Влияние температуры воды и количества осадков на численность преимагинальной гемипопуляции малярийного комара (комплекс видов Anopheles maculipennis Meigen, 1818) в условиях урбанизированной среды обитания
 в условиях урбанизированной среды обитания")
Автор: Колпаков А.Д.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Водные экосистемы
Статья в выпуске: 2 т.10, 2008 года.
Бесплатный доступ
Оценивается влияние температуры воды и количества осадков на численность преимагинальной геми- популяции малярийного комара в условиях г. Ижевска.
Короткий адрес: https://sciup.org/148198124
IDR: 148198124 | УДК: 595.7-15
The influence of water temperature and rainfall quantity on the praeimaginal hemipopulanion of malarial mosquito (species complex Anopheles maculipennis Meigen, 1818) quantity in conditions of urban environment
The influence of water temperature and rainfall quantity on the praeimaginal hemipopulation of malarial mosquito quantity in conditions of the Izhevsk-city is estimating.
Текст научной статьи Влияние температуры воды и количества осадков на численность преимагинальной гемипопуляции малярийного комара (комплекс видов Anopheles maculipennis Meigen, 1818) в условиях урбанизированной среды обитания
ГОУ ВПО «Удмуртский государственный университет»
Оценивается влияние температуры воды и количества осадков на численность преимагинальной гемипопуляции малярийного комара в условиях г. Ижевска.
Температура воды, форма и количество осадков – важнейшие абиотические факторы, определяющие численность личинок малярийного комара. Температура воды определяет возможность и сроки развития преима-гинальных стадий в данном водоеме, а количество осадков, выпавших за определенный период, оказывает влияние на величину ано-фелогенной зоны водоема и может изменять направление действия температурного фактора. В связи с этим имеет смысл рассматривать их совокупное влияние на численность личинок. В урбоэкотопах температурный режим и режим осадков существенно отличаются от таковых индигенных экотопов. Поэтому и характер влияния этих факторов на численность преимагинальной гемипопуляции малярийного комара будет отличаться
Материалы и методика исследований
Исследования проводились в сезонах1 2004-2007 г.г. на трех анофелогенных водоемах, расположенных в пределах административно-территориальных границ г. Ижевска.
Водоем 1 (средняя за период наблюдений площадь 2372 м2) представляет собой водоем прудового типа и расположен на пересечении улиц Сабурова и Автозаводской.
Водоем 2 (средняя за период наблюдений площадь 712 м2) представляет собой дренажную канаву, располагающуюся вдоль железной дороги «Ижевск-Воткинск» и соединяющуюся с р. Иж. Отлов личинок производился в предустьевом участке канавы.
Водоем 3 (средняя за период наблюдений площадь 1186 м2) представляет собой водоем карьерного типа и расположен возле стройплощадки ТЭЦ-3 в пойме р. Позими.
Отлов личинок в течение сезона производился ежедекадно кюветой площадью 0,0167 м2. Плотность личинок рассчитывалась на 1 м2 анофелогенной площади водоема [1]. Отловленные личинки фиксировались в 70%-м растворе этилового спирта для дальнейшего определения возраста и видовой диагностики по личинкам IV возраста [1; 2].
Температура на поверхности воды измерялась ртутным химическим термометром с точностью измерений 0,1°С ежедекадно в контрольной точке в пределах анофелогенной площади. Данные о подекадной сумме осадков предоставлены Удмуртским центром гидрометеорологии.
Видовая диагностика личинок проводилась с использованием определителя С.Н. Заречной и Е.В. Маханько [2].
Математическая обработка данных проводилась на персональном компьютере с использованием пакета анализа Microsoft Excel. Также использовались учебное пособие Г.Ф. Лакина [3] и руководство В. Боровикова [4].
Интерпретация результатов математического анализа проводилась, исходя из положе- ния о том, что самки малярийного комара откладывают яйца более или менее равномерно в течение всего сезона.
Результаты и обсуждение
Все собранные насекомые относятся к ком- плексу видов Anopheles maculipennis.
В таблице 1 приведены результаты двухфакторного дисперсионного анализа влияния температуры воды и количества осадков на численность личинок малярийного комара.
Таблица 1 . Результаты двухфакторного дисперсионного анализа влияния температуры воды и количества осадков на численность личинок малярийного комара
|
№ водоема |
Источник вариации |
Дисперсия |
Критерий Фишера F |
Уровень значимости p |
Критическое значение критерия Фишера Fкр |
Сила влияния, % |
|
1 |
Температура воды |
12,78 |
17,54 |
0,0004 |
4,3 |
2,2 |
|
Подекадная сумма осадков |
189,88 |
14,14 |
0,001 |
4,3 |
32,91 |
|
|
Температура воды и подекадная сумма осадков |
272,98 |
2,69 |
0,13 |
4,84 |
47,31 |
|
|
Остаток |
101,4 |
- |
- |
- |
- |
|
|
Итого |
577,04 |
- |
- |
- |
- |
|
|
2 |
Температура воды |
10,25 |
27,01 |
3,28×10-5 |
4,3 |
1,5 |
|
Подекадная сумма осадков |
189,88 |
24,45 |
6,03×10-5 |
4,3 |
27,54 |
|
|
Температура воды и подекадная сумма осадков |
392,45 |
4,05 |
0,07 |
4,84 |
56,92 |
|
|
Остаток |
96,88 |
- |
- |
- |
- |
|
|
Итого |
689,46 |
- |
- |
- |
- |
|
|
3 |
Температура воды |
13,13 |
29,41 |
1,43×10-5 |
4,26 |
2,2 |
|
Подекадная сумма осадков |
195,58 |
25,14 |
4,01×10-5 |
4,26 |
33,01 |
|
|
Температура воды и подекадная сумма осадков |
292,68 |
3,22 |
0,1 |
4,75 |
49,41 |
|
|
Остаток |
91,01 |
- |
- |
- |
- |
|
|
Итого |
592,4 |
- |
- |
- |
- |
Анализируя табл. 1, следует отметить, что температура и количество осадков оказывают влияние на численность личинок во всех исследуемых водоемах (F>>Fкр., р<<0,05), при этом сила влияния количества осадков значительно превышает таковую температурного фактора (27,54-33,01% против 1,5-2,2%). Совместное же влияние указанных факторов на численность личинок в целом за период наблюдений во всех водоемах не выявлено (F<
Очевидно, численность личинок в значительной мере (приблизительно на 30%) определяется количеством осадков. Влияние температурного фактора выражено гораздо слабее по причине его относительной стабильности: как было указано выше, температурные условия исследуемых водоемов относительно благоприятны для развития личинок в течение всего сезона, и варьирование их несущественно (табл. 2).
Таблица 2 . Средние значения и варьирование температуры воды на поверхности исследуемых водоемов
|
№ водоема |
Среднее, ?С |
Коэффициент вариации, % |
Вербальная оценка степени варьирования |
|
1 |
20,7 |
17,3 |
Слабое |
|
2 |
19,3 |
16,6 |
Слабое |
|
3 |
19,4 |
18,6 |
Слабое |
Вышесказанное подтверждается отсут- ствием достоверного совместного влияния температуры и количества осадков: стабильные температурные условия водоемов ни усиливают, ни сглаживают влияние количества осадков. Тем не менее, как показывают наши наблюдения, в отдельные годы совместное влияние температуры и осадков практически полностью определяет численность преимагинальных гемипопуляций. По-види-мому, такая ситуация имеет место в годы с нестабильными температурными условиями и неравномерным распределением осадков. В этом случае совместное влияние указанных факторов на численность преимагинальных гемипопуляций малярийного комара будет выраженным и очень сильным. Можно также предположить, что отсутствие совместного влияния температуры и осадков в большей степени характерно для урбанизированной среды, поскольку ее условия в немалой степени определяют стабильность температурных условий водоемов [5-8].
Направление влияния описываемых факторов на численность личинок представлено на рис. 1-6.
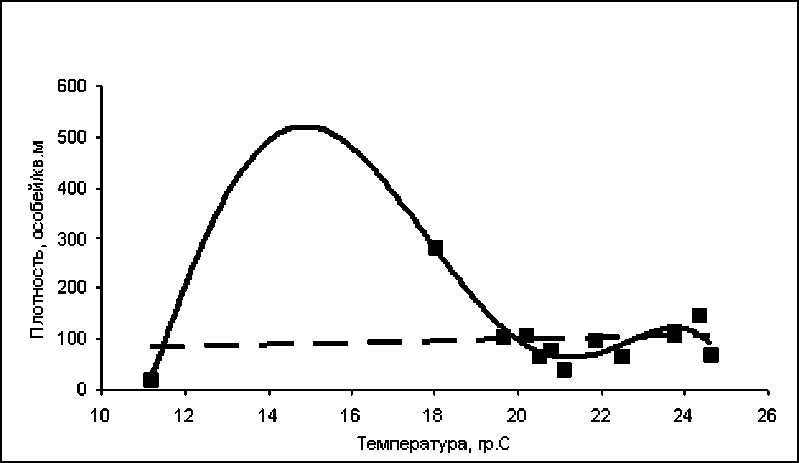
Рис. 1. Зависимость численности личинок от температуры поверхностных слоев воды в водоеме 1
Полиномиальный тренд описывается уравнением у = -0,0422х5+ 3,6228х4- 119,77х3 + 1887х2- 14006х + 38880, R2= 0,9; линейный тренд описывается уравнением у = 1,6533х + 66,623, R2= 0,008.
Для водоема 1 зависимость численности личинок от температуры воды достоверно описывается параболой 5-го порядка (R2=0,9). На рис. 1 видно, что с повышением температуры воды численность личинок резко увеличивается, достигая пика приблизительно при 15°С, а затем столь же резко уменьшается. Следующее увеличение численности личинок, не столь резкое и значительное, начинается при температуре приблизительно 20°С с пиком в области 24°С, после чего численность вновь резко уменьшается. Таким образом, влияние температурных условий на численность личинок в водоеме 1 имеет сложный нелинейный характер (R2 для линейного тренда значительно меньше 0,25). Максимальная численность личинок достигается при относительно низких температурах воды (около 15°С). По-видимому, это обусловлено особенностями водоема: нестабильностью величины площади водного зеркала в течение сезона, значительной гетерогенностью фитоценозов и т.д. Все это приводит к тому, что преимагинальная гемипопуляция достигает максимальной численности во второй половине сезона, ближе к его концу, когда температура воды существенно ниже, чем в июне-июле. Очевидно, что про- гревание воды до температуры выше 20°С в нию численности: об этом свидетельствует середине сезона также приводит к увеличе- второй пик на графике (рис. 1).
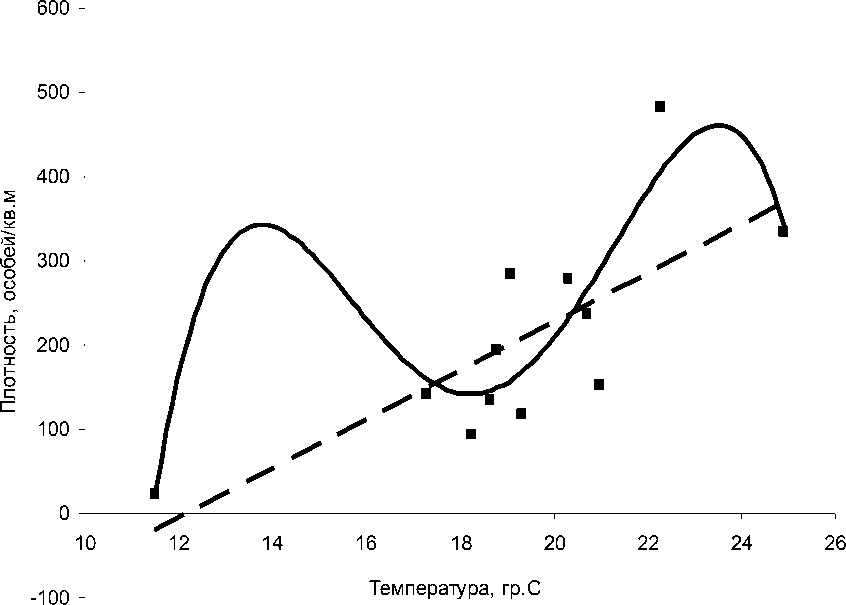
Рис. 2. Зависимость численности личинок от температуры поверхностных слоев воды в водоеме 2
Полиномиальный тренд описывается уравнением у=-0,4575х4+33,857х3-917,84х2+10797х-46247, R2=0,7; линейный тренд описывается уравнением у=29,083х-355,59, R2=0,56.
Для водоема 2 характер влияния температурного фактора схож с таковым для водоема 1 (рис. 1, 2): зависимость достоверно описывается параболой 4-го порядка (R2=0,7) наблюдается два пика плотности при относительно низкой температуре воды (около 13°С) и при достаточно высокой температуре середины сезона (приблизительно 24°С). В отличие от водоема 1, в данном случае второй пик гораздо больше, сопоставим по величине с первым. Очевидно, в водоеме 2 положительное влияние повышения температуры на численность преимагинальной гемипопуляции выражено гораздо четче, чем в водоеме 1. Это подтверждается и возможностью достоверно показать общую тенденцию линейным трендом (R2=0,56): на рис. 2 отражена прямая зависимость численности личинок от температуры воды.
Полиномиальный тренд описывается уравнением у=1,006х3-53,903х2+949,37х-5366,5, R2=0,56; линейный тренд описывается уравнением у=13,429х-129,94, R2=0,43.
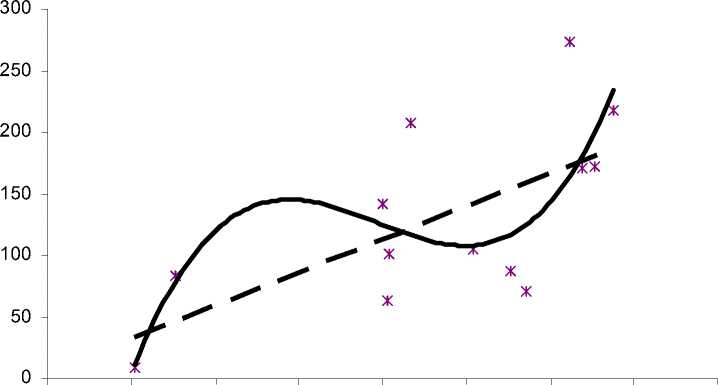
Для водоема 3 зависимость численности личинок от температуры воды достоверно описывается параболой третьего порядка (R2=0,56). На рис. 3 видно, что 2 пика численности личинок в водоеме 3 выражены гораздо слабее, чем в двух других (рис. 1-3): после пика приблизительно при 15°С наблюдается незначительное снижение численности, близкое к выходу на плато, с последующим увеличением и пиком приблизительно при 24°С. Также зависимость можно досто-
10 12 14 16 18 20 22 24 26
Температура, гр.С
Рис. 3. Зависимость численности личинок от температуры поверхностных слоев воды в водоеме 3
верно описать линейным трендом (R2=0,43): с увеличением температуры воды численность личинок увеличивается. Таким образом, характер влияния температуры на численность преимагинальной гемипопуляции малярийного комара в водоеме 3 несколько отличается от такового в водоемах 1 и 2, при этом между последними прослеживается определенное сходство (рис. 1, 2). Анализируя полученные результаты, можно предположить, что в водоемах 1 и 2, испытывающих более интенсивное антропогенное воздействие по сравнению с водоемом 3, особенности влияния температурного фактора на численность личинок во многом определяются именно этим воздействием. Вероятно, характер влияния температуры в водоеме 3 близок к таковому для индигенных местообитаний, а в условиях урбанизированной среды обитания будет наблюдаться тенденция, выявленная для водоемов 1 и 2.
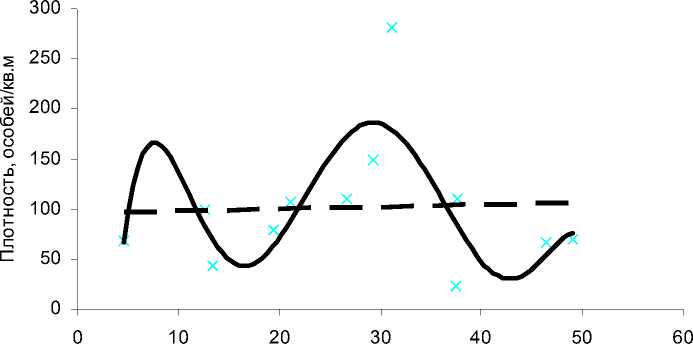
Осадки, мм
Рис. 4. Зависимость численности личинок от количества осадков в водоеме 1 в сезонах 2005-2007 г.г. и в среднем за период наблюдений
Полиномиальный тренд описывается уравнением у=-9Ч10-6х6+0,0016х5-0,105х4+3,4441х3-56,225х2+418,96х-969,29, R2=0,54; линейный тренд описывается уравнением у=0,2287х+94,542, R2=0,002.
Для водоема 1 зависимость численности личинок от количества осадков достоверно описывается параболой 6-го порядка (R2=0,54) и имеет сложный нелинейный характер (R2 для линейного тренда значительно меньше 0,25). Максимальная численность личинок наблюдается при достаточно малой декадной сумме осадков – менее 10 мм (рис. 4). По-видимому, такую картину дает подъем численности личинок в периоды сезона, характеризующиеся высокими температурами воды и наименьшим количеством осадков. Обычно это конец июня-июль, а в конкретном случае сказывается еще и аномально теплый и сухой май 2005 г. Второй пик численности соответствует декадной сумме осадков приблизительно 40 мм и отражает подъем численности личинок во II половине сезона, характерный для водоема 1. С увеличением количества осадков в этот период происходит увеличение площади водного зеркала, которое, как указывалось выше, благоприятно сказывается на численности преимаги-нальной гемипопуляции водоема 1. Падение численности личинок при декадной сумме осадков более 40 мм объясняется неблагоприятным влиянием избыточного их количества: одинаково неблагоприятно как единовременное выпадение большого количества осадков (ливни), так и более или менее равномерное их распределение в течение декады. Картина снижения численности преима-гинальной гемипопуляции при декадной сумме осадков порядка 10-20 мм, вероятно, объясняется вышеуказанными особенностями водоема (нестабильностью величины площади водного зеркала в течение сезона, значительной гетерогенностью фитоценозов и т.д.) Таким образом, влияние количества осадков на численность личинок в водоеме 1 имеет сложный нелинейный характер (R2 для линейного тренда значительно меньше 0,25).
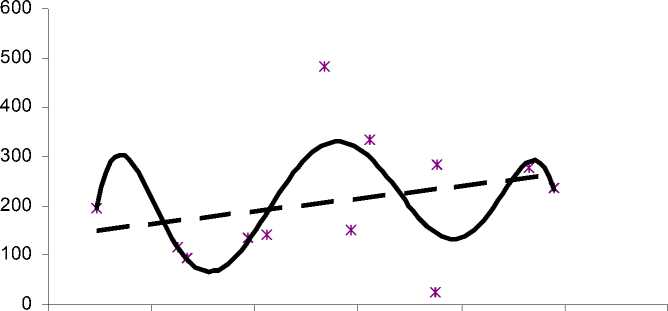
0 10 20 30 40 50 60
Осадки, мм
Рис. 5. Зависимость численности личинок от количества осадков в водоеме 2
Полиномиальный тренд описывается уравнением у=-2Ч10-5х6+0,0036х5-0,288х4+7,0109х3-106,91х2+737,19х-1532,1, R2=0,46; линейный тренд описывается уравнением у=2,5372х+137,23, R2=0,08.
Для водоема 2 зависимость численности личинок от количества осадков достоверно описывается параболой 6-го порядка (R2=0,46) и имеет сложный нелинейный характер (R2 для линейного тренда значительно меньше 0,25). В течение сезона 3 пика численности личинок соответствуют приблизительно 5, 20-30 и 50 мм декадной суммы осадков (рис. 5). Причины первого пика на линии тренда аналогичны таковым для водо- ема 1: такая картина отражает увеличение плотности личинок в середине сезона, когда температура воды высока, а количество осадков минимально. С увеличением количества осадков до 10 мм численность личинок плавно возрастает и, начиная приблизительно с 30 мм, плавно снижается. Эти результаты соответствуют приводимым в литературе данным и отражают неблагоприятное влия- ние как недостатка, так и избытка осадков [1, 9, 10]. Резкий рост численности личинок в водоеме 2 отмечается также при декадной сумме осадков около 50 мм (рис. 5). По-ви-димому, такую картину формирует увеличение площади водного зеркала (и анофелоген-ной площади) после обмеления водоема в середине сезона.
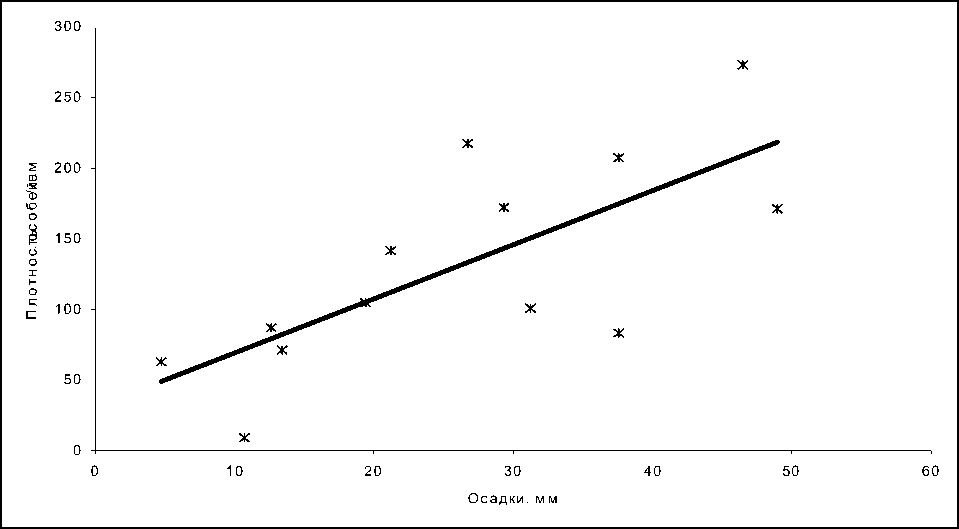
Рис. 6. Зависимость численности личинок от количества осадков в водоеме 3
Линейный тренд описывается уравнением у=3,8336х+30,765, R2=0,52.
Зависимость численности личинок от количества осадков в водоеме 3 достоверно описывается уравнением линейной функции (R2=0,52). С увеличением декадного количества осадков от 10 до 50 мм численность личинок возрастает.
Таким образом, превалирующим фактором, определяющим численность личинок в исследуемых водоемах, является количество осадков, сила влияния этого фактора составляет приблизительно 30% от суммарного влияния всех факторов. Влияние температуры и осадков на численность личинок малярийного комара носит преимущественно сложный нелинейный характер, хотя можно сказать, что в целом повышение температуры и увеличение количества осадков благоприятно сказывается на численности личинок. Выявлено два диапазона температуры воды, благоприятных для увеличения численности: первый составляет приблизительно 12-15єС, второй – 20-24єС. По-видимому, такая картина характерна для урбанизированной среды обитания и обусловлена особенностями водоемов, испытывающих значительный антропогенный прессинг. Возможно, это также свидетельствует о существовании двух или более морф, адаптированных к различным температурам воды.
По-видимому, оптимальная для увеличения численности личинок декадная сумма осадков колеблется от 20 до 50 мм. При этом важную роль играют величина площади водного зеркала (и анофелогенной зоны), существующая на момент выпадения осадков, а также интенсивность и качественная характеристика осадков. Если осадки способству- ют увеличению площади анофелогенной зоны, то это благоприятно сказывается на численности личинок. Благоприятно сказывается и равномерное выпадение осадков низкой интенсивности в течение определенного периода. Ливневые осадки и град будут способствовать сокращению численности преимагинальной популяции.
Выводы
-
1. Превалирующим фактором, определяющим численность личинок в водоемах урбани- зированной среды обитания, является количество осадков.
Список литературы Влияние температуры воды и количества осадков на численность преимагинальной гемипопуляции малярийного комара (комплекс видов Anopheles maculipennis Meigen, 1818) в условиях урбанизированной среды обитания
- Малярийные комары и борьба с ними на территории Российской Федерации: Методические указания. М.: Федеральный центр госсанэпиднадзора Минздрава России, 2000.
- Заречная С.Н., Маханько Е.В. Видовая диагностика кровососущих комаров и синантропных мух: Учебн. пособие. М.: Изд-во ЦИУВ, 1992.
- Лакин Г.Ф. Биометрия: Уч. пособие для ун-тов и пед. ин-тов. М.: Высшая школа, 1973.
- Боровиков В. Statistica: искусство анализа данных на компьютере. Для профессионалов. СПб.: Питер, 2001.
- Алексеев С.В., Пивоваров Ю.П. Экология человека. М.: Изд. ГОУ ВУНМЦ МЗ РФ, 2001.
- Новиков Ю.В., Куценко Г.И., Подольский В.М. Современные эколого-гигиенические проблемы среды обитания человека и совершенствование санитарно-эпидемиологического надзора. М.: Изд. ТОО «Рагоръ», 1997.
- Пивоваров Ю.П. Гигиена и экология человека (курс лекций). М.: Изд. РГМУ, 1999.
- Заречная С.Н. Особенности биологии комаров -переносчиков малярии в СССР: Лекция. М.: Изд. РИО ЦОЛИУВ, 1982. 25 с.
- Ануфриева В.Н. Переносчики малярии в Российской Федерации: Материалы совещания «Современные проблемы эпиднадзора за малярией». М.: Изд. отд. ФЦГСЭН Минздрава России, 2001.
- Иванова Т.Н. Природные и социальные условия, способствующие восстановлению местной передачи трехдневной малярии в Московском регионе/Здоровье населения и среда обитания: информационный бюллетень. №10/151/. М.: Изд. ФГУЗ «ФЦГиЭ», 2005.
