Влияние теплового закаливания на углеводный обмен проростков пшеницы и фасоли при адаптации к гипертермии

Автор: Чудинова Лариса Алексеевна
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Экология
Статья в выпуске: 2, 2019 года.
Бесплатный доступ
Изучено содержание основных углеводов (глюкозы и сахарозы) и активность ключевых ферментов углеводного обмена (амилазы и инвертазы) в процессе адаптации проростков пшеницы и фасоли к гипертермии (+40°С, 3 сут.) в условиях наличия или отсутствия теплового закаливания (+37°С, 3 ч.). Установили, что предварительное тепловое закаливание приводит к значительному росту количества глюкозы и сахарозы у обоих растений в условиях последующей гипертермии. При этом у пшеницы амилазная и инвертазная активности были понижены, тогда как у фасоли на фоне снижения активности амилазы, активность инвертазы значительно возрастала. Это позволило предположить, что в основе теплового закаливания обеих культур лежит повышенный уровень растворимых сахаров, который достигается у них разными механизмами: у пшеницы - за счет роста интенсивности фотосинтеза, а у фасоли - за счет торможения оттока сахарозы во флоэму.
Пшеница, фасоль, гипертермия, тепловое закаливание, глюкоза, сахароза, амилаза, инвертаза
Короткий адрес: https://sciup.org/147227083
IDR: 147227083 | УДК: 581.1:632.122.1 | DOI: 10.17072/1994-9952-2019-2-206-211
Influence of thermal hardening on the carbohydrate metabolism of wheat and beans seedlings in adaptation to hyperthermia
The content of basic carbohydrates (glucose and sucrose) and the activity of the key enzymes of carbohydrate metabolism (amylase and invertase) in the process of wheat and bean seedlings adaptation to hyperthermia (+ 40°C 3 days) in the presence or absence of heat hardening (+ 37°C, 3 h) were studied. It was established that preliminary thermal hardening leads to a significant increase in the amount of glucose and sucrose in both plants under conditions of subsequent hyperthermia. At the same time, in wheat, amylase and invertase activities were reduced, whereas in beans, amylase decreased, invertase activity increased significantly. This suggests that the heat hardening of both cultures is based on an increased level of soluble sugars, which is achieved by different mechanisms: in wheat, due to an increase in the intensity of photosynthesis, and in beans, due to inhibition of sucrose outflow to the phloem.
Текст научной статьи Влияние теплового закаливания на углеводный обмен проростков пшеницы и фасоли при адаптации к гипертермии
Высокотемпературный стрессор представляет собой один из самых значимых абиотических факторов, оказывающих существенное влияние на интенсивность и направленность физиологических и биохимических процессов, рост и продуктивность растений.
Важная роль углеводного обмена для процессов адаптации растений к температурным воздействиям в настоящее время не вызывает сомнения. Однако направленность превращений углеводноферментного комплекса в условиях стресса, в том числе и температурного, до настоящего времени остается предметом дискуссии. Одни авторы счи- тают, что это результат деструктивных процессов, связанных с повреждением клеток в условиях стресса, другие – одной из составляющих комплекса защитных реакций на стресс [Колупаев, Трунова, 1992; Титов, Таланова, 2009]. Следует отметить, что наибольшее количество экспериментальных данных по углеводному обмену получено при изучении гипотермии. В то же время исследованию роли данного обмена в формировании теплоустойчивости уделялось значительно меньше внимания. Особенно мало работ по физиологобиохимическим основам теплового закаливания, по результатам которых известно, что тепловое закаливание представляет собой сложный многокомпонентный кооперативный процесс, в который
вовлечены как специфические, так и неспецифические изменения [Шакирова, 2001]. Поскольку многие авторы считают, что усиление гидролиза полимерных форм углеводов, в том числе и низкомолекулярных олигосахаридов, является неспецифической ответной реакцией растений на неблагоприятные воздействия различной природы (холодовой, тепловой, осмотический, солевой и др.) [Карпец, Колупаев, 2009; Чудинова, Суворов, 2011], то можно предполагать участие углевод-ферментного комплекса в развитии теплоустойчивости при закаливании. В этом плане большой интерес представляют гидролитические ферменты, играющие важную роль в углеводном метаболизме, в частности, амилаза и инвертаза. Однако на сегодняшний день отсутствуют четкие экспериментальные доказательства связи активности данных ферментов с формированием устойчивости растений к гипертермии после закаливания.
В связи с вышеизложенным, цель нашей работы – изучение влияния теплового закаливания на содержание растворимых углеводов и активность ключевых гидролитических ферментов углеводного обмена в проростках пшеницы и фасоли в процессе их адаптации к последующей гипертермии.
Объекты и методы исследований
Проростки пшеницы мягкой ( Triticum аestivum L.), сорт ‘Иргина’, и фасоли спаржевой ( Phaseolus nanus L.), сорт ‘Журавушка’, выращивали 10 сут. в климатической камере ШН-М на смеси вермикулита и песка в соотношении 1:1 при температуре 22°C, влажности 80–85% и освещенности люминесцентными лампами 18 ч. в сут. Затем эксперимент в течение 3 сут. проводили по следующей схеме:
-
1 вариант – экспозиция проростков при +22°C (контроль);
-
2 вариант – экспозиция проростков при +40°С (гипертермия);
-
3 вариант – экспозиция при +40°С с предобработкой +37°С 3 ч. (тепловое закаливание).
Для анализа растения брали на 1-е и 3-и сут. адаптации. Побеги фиксировали паром в течение 5 мин. и досушивали до воздушно-сухого состояния. Определяли содержание глюкозы и сахарозы фотометрическим методом [Большой практикум …, 2012]. Суммарную активность амилаз определяли по степени гидролиза крахмала, активность инвертазы – по накопленной в пробе глюкозы.
Повторность определений – 3-кратная. Результаты обрабатывали статистически с использованием однофакторного дисперсионного анализа. Определяли достоверность различий между некоторыми вариантами по наименьшей существенной разнице (НСР 0,05 ).
Результаты и их обсуждение
Полученные нами результаты (рис.1, А, Б) показали, что исходный статус глюкозы (контроль) у проростков пшеницы был намного выше, чем у проростков фасоли. Это согласуется с мнением ряда авторов о том, что злаки в целом являются растениями с углеводным типом обмена веществ, тогда как бобовые – с белковым типом [Строгонов, 1962].
Содержание глюкозы в проростках пшеницы при гипертермии на 1-е сут. экспозиции сохранялось на уровне контроля, тогда как на 3-и сут. – незначительно снижалось. Тепловое закаливание достоверно стимулировало накопление глюкозы на 3-и сут. адаптации проростков пшеницы к гипертермии (рис. 1, А).
Иная тенденция прослеживалась в проростках фасоли (рис. 1, Б). Так, во всех опытных вариантах при 1-суточной экспозиции наблюдались незначительные колебания содержания глюкозы в сравнении с контролем, тогда как на 3-и сут. адаптации к гипертермии отмечалось достоверное накопление глюкозы, которое сохранялось и в варианте с предварительным закаливанием.
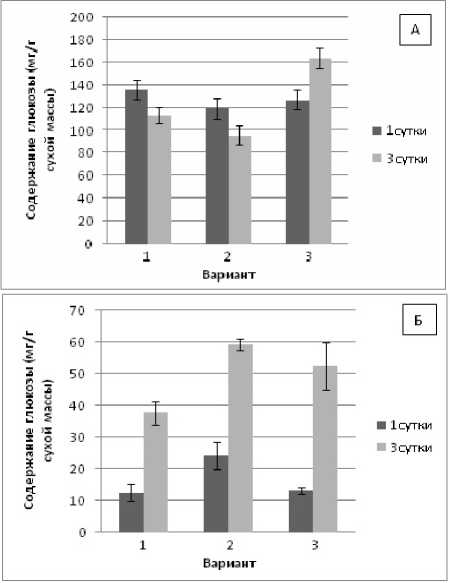
Рис. 1 . Изменение содержания глюкозы в проростках пшеницы (А) и фасоли (Б) при гипертермии без теплового закаливания и после него:
Здесь и далее: 1 вариант – контроль; 2 вариант – гипертермия (+40 ° C); 3 вариант - тепловое закаливание (+37 ° C, 3 ч) и последующая гипертермия (+40 ° C)
Таким образом, в целом, для пшеницы и фасоли характерен довольно стабильный или повышенный уровень глюкозы при гипертермии на 3-и сут. адаптации без закаливания и, особенно, с предварительным тепловым закаливанием. По мнению рядя авторов, повышенное содержание глюкозы можно считать защитно-приспособительной реакцией на стрессор [Дерябин и др., 2007]. Аналогичное предположение было выдвинуто и в отношении сахарозы, поскольку зафиксировано увеличение ее содержания и изменение изоферментного спектра в ответ на стрессы самой различной природы – гипертермию, засоление, обезвоживание, раневые воздействия, заражение патогенном [Колупаев, Трунова, 1992].
Мы наблюдали снижение содержания сахарозы в проростках пшеницы на 1-е и 3-и сут. адаптации при воздействии гипертермии (рис. 2, А) и заметное увеличение в условиях предварительного теплового закаливании (на 35% по отношению к контролю и в 2.8 раза по сравнению с вариантом без закаливания).
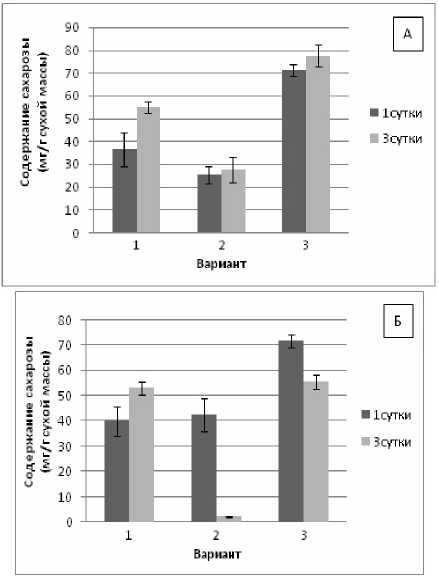
Рис. 2. Изменение содержания сахарозы в проростках пшеницы (А) и фасоли (Б) при гипертермии без теплового закаливания и после него
В проростках фасоли при 1-суточном воздействии гипертермии количество сахарозы оставалось на уровне контроля, а через 3-е сут. экспозиции – резко снижалось по сравнению с контролем (рис. 2, Б). Предварительное тепловое закаливание стимулировало накопление сахарозы у фасоли, что говорит о включении механизмов адаптации.
Механизмы защитного действия растворимых углеводов в растениях при действии стрессоров могут быть достаточно разнообразны. Так, показано их активное участие в «тушении» окислитель- ного стресса, вызванного гипертермией, антиденатурирующее действие на белково-липидный комплекс мембран, стабилизация белковых компонентов клеток, подвергнутых нагреву. Наконец, сахара могут неспецифически ингибировать метаболизм вследствие адсорбции на поверхности макромолекул ферментов и таким образом препятствовать конформационным изменениям, необходимым для осуществления катализа [Колупаев, Трунова, 1992; Кафи, Стюарт, Борланд, 2003; Карпец, Колупаев, 2009].
Содержание растворимых сахаров в значительной степени определяется активностью гидролитических ферментов – инвертазы и амилазы, которая изменяется в условиях воздействия стрессоров различной природы [Трунова, 2007].
Инвертаза, катализирующая реакцию расщепления сахарозы на глюкозу и фруктозу, обеспечивает гексозами энергетические процессы, вовлечена в разгрузку флоэмы, играет ключевую роль в процессах роста и развития [Даффус, Даффус, 1987].
Полученные нами результаты показали (рис. 3, А), что активность инвертазы контрольных растений пшеницы снижалась на 3-и сут. в 2.3 раза, что свидетельствует о загрузке флоэмы сахарозой. В условиях воздействия гипертермии без теплового закаливания и, особенно, с предварительным закаливанием активность инвертазы пшеницы была достоверно ниже контроля. Поскольку в надземной части проростков пшеницы одновременно с пониженной активностью инвертазы наблюдалось и снижение содержания сахарозы, то можно говорить о преобладании транспорта сахарозы по флоэме в корневую часть растения в условиях действия гипертермии.
В проростках фасоли (рис. 3, Б) активность инвертазы во всех вариантах была на низком уровне. Исключение – в варианте с предварительным закаливанием, где активность инвертазы резко возрастала на 3-и сут. адаптации к гипертермии. Можно предположить, что в данном случае происходит интенсивный гидролиз сахарозы, который служит механизмом накопления глюкозы.
Данные, имеющиеся в литературе, об изменении активности инвертазы после температурного воздействия – противоречивы. Так, одни авторы отмечают усиление активности этого фермента [Дерябин и др., 2007], а другие – снижение [Кси-шев, 1992]. Одной из причин расхождения результатов о характере ответной реакции инвертазы на стрессы может быть различное поведение фермента в зависимости от напряженности и продолжительности стресса. Известно, что изменение активности инвертазы при воздействии стрессора может происходить за счет изменения количества фермента в ткани, преобразования изоферментного состава, регуляции каталитической активности уже существующих молекул фермента, изменения активности фермента вследствие взаимного перехода его растворимой и связанной с клеточными стенками форм [Сабельникова, 2007].
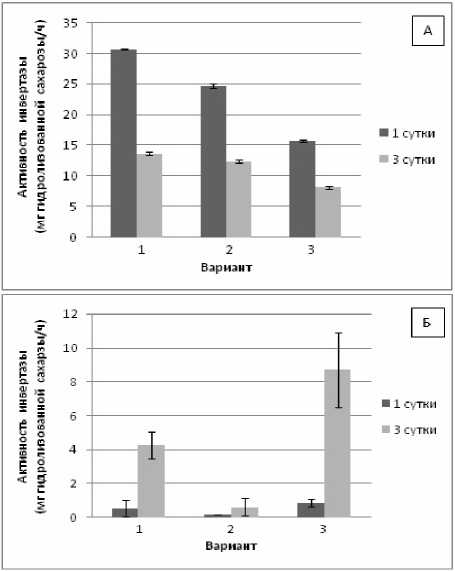
Рис. 3 . Изменение активности инвертазы в проростках пшеницы (А) и фасоли (Б) при гипертермии без теплового закаливания и после него
В наших исследованиях низкая активность инвертазы и следовые количества сахарозы у растений фасоли при гипертермии указывают на процессы загрузки флоэмы и транспорта сахарозы в корни. Апопластный способ загрузки флоэмы и подавление активности кислой инвертазы в растениях делает возможным отток сахаров из листьев, что позволяет обеспечивать сахарами корневую систему. Показано [Попов, 2002], что процессы оттока сахаров из листьев в корни играют значительную роль в адаптации теплолюбивых растений к низким положительным температурам.
Как известно, амилолитическая активность определяет уровень пластидного крахмала, а также крахмала амилопластов, выполняющих роль своеобразного депо глюкозы [Даффус, Даффус, 1987]. Нами установлено достоверное увеличение активности амилазы в проростках пшеницы на 3-и сут. действия повреждающей температуры по отношению к контролю (рис. 4, А). При предварительном закаливании активность амилазы была существенно пониженной в оба срока определения в сравнении с контролем, а также с вариантом – гипертермия без закаливания.
В проростках фасоли было отмечено снижение активности амилазы в побегах опытных вариантах на 3-и сут. экспозиции по сравнению с контролем
(рис.4, Б). Можно предположить, что благодаря закаливанию, в проростках фасоли происходит восстановление фотосинтетических процессов.
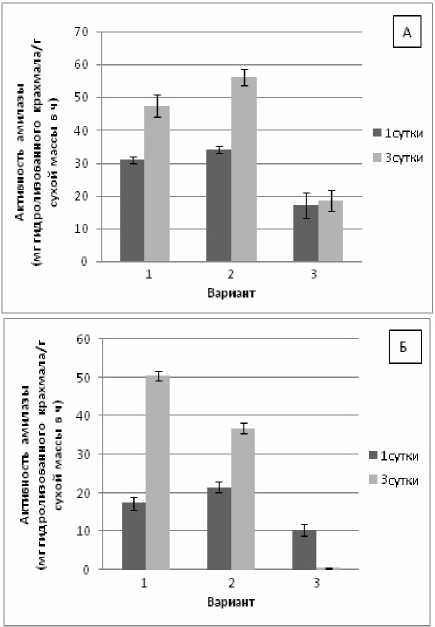
Рис. 4. Изменение активности амилазы в проростках пшеницы (А) и фасоли (Б) при гипертермии без теплового закаливания и после него
На основании полученных нами результатов можно сделать следующие основные выводы:
Адаптация к гипертермии проростков пшеницы и фасоли сопровождается ростом уровня глюкозы, что происходит, предположительно, за счет повышения интенсивности фотосинтеза, а не за счет амилолиза. На это указывает снижение активности амилазы в этих условиях. Выявленный при этом низкий уровень сахарозы и активности инвертазы указывает на активную загрузку флоэмы и отток сахаров из листьев, что позволяет обеспечивать сахарами корневую систему в условиях температурного стресса.
Предварительное тепловое закаливание приводит к значительному росту количества глюкозы и сахарозы у обоих растений в условиях последующей гипертермии. При этом амилазная активность оставалась ингибированной, тогда как инвертазная активность была низкой у пшеницы, у фасоли она значительно возрастала. Это позволяет предположить, что в основе теплового закаливания обеих культур лежит повышенный уровень растворимых сахаров, который достигается у них разными механизмами: у пшеницы – за счет повышенной ин- тенсивности фотосинтеза, а у фасоли – за счет торможения оттока сахарозы во флоэму.
Список литературы Влияние теплового закаливания на углеводный обмен проростков пшеницы и фасоли при адаптации к гипертермии
- Большой практикум «Биохимия»: лабораторные работы / сост. М.Г. Кусакина, В.И. Суворов, Л.А. Чудинова. Пермь: Изд-во ПГНИУ, 2012. С. 26-35.
- Даффус К., Даффус Дж. Углеводный обмен растений. М.: Агропромиздат, 1987. 150 с.
- Дерябин А.Н. и др. Влияние сахаров на развитие окислительного стресса, вызванного гипотермией (на примере растений картофеля, экспрессирующих ген инвертазы дрожжей) // Физиология растений. 2007. Т. 54, № 1. С. 39-46.
- Карпец Ю.В., Колупаев Ю.Е. Ответ растений на гипертермию: молекулярно-клеточные аспекты // Вестник Харьковского национального аграрного университета. Сер. Биология. 2009. Вып.1 (16). С. 16-38.
- Кафи М., Стюарт В.С., Борланд А.М. Содержание углеводов и пролина в листьях, корнях и апексах пшеницы, устойчивых и чувствительных к засолению // Физиология растений. 2003. Т. 50, № 2. С. 174-182.
- Колупаев Ю.Е., Трунова Т.И. Особенности метаболизма и защитные функции углеводов растений в условиях стрессов // Физиология и биохимия культ. растений. 1992. Т. 24, № 6. С. 523-533.
- Ксишев Ю.Е. Стресс-индуцируемое изменение активности инвертазы и превращение растворимых углеводов в злаковых растениях: возможные причины и физиологическая роль: автореф. дис. … канд. биол. наук. М., 1992. 20 с.
- Попов В.Н. Сравнительное исследование низко-температурной адаптации томата и огурца в связи с их углеводным метаболизмом: дис. … канд. биол. наук. М., 2002. 115 с.
- Сабельникова Е.П. Роль углеводного метаболизма в устойчивости растений к гипотермии на примере картофеля, трансформированного геном дрожжевой инвертазы: дис.... канд. биол. наук. М., 2007. 137 с.
- Строгонов Б.П. Физиологические основы солеустойчивости растений. М.: Наука, 1962. 260 с.
- Титов А.Ф., Таланова В.В. Устойчивость растений и фитогормоны. Петрозаводск, 2009. 206 с.
- Трунова Т.И. Растение и низкотемпературный стресс. М.: Наука, 2007. 54 с.
- Чудинова Л.А., Суворов В.И. Роль некоторых низкомолекулярных соединений в механизме перекрестной адаптации растений // Вестник Пермского ун-та. Сер. Биология. 2011. Вып. 1. С. 17-20.
- Шакирова Ф.М. Неспецифическая устойчивость растений к стрессовым факторам и ее регуляция. Уфа: Гилем, 2001. 160 с.
