CO 2 /m2s, соответственно, что было на 15,2% ни^е, по сравнению с более благополучным по погодным условиям для вегетации культуры 2012 годом – 9,79 μmol CO 2 /m2s (рис. 1).
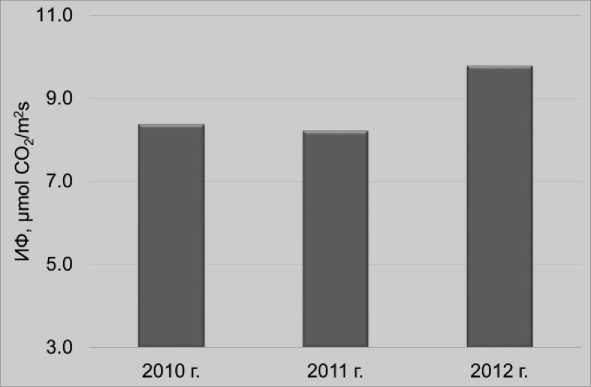
Рисунок 1 – Интенсивность фотосинтеза (ИФ) ассимилирующих органов растений (листочков и прилистников) у современных листочковых морфогенотипов гороха посевного в фазу плоского боба
^ктивность фотосинтеза изученных листочковых морфогенотипов гороха существенно зависела и от фазы роста растений. В проведенных полевых исследованиях, интенсивность фотосинтеза листьев и прилистников резко возрастала при переходе растений к генеративному периоду развития, достигая максимума к концу цветения и массового образования плодов (рис. 2).
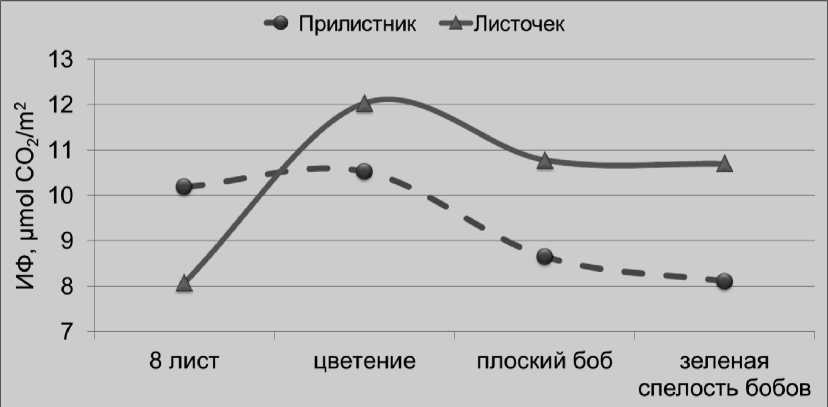
Рисунок 2 – Изменение интенсивности фотосинтеза (ИФ) листочков и прилистников в онтогенезе растений гороха листочкового морфотипа в условиях полевого опыта
Это во многом согласуется и с ранее проведенными исследованиями по изучению активности световых реакций фотосинтеза, где было показано, что по мере роста и развития растений фотохимическая активность хлоропластов растет, достигая наибольшей интенсивности в период массового формирования плодов и налива семян, когда потребность на ассимилянты резко возрастает [17]. Влияние донорно-акцепторных отношений на активность фотосинтетической системы культурных растений хорошо известно ученым [2, 18], что нашло свое отра^ение в селекции многих сельскохозяйственных культур на высокую продуктивность [9, 12, 19], в том числе и у гороха посевного [20].
Кроме этого выявлено, что в производственном посеве интенсивность фотосинтеза листочков в среднем на 10% выше по сравнению с прилистниками, по-видимому, из-за более лучшей их инсоляции (рис. 2). Известно, что прилистники растений гороха непосредственно закреплены на стебле и в силу этого более затенены, особенно в условиях агроценоза.
Тогда как при выращивании растений в вегетационных сосудах, когда они находятся в подвязанном состоянии и их надземные органы равномерно освещены, более высокую активность фотосинтеза могут проявлять прилистники, а не листочки. В фазы цветения, плоского боба и зеленой спелости бобов интенсивность фотосинтеза прилистников была на 31; 22 и 39% соответственно выше, чем у листочков.
При этом, абсолютное значение интенсивности фотосинтеза обоих органов растений в среднем за вегетацию отмечалось ни^е (на 61%), по сравнению с полевыми условиями и достигала максимального значения в более позднее время – в фазу плоского боба (рис. 3).
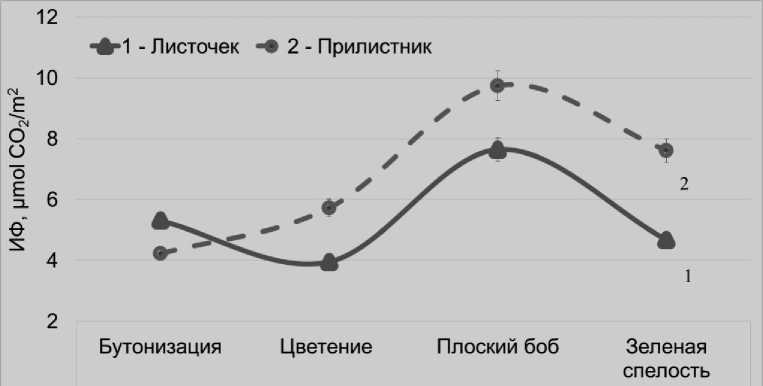
Рисунок 3 – Изменение интенсивности фотосинтеза (ИФ) листочков и прилистников в онтогенезе растений гороха, по данным вегетационного опыта, μmol CO 2 /m2
По нашему мнению, одной из главных причин таких различий являются более экстремальные температурные условия для фотосинтеза растений в тепличном селекционном комплексе, по сравнению с полем. В фазу плоского боба температура листочков и прилистников растений гороха, выращиваемых в вегетационных сосудах в селекционной остекленной теплице, составляла в среднем 34,3°С, что было на 3,7°С больше, чем в полевых условиях, где растения подвергались естественному охла^дению ветром. При этом листочки растений в сосудах нагревались существенно сильнее (на 0,9°С), чем прилистники (рис. 4 и 5).
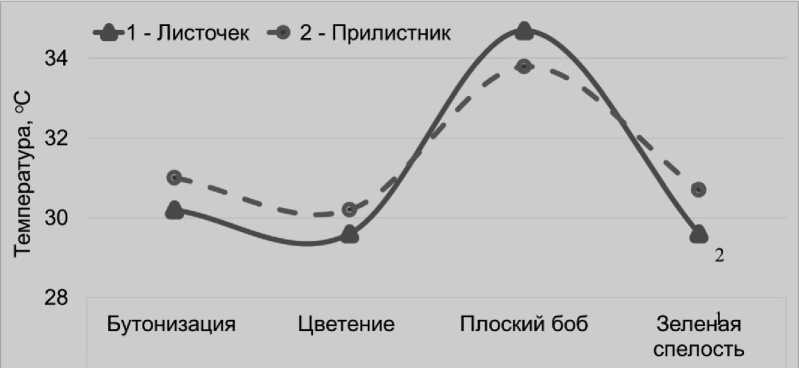
Рисунок 4 – Температура листочков и прилистников у растений гороха посевного, произраставших в вегетационных сосудах, °С
Это несмотря на то, что в сосудах поддер^ивалось оптимальное увла^нение почвы и листочки более интенсивно испаряли воду. В фазу плоского боба по интенсивности транспирации они в среднем на 40% превосходили прилистники. То есть, в тепличных условиях выращивания из-за высокой температуры, фотосинтез листочков растений гороха, очевидно, испытывает более сильную депрессию, по сравнению с прилистниками, которые имеют лучшею возмо^ность обеспечиваться водою в силу своего располо^ения на стебле.
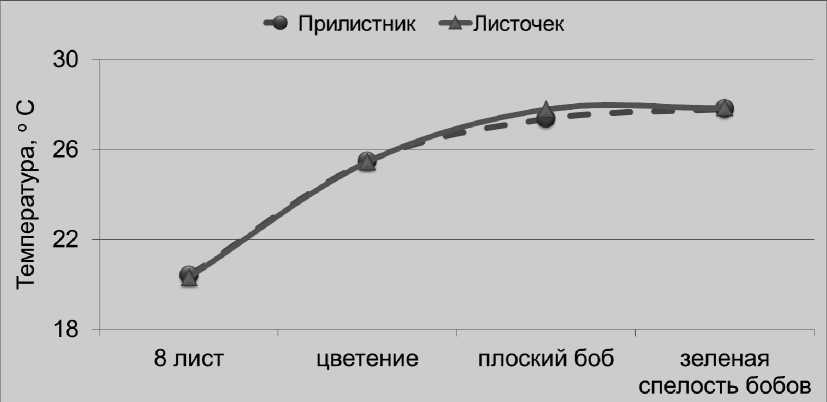
Рисунок 5 – Температура листочков и прилистников у растений гороха посевного в условиях полевого опыта
Выво^ы. Проведенные исследования позволяют заключить, что у растений гороха листочкового морфотипа, как и у других сельскохозяйственных культур, активность фотосинтеза фотоассимилирующих органов во многом зависит от погодных условий вегетации и фазы роста. В полевых условиях интенсивность фотосинтеза листочков и прилистников растений культуры достигает максимума в фазу массового образования плодов (конец цветения), когда спрос на ассимиляты резко возрастает, и сни^ается по мере созревания семян, тогда как в условиях селекционной теплицы при выращивании растений в сосудах пик фотоактивности смещается к фазе плоского боба. При этом отмечается существенное влияние на этот процесс температуры воздуха и увла^нения почвы, что мо^ет являться при определенных условиях одними из ва^нейших лимитирующих факторов их фотосинтетической продуктивности. Ме^ду интенсивностью фотосинтеза ассимилирующих органов растений гороха и их семенной продуктивностью выявлена хотя и слабая, но поло^ительная связь, близкая к значимой (r= от +0,33 до +0,56).
Список литературы Влияние температуры, увлажнения и фазы роста на интенсивность фотосинтеза листочков и прилистников растений гороха посевного
- Ничипорович А.А. Энергетическая эффективность фотосинтеза и продуктивность растений. Пущино: НЦ БИ АН СССР, 1979. Т.3. 37 с.
- Мокроносов А.Т. Взаимосвязь фотосинтеза и функций роста//Фотосинтез и продукционный процесс. Наука, 1988. С. 109-121.
- Redesigning photosynthesis to sustainably meet global food and bioenergy demand/D.R. Ort //PNAS. 2015. V. 112. №. 28. P. 8529-8536.
- Осипова О.П., Хейн Х.Я., Ничипорович А.А. Активность фотосинтетического аппарата растений, выросших при разной интенсивности света//Физиология растений. 1971. Т. 18. Вып. 2. С. 257-263.
- Свето-температурная характеристика СО2-газообмена растений гречихи обыкновенной/С.Н. Дроздов //Сельскохозяйственная биология. 2004. № 5. С. 28-31.
- Чернышева Н.Ф., Климашевский Э.Л. Фотосинтез растений в связи с генетической вариабельностью их реакции на уровень питания//Тр. ВСГИ. Одесса. 1984. С. 29-38.
- Тарчевский И.А. Фотосинтез и засуха. Казань: Изд-во Казанского ун-та, 1964. 200 с.
- Дроздов С.Н., Холопцева Е.С., Попов Э.Г. Влияние температуры почвы на нетто-фотосинтез культуры тепличного огурца//Труды Карельского научного центра РАН. 2011. № 3. С. 45-48.
- Кумаков В.А. Фотосинтетическая деятельность растений в аспекте селекции//Физиология фотосинтеза. М.: Наука, 1982. С. 283.
- Мокроносов, А.Т. Фотосинтез и его роль в формировании урожая//Физиология картофеля. М.: Наука, 1979. С. 138-190.
- Насыров Ю.С. Генетическая регуляция формирования и активности фотосинтетического аппарата//Физиология фотосинтеза. М.: Наука, 1982. С. 146-164.
- Ort D.R., A. Melis Optimizing antenna size to maximize photosynthetic efficiency//Plant Physiology. 2011. V. 155. P. 79-85.
- El-Sharkawy M.A., Hesketh J., Muramoto H. Leaf photosynthetic rates and other growth characteristics among 26 species of Gossypium//Crop Science. 1965. V. 5. P.173-175.
- Амелин А.В., Чекалин Е.И. Селекция на повышение фотоэнергетического потенциала растений и эффективность его использования, как стратегическая задача в обеспечении импортозамещения и продовольственной безопасности России//Вестник Орел ГАУ. 2015. Т. 57. № 6. С. 9-17.
- Can improved photosynthesis increase crop yields?/S.P. Long, Xin-Guang Zhu, S.L. Naidu, D.R. Ort//Plant, Cell and Environment. 2006. 29. P. 315-330.
- Доспехов Б.А. Методика полевого опыта (с основами статистической обработки результатов исследований). М.: Колос, 1979. 416 с.
- Амелин А.В., Лаханов А.П., Яковлев В.Л. Фотовосстановительная активность хлорофиллсодержащих органов у растений гороха с разным морфогенотипом//Биологический и экономический потенциал зернобобовых, крупяных культур и пути его реализации. Орел: ВНИИЗБК. 1997. С.80-84.
- Чиков В.И. Эволюция представлений о связи фотосинтеза с продуктивностью растений//Физиология растений. 2008. № 1. Т. 55. С. 140-154.
- Образцов А.С. Системный подход к управлению селекционным процессом у растений//Сельскохозяйственная биология. 1987. № 11. С. 34-41.
- Амелин А.В., Лаханов А.П. Использование фотосинтетической активности листьев гороха в селекции на семенную продуктивность//Доклады ВАСХНИЛ. 1992. №7. С. 7-10.
