Влияние типа лесорастительных условий на поглотительную деятельность корневой системы и биологическую продуктивность лиственницы сибирской на уровне организма в онтогенезе

Автор: Лебедев Е.В.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Экология
Статья в выпуске: 1, 2013 года.
Бесплатный доступ
Получены количественные данные чистой продуктивности фотосинтеза, минеральной и биологической продуктивности и характер связи между ними у лиственницы сибирской разных типов леса юга Красноярского края.
Лиственница сибирская, чистая продуктивность фотосинтеза, минеральное питание, биологическая продуктивность, типы леса, онтогенез, красноярский край
Короткий адрес: https://sciup.org/14082693
IDR: 14082693 | УДК: 581.43:631.811:630*161.32*187:674.032.475.352
The forest vegetation condition type influence on the root system absorbing activity and on Siberian larch biological productivity at the organism level in ontogenesis
The quantitative data of the photosynthesis net productivity, the mineral and biological productivity, the nature of the relationship between them in the Siberian larch in the Krasnoyarsk Krai South different forest types are received.
Текст научной статьи Влияние типа лесорастительных условий на поглотительную деятельность корневой системы и биологическую продуктивность лиственницы сибирской на уровне организма в онтогенезе
Введение. Успешное управление ростовыми процессами растений невозможно без учета количественных данных фотосинтетической активности, минеральной и биологической продуктивности. Подобные комплексные исследования весьма редки и разрознены [7], поскольку изучение продуктивности изначально строилось на измерениях хозяйственно ценной части ствола [15]. Фотосинтез хвойных изучался газометрическими методами, либо на основе анализа флюоресценции хвои [12], что не давало количественных данных прироста органического вещества за длительный период (вегетацию) из-за весьма сложного учета потерь на дыхание и корневые экссудаты [3]. Минеральное питание изучалось чаще всего на декапитирован-ных корнях и ограничивалось определением адаптивной реакции корневой системы на условия произрастания [10]. Указанные методы не позволяют перейти на уровень целого организма и связать поглощение элементов питания с фотосинтезом и продуктивностью растения. Между тем табличные данные по фитомассе лесов Северной Евразии, представленные В.А. Усольцевым [13] на основе рекурсивно-блочного моделирования обширного материала таблиц хода роста древостоев на уровне организма в различных условиях, позволяют существенно расширить знания о биологии древесных растений в онтогенезе, привлекая сведения, полученные в модельных микрополевых опытах с изучаемыми растениями [7, 9] и природно-климатические данные мест произрастания [4, 6].
Целью исследования было проведение по табличным материалам [13] комплексного физиологического анализа на уровне организма и получение расчетных количественных данных чистой продуктивности фотосинтеза, минеральной и биологической продуктивности, депонирования углерода и характера связи между ними у растений лиственницы сибирской в различных типах лесорастительных условий юга Красноярского края.
Объекты и методы. Физиологическому анализу подвергнуты табличные данные [13, с. 137 – 139 ] модельных лиственничников ( Larix sibirica Ledeb.) южных районов Красноярского края, произраставших в зоне лесостепи Алтае-Саянской горной провинции, составленные по материалам [14]. Типы лесорастительных условий: широкотравный (бонитет I), разнотравный (бонитет II), мшисто-ягодный (бонитет III) и остепненный (бонитет IV). Возрастной период от 30 до 220 лет с интервалом 10 лет. Безморозный период 120 дней. Годовое количество осадков 470 мм. Климат резко континентальный.
Таксационные данные масс корней, листьев, древесины стволов и сучьев пересчитывали на одно растение по возрастам. Поверхность хвои определяли по вычисленным нами коэффициентам на свежем материале [1]. На 1 г сухой хвои приходилось 140 см 2 поверхности. Чистую продуктивность фотосинтеза (ЧПФ) определяли за каждый сравниваемый период в г/м 2 день [9]. Депонирование углерода в расчете на 1 га определяли по [2]. Для расчета активной поверхности корней всего растения применяли данные наших модельных микрополевых опытов с 1–3-летними растениями на дерново-подзолистой и серой лесной почвах [8]. В силу высокого постоянства морфологии активных корней в пределах растения (диаметра, длины активного корня, величины удельной активной поверхности корневой системы (УАПКС) и длины корней, приходящихся на единицу массы корневой пряди диаметром 2–3 мм), для расчета активной поверхности корней растения применяли средние значения УАПКС и длины активных корней, приходящихся на единицу массы пряди: 3,8 см 2 /м и 24 м/г соответственно [7]. На 1 г сухой массы пряди приходилось 91,2 см 2 активной поверхности корней. Листовой аппарат и активная часть корневой системы – две стороны единого процесса питания, и между ними существует тесная функциональная связь. Отношение корневого потенциала (КП) к фотосинтетическому (ФП) в наших опытах [7] было в среднем равно 0,24. В функциональном отношении это означает, что 1 м 2 активной поверхности корней обслуживал 4,17 м 2 хвои. Используя средние значения отношения поверхности активных корней к поверхности хвои, полученные в модельных опытах, определяли поверхность активных корней всего растения и долю их в массе корней в каждом возрасте. По полученным данным активная часть корневой системы не превышала 3% от массы корней растения, которая использовалась в дальнейших расчетах. Вычисленные размеры КП в каждом возрасте позволяют определить минеральную продуктивность корней целого растения [8]. С растений разного возраста отбирали пробы хвои, ветвей, стволов и корней разного диаметра, группировали по органам и определяли в них содержание N, P, К, Са и Мg по [11]. Содержание элементов в единице массы дерева в каждом сравниваемом периоде определено с учетом соотношения между органами. Потребность в элементах минерального питания при формировании единицы биомассы у различных видов растений довольно стабильна и контролируется генетически, несмотря на варьирование условий их произрастания. Это связано со спецификой обменных процессов, которые целенаправленны и требуют строго нормированного количества элементов [5]. Поэтому полученные нами данные использованы в расчетах. Биологическую продуктивность (БП) находили по относительному увеличению исходной массы растения в сравниваемых периодах. Полученные данные подвергали корреляционному и регрессионному анализам.
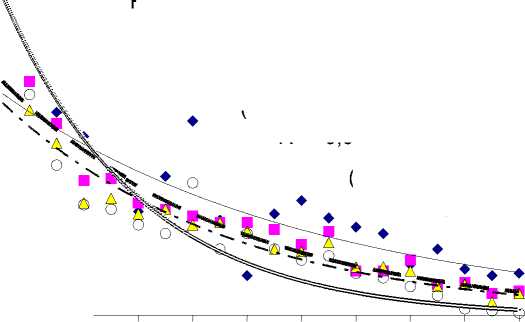
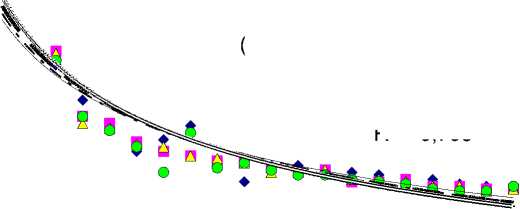
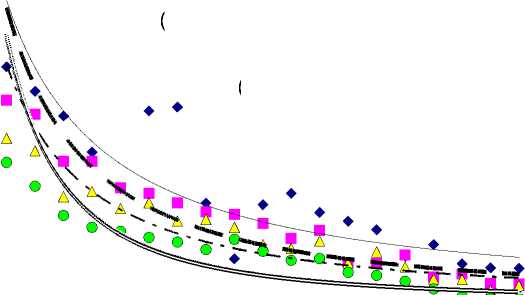
Результаты исследования. За период от 30 до 220 лет чистая продуктивность фотосинтеза (ЧПФ) снизилась в 5,9, 9,6, 9,3 и 100,0 раз соответственно у растений бонитетов I–IV (рис. 1,а). Связь ЧПФ с возрастом была высокая отрицательная ( r = от -0,855 до -0,936). Биологическая продуктивность (БП) падала с возрастом во всех типах леса (рис. 1,б) в 1,8–2,2 раза ( r = от -0,709 до -0,795). Количество углерода, депонированного фотосинтетическим аппаратом в расчете на 1 га за период онтогенеза, уменьшалось в 7,2, 11,8, 10,7 и 62,0 раза соответственно от I к IV бонитетам (рис. 1,в). Связь с возрастом была высокая обратная ( r = от -0,888 до -0,963). Минеральная продуктивность (МП) во всех типах леса по всем минеральным элементам падала с возрастом: по N – в 21,0–335,0, по P – в 27,3–330,0, по K – в 17,7–490,0, по Ca – в 20,9– 362,0, а по Mg – в 19,4–425,0 раза соответственно у растений бонитетов I–IV (рис. 2). Максимальные различия МП в онтогенезе были в остепненном типе (IV бонитет), а минимальные – в широкотравном (I бонитет). Максимальное количество каждого элементов поглощалось в широкотравном типе лесорастительных условий, а минимальное – в остепненном.
1,6
J)1,4
s1,2
2s
1,0
m
2 0,8
о0,6 о
A 0,4
0,2
0,0
(4) y = 2,641e-0,020x R2 = 0,809
(1) y = 1,303e-0,0087x R2 = 0,727
-
(2) y = 1,504e-0,012x
R2 = 0,940
-
(3) y = 1,392e-0,012x R2 = 0,910
2,2
2,0
1,8
К Я * о ф
о о S ш
£ 1,6 о г 1,4
Ё 1,2
Q. 1,0
0,8
(1) y = 4,6605x-0,2888 R2 = 0,848
(2) y = 5,2847x-0,3163
R2 = 0,846
-
(3) y = 5,0103x-0,3058
R2 = 0,847
-
(4) y = 5,7752x-0,3356
R2 = 0,768
Экология
20 40 60 80 100 120 140 160 180 200 220 240
20 40 60 80 100 120 140 160 180 200 220 240
Возраст, лет
Возраст, лет
-
(1) y = 439,49x-1,1329 R2 = 0,758
(4) y = 8899,3x-1,9829 R2 = 0,692
-
(2) y = 1173,2x-1,4057
R2 = 0,868
-
(3) y = 822,27x-1,3683 R2 = 0,812
-
♦ широкотравный (б I)
-
■ разнотравный (б II)
А мшисто-ягодный (б III)
О остепнённый (б IV)
20 40 60 80 100 120 140 160 180 200 220 240
Возраст, лет
Рис. 1. Чистая продуктивность фотосинтеза, биологическая продуктивность 7 и 2 депонирование углерода у лиственницы в разных типах леса в онтогенезе
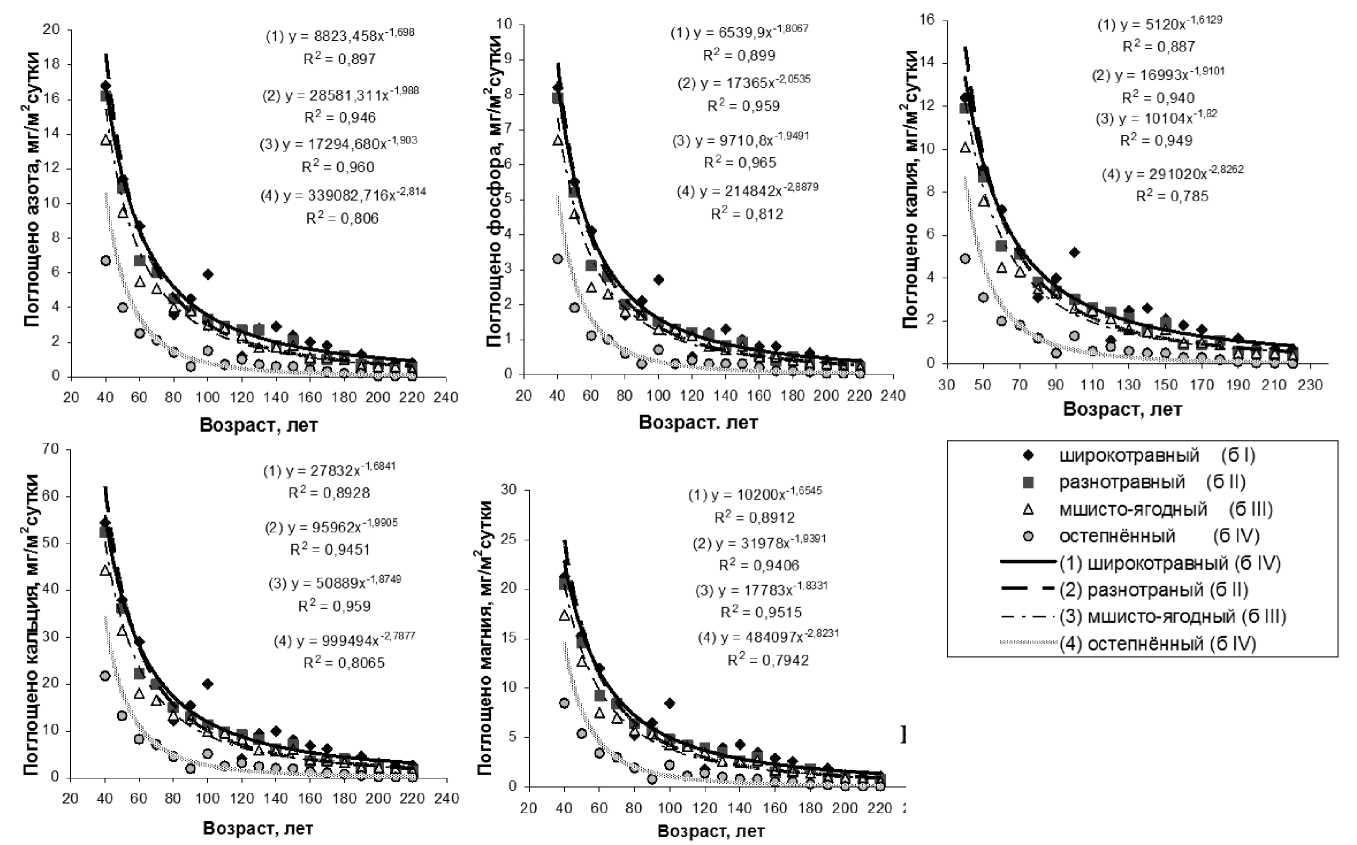
Рис. 2. Минеральная продуктивность лиственницы в разных типах леса в онтогенезе
Вестник КрасГАУ. 2013. № 1
120 п
2 о
S £ ГС г
I-о
о L. о Z л
ГС г
о о; с о
^ 100 -
к го и S о
80 -
60 -
40 -
120 -,
к
ГС ГС ГС ¥ о
§
I ф
80 -
60 -
40 -
20 -
широкотравный (б I)
(1) у = -44,238Ln(x) + 256,77
R2 = 0,782
JR ^ - Д' дЛГД'*»
(4) у = 52520,583k'1 698 v *
---Bi^W--т--°
(3) у = 2,434x06SS
R2 = 0,990 Л
. ТС
1 (2) у= 247,900х
R2 = 0,848
140 и
,-0,209
40 60 80 100 120 140 160 180 200 220 240
Возраст, лет
МШИСТО-ЯГОДНЫЙ (б III)
(4) у = 126238,541 х-1да R2 = 0,960
О I л
ГС г
о
X с о
(1) у = -46,500Ln(x) + 260,305 R2 = 0,942
А . Д'
(3) у = 2,636х0 671
R2 = 0,998
(2) у = 254,328хода R2 = 0,847
20 40 60 80 100 120 140 160 180 200 220 240
Возраст, лет
к5 юо -
ГС ГС ГС ^ о
к S I ф 7 ГС I ГС
2 S о ¥ ГС 2
с о
80 -
60 -
40 -
20 -
.100 -
К
С
Ф
Й
о
К
80 -
60 -
40 -
разнотравный (6 II)
(1)y=-46,437Ln(x) +259,147 R2 = 0,945
(4)у= 176427,843х'1 988 R2 = 0,946
40 60 80 100 120 140 160
Возраст, лет
остепнённый (б IV)
(1) у = -47,001 Ln(x)+ 256,171
R2 = 0,889
(4) у = 79754,466х'1807 R2 = 0,899
,'Д
(3) у = 2,351 х°697 R2 =0,997
(2) у = 261.619х'031Б
180 200 220 240
(3) у = 0,982х° 547
R2 = 0,978 д
д
(2)у = 257.823х-°336
R2= 0,768
40 60 80 100 120 140 160 180 200 220 240
Возраст, лет
« ЧПФ ■ БП Д КП/ФП О МП(М)
----(1)ЧПФ -- —(2)БП ----(3) КП/ФП ..............................(4)МП{Ы)
Рис. 3. Характер взаимосвязи основных физиологических показателе лиственницы в онтогенезе
Экология
Элементы по интенсивности их поглощения единицей активной поверхности корней в сутки во все возрастные периоды располагаются в порядке: Ca>Mg>N>K>P. Связь поглощения N с возрастом была высокой отрицательной по всем типам леса ( r = от -0,784 до -0,830). По P, K, Ca и Mg по всем четырем типам также наблюдалась высокая обратная корреляция поглощения их с возрастом ( r = от -0,768 до -0,812; от -0,812 до -0,863; от -0,789 до -0,836 и от -0,805 до -0,853 соответственно). С возрастом растений происходило снижение количественных значений изучаемых показателей. Небольшие колебания БП у всех бонитетов можно объяснить внутренней регуляторной деятельностью растения.
Для лучшего понимания взаимосвязи величины: БП, ЧПФ, МП (по азоту), а также отношения КП/ФП были представлены в одном масштабе в виде доли от максимальных значений, выраженных в процентах (рис. 3). Физиологические показатели сравнивались с усвоением корнями азота – ведущего элемента питания. Отношение КП/ФП в онтогенезе во всех типах леса изменялось в 3,11–4,24 раза. С возрастом во всех типах леса падала функциональная связь корневой системы с хвоей. Так, если в 40 лет у растений широкотравного типа 1 м 2 активной поверхности корней мог обслужить 9,3 м 2 поверхности хвои, то к 120 годам он мог обслужить только 2,8 м 2 . В остепненном типе в 40 лет эта величина составила 8,8, а в конце онтогенеза – 2,8 м 2 . Связь между КП/ФП с возрастом была высокой положительной ( r = от 0,989 до 0,999), а с МП – высокой обратной ( r варьировал от -0,798 до -0,849 в зависимости от типа условий). Корреляции КП/ФП с БП и ЧПФ также были отрицательными ( r варьировал соответственно от -0,723 до -0,828 и от -0,852 до -0,944). В условиях нарастающей нехватки минеральных элементов в течение онтогенеза во всех типах леса корневая система не могла активизировать поглотительную активность, и для улучшения условий питания на уроне организма смещалось соотношение КП/ФП в пользу корней, что позволяло в течение длительного времени обеспечить надземную часть дополнительными количествами элементов для поддержания основного процесса питания – фотосинтеза. Поэтому при низкой обеспеченности растений элементами увеличение ростовой реакции корней для освоения новых горизонтов почвы является ответной неспецифической адаптивной реакцией. При растущей нехватке азота и падении МП снижалась ЧПФ, а значит и БП. Связь МП с ЧПФ и БП была высокой положительной ( r был на уровне от 0,910 до 0,974 и от 0,979 до 0,998 соответственно для I –IV бонитетов). Тем не менее, падение БП происходило более медленными темпами, чем МП и ЧПФ (см. рис. 3). Закономерности изменения физиологических показателей в онтогенезе в сравнении с поглощением азота и их связи были аналогичны у растений всех типов леса и при сравнении их с поглощением P, K, Ca и Mg.
Выводы
-
1. Чистая продуктивность фотосинтеза, количество депонированного углерода и минеральная продуктивность лиственницы сибирской были максимальными в широкотравном типе лесорастительных условий и падали пропорционально ухудшению условий в 5,9–100,0, в 7,2–62,0 и от 17,7–27,3 до 330,0–490,0 раза соответственно. Биологическая продуктивность растений в изучаемом возрастном периоде изменялась во всех бонитетах в 1,8–2,2 раза. Резкое падение БП наблюдалось к 70–80 годам, после чего она снижалась крайне медленно.
-
2. Во всех типах леса в онтогенезе отмечена высокая положительная корреляционная зависимость между поглощением корнями азота и ЧПФ и БП ( r = от 0,918 до 0,964 и от 0,988 до 0,997 соответственно). При сравнении величин ЧПФ и БП с поглощением P, К, Ca и Mg результаты были близкими с азотом.
-
3. В условиях ухудшения питания с возрастом увеличение КП относительно ФП является неспецифической адаптивной реакцией, позволяющей растению за счет дополнительного минерального питания поддерживать фотосинтез на жизненно необходимом уровне.
-
4. Предложенный способ преобразования таксационных данных в физиологические позволяет получать количественные данные фотосинтетической активности, минеральной и биологической продуктивности древесных пород на уровне организма в онтогенезе, что может быть теоретической основой для разработки агроприемов, повышающих продуктивность растений.
