Влияние трития на морфо-анатомическую структуру растений вида тростник обыкновенный (Phragmites australis)
")
Автор: Янкаускас А.Б., Ларионова Н.В., Шатров А.Н.
Рубрика: Научные статьи
Статья в выпуске: 2 т.30, 2021 года.
Бесплатный доступ
В статье представлены результаты исследований морфо-анатомической структуры растений, произрастающих в условиях хронического облучения ионизирующим излучением трития. Исследования проводили на территории Семипалатинского испытательного полигона, район реки Шаган. Радиоэкологическая ситуация исследуемого района характеризуется присутствием долгоживущих радионуклидов, таких как 3Н, 90Sr и 239+240Pu. Однако наибольшую удельную активность в природных компонентах имеет 3Н. В результате исследований установлено, что при увеличении удельной активности 3Н в свободной воде растений до значений n´103-n´105 Бк/кг наблюдается уменьшение анатомических параметров стебля и листа. При исследовании влияния содержания 3Н на толщину склеренхимы и диаметра стебля установлено, что при увеличении удельной активности 3Н в диапазоне от n´102 до n´105 Бк/кг наблюдается значимое уменьшение анатомических параметров стебля. При увеличении удельной активности 3Н до значений n´104 Бк/кг значения анатомических параметров листа уменьшаются, достигая минимальных значений. При дальнейшем увеличении удельной активности 3Н до значений n´105 Бк/кг наблюдается общая тенденция к незначительному увеличению анатомических параметров.
Растения, семипалатинский испытательный полигон, морфо-анатомические параметры растений, тростник обыкновенный (phragmites australis), ионизирующее излучение, радионуклидное загрязнение, тритий, удельная активность
Короткий адрес: https://sciup.org/170179212
IDR: 170179212 | УДК: 614.876:546.11.02.3:581 | DOI: 10.21870/0131-3878-2021-30-2-133-145
Effect of tritium on the morpho-anatomical structure of the common reed (Phragmites australis)
The article presents the results of research on the morpho-anatomical structure of the plants growing under chronic exposure to ionizing radiation. The research was conducted in the territory of the Semipalatinsk test site in the vicinity of the Shagan river. The radioecological situation in the study area is determined by the presence of the long-lived radionuclides 3H, 90Sr, 239+240Pu in components comprising the natural environment. The main contributor to the radioactive contamination of the nature components is 3H. The findings of the study show that when 3H specific activity in the free water of the common reed grass increases to n´103-n´105 Bq/kg, some anatomical parameters of its cane and leaf are reduced. The study also shows that with the increase in tritium specific activity from n´102 to n´105 Bq/kg the anatomical parameters of the cane (sclerenchyma, cane diameter) decrease. When the specific activity of tritium increases to n´104 Bq/kg, the leaf anatomical parameters are reduced. When specific activity of 3H increases to n´105 Bq/kg there is the general tendency to the insignificant increase in anatomical parameters.
Текст научной статьи Влияние трития на морфо-анатомическую структуру растений вида тростник обыкновенный (Phragmites australis)
Одной из проблем бывшего Семипалатинского испытательного полигона (СИП) является загрязнение природных зон радионуклидом тритием. К такой зоне относится р. Шаган [1, 2]. Радионуклидное загрязнение вод р. Шаган обусловлено влиянием подземного термоядерного испытания с целью отработки технологии создания искусственных водоёмов, в результате которого образовался водоём «Атомное» озеро. Кроме того, загрязнение является и последствиями подземных ядерных испытаний в вертикальных скважинах на испытательной площадке «Бала-пан». Современное состояние водной среды р. Шаган характеризуется присутствием долгоживущих техногенных радионуклидов 3Н, 90Sr, 239+240Pu. Присутствие 90Sr и 239+240Pu фиксируется только в пределах выхода реки из загрязнённых грунтов «Атомного» озера на уровне 0,03 Бк/кг и 0,002 Бк/кг соответственно. Основное загрязнение компонентов экосистемы р. Шаган обусловлено повышенным содержанием радионуклида 3Н [3, 4]. На некоторых участках русла в поверхностных водах содержание 3 Н составляет 4-10 5 Бк/кг, что более чем в 50 раз превышает уровень вмешательства для питьевой воды. Содержание 3Н в свободной воде растений, произрастающих в её береговой зоне, достигает 2-10 5 Бк/кг. Значения удельной активности 3 Н в свободной воде растений и органически связанном веществе достигают n x 102-n x 105 Бк/кг [5, 6].
Особенностью последствий ядерных испытаний является формирование высоких дозовых нагрузок на биоту до уровней, способных привести к разрушению наиболее радиочувствитель-
Янкаускас А.Б. * – инженер; Ларионова Н.В. – учёный секретарь, к.б.н.; Шатров А.Н. – начальник лаборатории. Филиал ИРБЭ НЯЦ РК. *Контакты: 071100, Республика Казахстан, Восточно-Казахстанская обл., Курчатов, ул. Бейбит-Атом, 2. Тел.: 8(72251) 3-29-13;
ных экосистем. В настоящее время признано, что система нормирования радиационного воздействия на биоту должна быть ориентирована на защиту популяций. Поэтому исследования биологических эффектов в популяциях, населяющих территории, контрастные по уровню и спектру дозообразующих радионуклидов, играют уникальную роль в развитии и обосновании принципов экологического нормирования [7]. Растительные популяции, подвергающиеся хроническому облучению, могут характеризоваться, как ускоренным мутационным процессом, так и различными в зависимости от мощности дозы, уровнями активации систем адаптации к стрессовым факторам окружающей среды. Вместе с тем, именно ситуация хронического облучения представляет особый интерес с точки зрения установления пределов допустимого радиационного воздействия на компоненты экосистем [8, 9].
Интерес к 3Н с точки зрения его влияния на растительный организм вызван его способностью с высокой скоростью включаться в биологические процессы. Вместе с тем, инкорпорированный в органические соединения тритий представляет определённую опасность, так как он обладает очень низкой энергией радиоактивного излучения. Данный факт служит причиной больших поглощённых доз в местах включения этого радионуклида. В связи с чем, он может являться мощным мутагенным фактором. Тритий в составе молекул воды (тритированная вода) очень легко усваивается растениями, как через устьичный аппарат, так и в результате корневого поглощения. В дальнейшем 3Н инкорпорируется в органические структуры растения вследствие метаболических процессов, в частности фотосинтетических реакций [10-13]. Примерно 10-30% органической фракции, содержащей инкорпорированный 3Н, может варьировать при изменении концентрации радионуклида в окружающей среде [14]. Если никаких изменений концентраций 3Н в окружающей среде не происходит, то его содержание в растениях находится на неизменном уровне. Включение 3Н в состав органического вещества может привести к накоплению данного радионуклида в органах и тканях растений [15].
До настоящего времени, вопрос о влиянии 3Н на морфо-анатомическую структуру растений тростника ( Phragmites australis ) на территории СИП не изучен. Таким образом, цель нашей работы – выявить наличие изменений в морфо-анатомической структуре растений, произрастающих в условиях тритиевого загрязнения.
Материалы и методы
В рамках исследования морфо-анатомических параметров растений, произрастающих на участках радионуклидного загрязнения СИП, в качестве территории обследования выбран район р. Шаган. Исследовательские площадки закладывали вдоль по руслу, всего заложено 10 исследовательских площадок (рис.1). Каждая площадка представляла собой участок площадью примерно 1 м2. Расстояние между площадками составило от 100 до 1000 м. По результатам измерения мощности эквивалентной дозы (МЭД) и плотности потока β-частиц можно отнести данные участки к фоновым, основные значения МЭД равны 0,09-0,10 мкЗв/ч, плотность потока β-частиц <10 част/мин⋅см2. В качестве опытного растения выбран тростник обыкновенный ( Phragmites australis ).
В условиях эксперимента в естественных условиях оказать влияние на ход исследования могут абсолютно любые факторы, как биотические, так и абиотические. Таким образом, помимо определения радионуклидного состава, одной из задач стало определение элементного состава воды и растений. Аналогичным образом в местах отбора проб растений произведён отбор проб воды для проведения химического анализа и определения степени минерализации. Для морфологических исследований отобраны надземные части растения, проведены измерения основных параметров, таких как высота растения, длина метёлки, длина листа. Для исследования анатомических параметров с каждого растения отбирали фрагменты стеблей и листьев. В полевых условиях образцы органов растений фиксировали с использованием копенгагенской смеси, которую готовили путём смешивания: этиловый спирт 96% – 13 мл, глицерин – 0,5 мл, дистиллированная вода – 5 мл. Данная смесь подходит для фиксации материала, если в дальнейшем его не предполагается обезвоживать и пропитывать парафином.
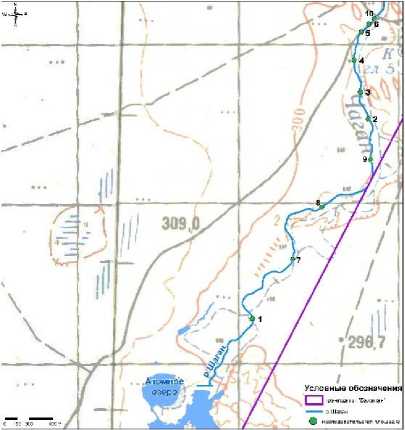
Рис. 1. Расположение точек отбора проб.
Из фиксированных образцов получали препараты для исследования. Из препаратов готовили срезы с использованием микротома. Толщина среза образца зависела от состояния растения и жёсткости волокон. В среднем для стебля толщина срезов составила 25-30 мкм, для листьев – 30 мкм. Полученный препарат исследовали при помощи тринокулярного микроскопа Microscope Rose MC 300 (Austria) с цифровой камерой Vision Cam V500/21 (Austria). При помощи камеры и программного обеспечения BioWizard 4.2 делали снимки и измерения параметров исследуемых образцов. В качестве исследуемых параметров стебля выбраны: диаметр стебля, толщина эпидермы, толщина склеренхимы, площадь проводящих пучков, расположенных в паренхиме. В качестве исследуемых параметров листа выбраны: толщина верхнего эпидермиса, толщина нижнего эпидермиса, диаметр пузыревидных клеток эпидермы, толщина мезофилла, площадь проводящего пучка 1-го порядка, площадь проводящего пучка 2-го порядка.
Содержание 3Н в растениях определялось в свободной воде, полученной в результате конденсации транспирируемой жидкости растениями внутри специальных герметичных установок с водосборниками. Все образцы подготовлены для проведения бета-спектрометрического анализа по определению 3Н в свободной воде растений и органической составляющей с использованием автоматической установки для сжигания и подготовки проб для определения органически связанного трития Sample Oxidyzer (USA). Измерение 3Н проводилось при помощи жидкосцинтилляционного спектрометра Quantulus 1220 (USA).
Определение металлов в воде проводилось согласно методике ИСО 17294-2 «Качество воды – применение масс-спектрометрии с индуктивно связанной плазмой. Часть 2: Определение
62 элементов». Определение содержания металлов проводилось методом ИСП-МС и ИСП-АЭС, с использованием квадрупольного масс-спектрометра Elan 9000 фирмы «Perkin Elmer SCIEX» (USA) в комплекте с компьютером и специализированным программным обеспечением.
За период исследования произведено порядка 8000 измерений анатомических параметров стебля и листа. Первым шагом в статистической обработке данных была проверка распределения морфо-анатомических параметров на нормальность, которую осуществляли с использованием критерия Колмогорова-Смирнова. Для сравнения эмпирического распределения с теоретическим используется критерий λ , который рассчитывается согласно выражению:
■^ эмпир = ^, (1)
где D – наибольшее значение абсолютной разности между накопленными частотами эмпирического и теоретического распределений; n – объём выборки.
Теоретическое значение λ не зависит от объёма выборки и числа степеней свободы, а определяется только уровнем значимости. Для уровня значимости 0,05 значение λ=1,36. Если Аэмпир < -^0,05, то нет оснований считать, что эмпирические и теоретические частоты отличаются существенно. Таким образом, в данной работе выполнено сравнение теоретических частот нормального распределения с частотами эмпирического распределения [16]. Как следует из расчётов, в подавляющем большинстве случаев значение Яэмпир < Ятеорет, что говорит о том, что эмпирические распределения морфо-анатомических параметров от нормального распределения отличаются несущественно. Исключением являются параметры склеренхимы и площади проводящего пучка в пробах площадки 4. Здесь величина Яэмпир существенно превышает значение Ятеорет. В данных случаях делается предположение о логарифмически нормальном распределении. Для проверки данной гипотезы от значений параметров склеренхимы и площади проводящего пучка были взяты натуральные логарифмы. Полученные значения были вновь проверены на нормальность распределения с использованием критерия Колмогорова-Смирнова. Расчёты показали, что значения натуральных логарифмов склеренхимы и площади проводящего пучка в пробе площадки 4 распределены по нормальному закону. Тогда исходные значения данных параметров распределены по логарифмически нормальному закону. На рис. 2 демонстрируются типичные распределения морфо-анатомических параметров по частоте встречаемости (эмпирические и теоретические).
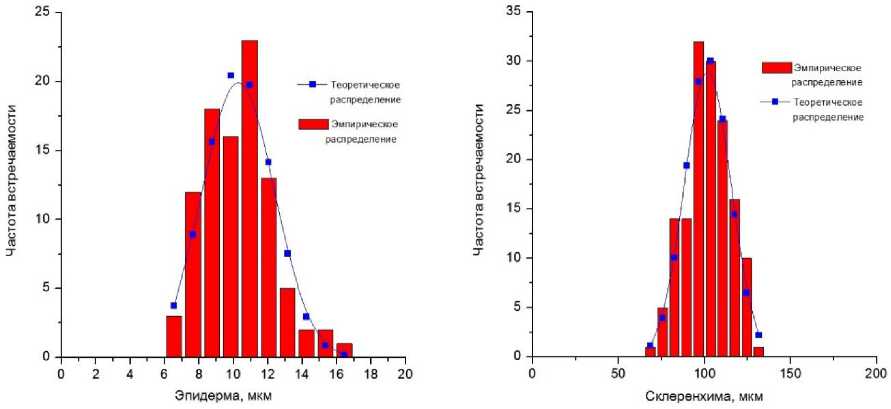
Рис. 2. Типичные эмпирические и теоретические распределения анатомических параметров по частоте встречаемости.
Как показано на графиках (рис. 2), практическое распределение значений стебля соответствует теоретическому распределению. Таким образом, мы можем утверждать, что полученные данные достоверны и могут быть использованы для дальнейших аналитических расчётов.
Аналогичным образом получены значения Я эмпир для всех морфо-анатомических параметров листа для каждой площадки. Как и в предыдущем случае, установлено, что в подавляющем большинстве значение Я эмпир < Я теорет . Исключением являются параметры мезофилла и площади проводящего пучка 2-го порядка в образцах площадки 2 и 10. Здесь величина Я эмпир превышает значение Я теорет . В данных случаях делается предположение о логарифмически нормальном распределении.
Результаты и обсуждение
С целью подтверждения идентичности условий произрастания все образцы растений и воды прошли элементный анализ (табл. 1).
Таблица 1 Элементный состав воды в точках отбора
|
№ |
Общая минерализация воды, мкг/г |
Содержание элементов в воде, мкг/л |
||||||||
|
Li |
Al |
Mn |
Co |
Ni |
Cu |
Mo |
Pb |
U |
||
|
1 |
7500 |
29±2 |
10±0,1 |
176±0,4 |
1,4±0,05 |
6,5±0,1 |
11±0,4 |
21±1 |
0,8±0,04 |
16±0,1 |
|
2 |
6600 |
36±3 |
6,0±0,3 |
46±1 |
1,7±0,03 |
6,4±0,5 |
7,0±0,4 |
14±1 |
0,14±0,04 |
17±0,4 |
|
3 |
6800 |
51±4 |
4,2±0,3 |
85±1 |
1,8±0,04 |
7,0±0,2 |
9,1±0,3 |
22±0,4 |
0,2±0,05 |
27±0,4 |
|
4 |
7700 |
34±3 |
5,6±0,1 |
42±1 |
1,2±0,02 |
4,5±0,2 |
8,4±0,1 |
14±1 |
0,1±0,01 |
18±0,1 |
|
5 |
6700 |
41±1 |
7,0±0,3 |
34±1 |
1,0±0,03 |
3,5±0,4 |
4,6±0,1 |
17±1 |
0,6±0,02 |
25±0,5 |
|
6 |
6700 |
28±1 |
6,7±0,6 |
43±0,2 |
0,8±0,06 |
0,7±0,1 |
3,0±0,1 |
12±0,4 |
0,4±0,06 |
16±0,1 |
|
7 |
4200 |
22±2 |
8,5±0,1 |
123±1 |
1,0±0,05 |
4,4±0,5 |
10±0,1 |
13±0,3 |
0,1±0,04 |
12±0,1 |
|
8 |
6600 |
26±2 |
9,1±0,2 |
14,0±0,1 |
1,0±0,03 |
3,4±0,4 |
7,9±0,1 |
14±0,3 |
0,1±0,03 |
13±0,2 |
|
9 |
5600 |
59±1 |
4,1±0,1 |
150±1 |
3,2±0,02 |
10±0,2 |
10±0,2 |
13±1 |
0,2±0,02 |
31±0,1 |
|
10 |
7300 |
70±6 |
12,0±0,4 |
180±2 |
1,8±0,1 |
8,9±0,4 |
9,5±0,1 |
27±1 |
9,0±0,2 |
41±0,2 |
Полученные значения элементного анализа воды для каждой исследовательской площадки соответствуют нормальным значения для водоёма данного типа и общей минерализацией в 5600-7700 мкг/г. Уран, зафиксированный в пробах воды, имеет природное происхождение и обусловлен геохимическими особенностями данного региона. Значения концентрации элементов в образцах воды находятся приблизительно в одном диапазоне значений. Из чего следует, что при обнаружении потенциальных отклонений в растениях влияние элементного состава воды можно исключить. Для полной картины образцы растений прошли проверку на химический состав (табл. 2).
Таблица 2
Содержание элементов в образцах растений
|
Элементный состав |
||||||||||
|
Al |
Mn |
Fe |
Co |
Ni |
Cu |
Zn |
Cd |
Pb |
U |
|
|
Содержание элементов в растениях, мг/кг |
9-30 |
170-470 |
66-100 |
0,05-0,13 |
0,9-2,9 |
1,3-19 |
4,2-16 |
<п.о. 0,01 |
0,11 – 7,0 |
0,01-0,08 |
|
Диапазон нормальных значений |
до 200 |
20-400 |
50-100 |
0,02-1 |
0,1-5 |
5-23 |
до 45 |
0,05-0,2 |
до 8 |
0,005-0,08 |
При анализе полученных данных элементного анализа проб воды и растений и для выявления наиболее важных элементов-загрязнителей использован метод сравнения полученных результатов со встречающимися типичными концентрациями элементов в растениях по данным исследователей [17, 18]. Содержание элементов в растениях для всех исследовательских площадок идентично. Значения концентраций элементов в растениях на исследовательских площадках не превышают значений опубликованных данных. Из чего следует, что идентичность условий произрастания опытных образцов установлена, и можно исключить влияние такого фактора, как загрязнение воды и растений металлами.
Радионуклидный анализ. На основе лабораторных анализов получены значения удельной активности радионуклида 3Н в свободной воде растений и в органической составляющей (табл. 3).
Содержание 3Н в свободной воде растений и в органическом веществе
Таблица 3
|
№ |
Удельная активность 3Н в свободной воде, Бк/кг (M±m) |
Удельная активность 3Н в органической составляющей, Бк/кг (M±m) |
|
1 |
580 ± 60 |
230 ± 30 |
|
2 |
900 ± 90 |
390 ± 50 |
|
3 |
6,5 х 104 ± 0,7 х 104 |
2,4 х 104 ± 0,2 х 104 |
|
4 |
2,9 х 104 ± 0,3 х 104 |
5,4 х 103 ± 0,5 х 103 |
|
5 |
1,3 х 104 ± 0,1 х 104 |
4,2 х 103 ± 0,4 х 103 |
|
6 |
8,0 х 103 ± 0,8 х 103 |
2,7 х 103 ± 0,3 х 103 |
|
7 |
1,0 х 103 ± 0,1 х 103 |
400 ± 50 |
|
8 |
1,2 х 103 ± 0,1 х 103 |
390 ± 50 |
|
9 |
850 ± 90 |
410 ± 50 |
|
10 |
5,9 х 103 ± 0,6 х 103 |
2,0 х 103 ± 0,2 х 103 |
По результатам проведённых исследований установлено, что удельная активность 3Н в свободной воде растений достигает значений 6,5 х 104±0,7 х 104 Бк/кг, в органической составляющей - 2,4 х 104±0,2 х 10 4 Бк/кг.
Морфо-анатомические параметры. Вид Phragmites australis - многолетнее травянистое растение. На основе лабораторных и аналитических работ произведен расчёт средних арифметических значений морфологических параметров с ошибкой репрезентативности (M±m). Высота растений колеблется в диапазоне 110±15 - 150±25 см (норма 100-400 см), длина листа 22±4 -30±4 см (норма 5-30 см), метелки 15±4 - 19±9 см (норма до 50 см). Значения морфологических параметров соответствуют нормальным встречающимся значениям для тростника Phramites australis [19].
Стебель полый, трубчатый, волокна жёсткие, диаметр стебля варьирует от 1,7±0,1 до 3,4±0,1 мм. Покровная ткань, представленная эпидермой, состоит из одного слоя клеток, толщина которых варьирует от 8,4±0,2 до 11,0±0,6 мкм. Клетки овальной, слегка продолговатой формы. На поперечном срезе видно, что стебель пронизан тяжами проводящих пучков, состоящих из сосудов ксилемы, погруженных в паренхиму (рис. 3). Площадь проводящего пучка стебля варьирует в пределах от 14,2±1,0 до 40,0±1,0 мкм2.
Сосуды ксилемы в проводящих пучках стебля Phragmites australis расположены беспорядочно, обеспечивая транспорт воды и минеральных веществ от корня по стеблям к остальным органам и тканям растения, а также выполняя функцию арматуры, что особенно важно для травянистых растений, и растений, лишённых древесного прочного стебля, как в случае с тростником. Толщина склеренхимы варьирует от 97±2,2 до 170±4,7 мкм.
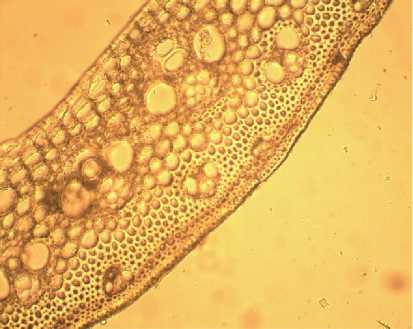
Рис. 3. Поперечный срез стебля Phragmites australis (х 20).
Листовые пластинки линейно-ланцетного типа. Толщина листа в поперечном срезе достигает 160±27 - 220±29 мкм (рис. 4). Покровная ткань листа представлена однослойным эпидермисом, форма клеток прямоугольная. В зависимости от стороны листа размеры клеток адаксиального и абаксиального эпидермиса различаются. Толщина адаксиального (верхнего) эпидермиса варьирует от 10,6±0,3 до 12,6±0,4 мкм, толщина абаксиального (нижнего) эпидермиса варьирует от 12,1 ±0,3 до 15,2±0,6 мкм. Произрастая в достаточно засушливых климатических условиях, эпидерма листьев выработала защитные приспособления: в наиболее засушливое время листья имеют свойство сворачиваться в трубочку за счёт расположения на адаксиальной стороне пузыревидных (водоносных) клеток эпидермиса, расположенных в виде продольных тяжей. Таким образом, пузыревидные (моторные) клетки способствуют сжиманию и разжиманию листовой пластинки, снижая возможность испарения. Диаметры пузыревидных (водоносных) клеток эпидермиса достигают значений от 75,0±2,4 до 110,5±2,1 мкм.
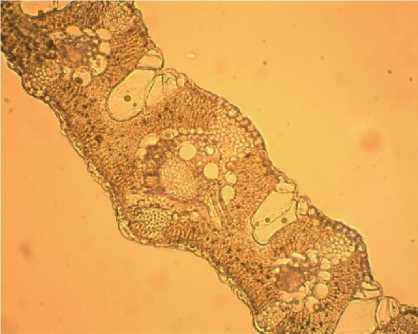
Рис. 4. Поперечный срез листа Phragmites australis (х20) .
Между двумя слоями эпидермы располагается слой мезофилла (хлоренхима), толщина которого варьирует от 120±5,1 до 200±7,3 мкм. Клетки неправильной округлой формы либо слегка вытянуты, однородны, размеры различны. Имеются включения в виде хлоропластов, погружённых в цитоплазму клеток мезофилла. Проводящая ткань листа представлена пучками 1-го и 2-го порядков, различающихся хорошо развитыми сосудами ксилемы, осуществляющими перенос воды от корней в листья. Площадь проводящего пучка 1-го порядка варьирует от 13000±1200 до 27000±1600 мкм2, площадь проводящего пучка 2-го порядка варьирует от 4200±300 до 13500±600 мкм2.
Зависимость анатомических параметров стебля от содержания 3Н в растениях. Изменения анатомических показателей стебля и листа от содержания 3Н в свободной воде и в органическом веществе рассмотрены графическим способом. При изучении эмпирических рядов, нанесённых на график, установлено, что количественные изменения параметров диаметра стебля и толщины склеренхимы происходят линейно и выражаются уравнением параболы первого порядка, т.е. прямой y=a-bx . При рассмотрении эмпирических рядов регрессии для толщины эпидермы и площади проводящего пучка, сделано предположение, что в данном случае имеются закономерности параболической ниспадающей, выражаемой уравнением параболы второго порядка, вида y=a+bx+cx2 . В результате расчётов установлено, что содержание 3Н в органически связанном веществе растений изменяется прямо пропорционально содержанию 3Н в свободной воде, и коэффициент корреляции между этими переменными соответствует значению 0,97. Таким образом, в качестве примера на графиках рис. 5 представлен характер изменений анатомических параметров стебля от изменений удельной активности 3Н в свободной воде растений.
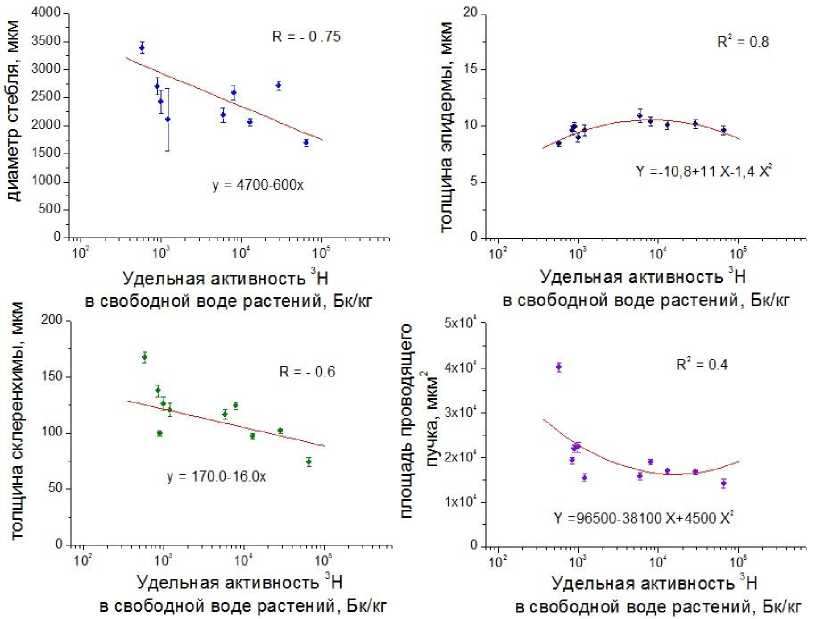
Рис. 5. Зависимость анатомических параметров стебля от содержания 3Н в свободной воде растений.
При исследовании влияния содержания трития на толщину склеренхимы и диаметра стебля установлено, что при увеличении удельной активности 3 Н в диапазоне nx 10 2 -10 5 Бк/кг наблюдается значимое уменьшение анатомических параметров. При изучении влияния содержания 3Н на толщину эпидермы и площади проводящего пучка при построении эмпирических рядов регрессии была обнаружена иная тенденция: при увеличении удельной активности трития до значений nx10 3 -10 4 Бк/кг, величина толщины эпидермы возрастает, а затем происходит незначительное уменьшение параметров, значения площади проводящего пучка уменьшаются.
В целом, установлена тенденция к изменениям анатомических параметров стебля от изменения содержания в них трития. Данный факт подтверждают значимые значения коэффициентов регрессии и детерминации. Предполагается, что значение коэффициента детерминации превышает 0,5. В этом случае коэффициент множественной корреляции превышает 70%. Такие модели можно признать достоверными и указывающими на функциональную зависимость между переменными.
Изменения анатомических показателей листа от содержания 3Н в свободной воде и в органическом веществе также рассмотрено графическим способом. При изучении эмпирических рядов, нанесённых на график (рис. 6), установлено, что количественные изменения параметров верхнего и нижнего эпидермиса происходят линейно и выражаются уравнением параболы первого порядка, т.е. прямой y=a+bx. При рассмотрении эмпирических рядов регрессии для осталь- ных изучаемых параметров, сделано предположение, что в данном случае имеются закономер- ности параболической ниспадающей, выражаемой уравнением параболы второго порядка, вида y=a+bx+cx2.
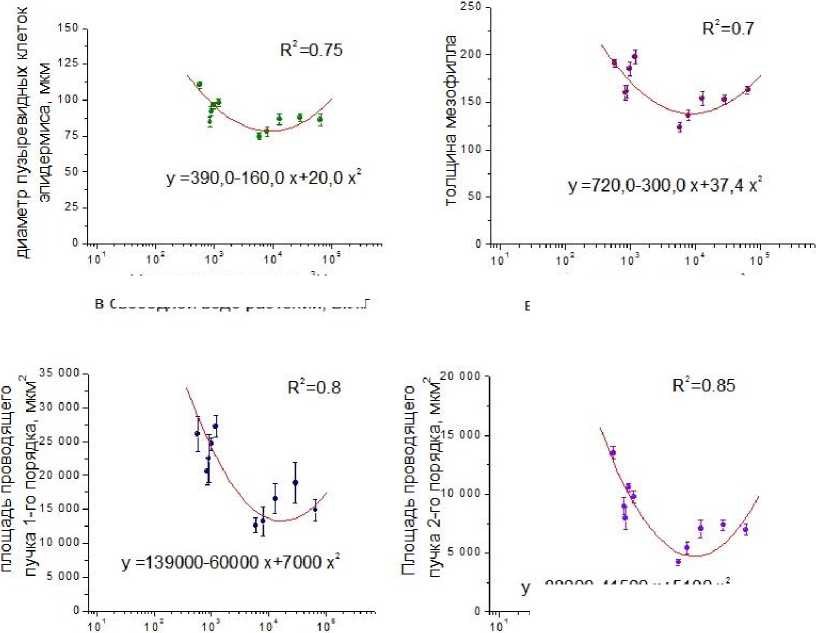
Удельная активность 3Н в свободной воде растений, Бк/кг
=88000-41500 х+5100 х
Удельная активность 3Н в свободной воде растений, Бк/кг
Рис. 6. Зависимость анатомических параметров листьев от содержания 3Н в свободной воде растений.
105 10* 10s
Удельная активность5 Н в свободной воде растений, Бк/кг
Удельная активность *Н свободной воде растений, Бк/кг
При изучении анатомических параметров листа установлено, что толщина верхнего и нижнего эпидермиса с увеличением удельной активности 3Н в свободной воде не изменяется. Коэффициент регрессии (R=-0,2 – -0,3) подтверждает отсутствие статистической связи между объясняемой переменной (толщиной эпидермиса) и факторами (удельной активностью 3Н).
При построении эмпирических рядов регрессии для остальных изучаемых параметров, а именно диаметра пузыревидных клеток, толщины мезофилла, площади проводящих пучков листа 1-го и 2-го порядков, сделано предположение, что здесь имеет место закономерность параболической ниспадающей, выражаемой уравнением параболы второго порядка, вида y=a+bx+cx2 . При увеличении удельной активности 3 Н до значений nx10 4 Бк/кг значения анатомических параметров листа уменьшаются, достигая минимальных значений. При дальнейшем увеличении удельной активности 3 Н до значений nx10 5 Бк/кг наблюдается общая тенденция к незначительному увеличению анатомических параметров. Коэффициент детерминации (R2=0,7 – 0,85) подтверждает наличие статистической связи между объясняемой переменной (диаметр пузыревидных клеток, толщина мезофилла, площадь проводящего пучка 1-го и 2-го порядков) и аргументом (удельной активностью 3Н в свободной воде растений).
Заключение
В результате проведённых исследований установлено наличие влияния радионуклида 3Н на анатомическую структуру растения тростник (Phragmites australis) . Произведена статистическая обработка данных. Получено типичное для всех площадок распределение значений анатомических параметров стебля и листа. В ходе исследований установлено, что при увеличении удельной активности 3 Н в свободной воде растений до значений nx10 3 -nx10 5 наблюдается уменьшение таких анатомических параметров стебля, как диаметр, толщина склеренхимы, площадь проводящего пучка; происходит изменение анатомических параметров листа таких, как толщина мезофилла, диаметр пузыревидных клеток, площадь проводящего пучка 1-го и 2-го порядков. Поскольку в данном эксперименте мы имеем дело, по большому счёту, с внутренним облучением от радионуклида 3Н, мы можем предполагать, что изменения на анатомическом уровне обусловлены специфичностью радионуклида 3Н, который, являясь изотопом водорода, имеет аналогичные свойства и ведёт себя подобно стабильному элементу: способен встраиваться в клеточные стенки растений.
Список литературы Влияние трития на морфо-анатомическую структуру растений вида тростник обыкновенный (Phragmites australis)
- Янкаускас А.Б., Ларионова Н.В., Шатров А.Н. Исследование морфо-анатомических показателей растений, произрастающих в местах испытания боевых радиоактивных веществ //Современные проблемы радиологии и агроэкологии, пути реабилитации техногенно-загрязнённых угодий: сб. докл. межд. конф., Обнинск, 15 декабря 2016 г. Обнинск: ФГБНУ ВНИИРАЭ, 2016. С. 208-212.
- Янкаускас А.Б., Ларионова Н.В. Морфо-анатомические исследования растений на территории СИП //Актуальные вопросы мирного использования атомной энергии: мат. межд. конф. молодых учёных и специалистов, Алматы, 6-8 июня 2012 г. Алматы: ИЯФ НЯЦ РК, 2012. С. 188-189.
- Lyakhova O.N., Lukashenko S.N., Larionova N.V., Tur Y.S. Contamination mechanisms of air basin with tritium in venues of underground nuclear explosions at the former Semipalatinsk test site //J. Environ. Radioact. 2012. V. 113. P. 98-107.
- Aidarkhanova A.K., Lukashenko S.N., Larionova N.V., Polevik V.V. Radionuclide transport in the "sediments - water - plants" system of the water bodies at the Semipalatinsk test site //J. Environ. Radioact. 2018. V. 184-185. P. 122-126.
- Айдарханов А.О., Лукашенко С.Н., Субботин С.Б. Радиоэкологическое состояние бывшего Семипалатинского испытательного полигона и прилегающих территорий //Актуальные вопросы радиоэкологии Казахстана: сб. трудов Института радиационной безопасности и экологии за 2007-2009 гг. Павлодар: Дом печати, 2010. Вып. 2. С. 9-55.
- Aidarkhanov A.O., Lukashenko S.N., Lyakhova O.N., Subbotin S.B., Yakovenko Yu.Yu., Genova S.V., Aidarkhanova A.K. Mechanisms for surface contamination of soils and bottom sediments in the Shagan river zone within former Semipalatinsk nuclear test site //J. Environ. Radioact. 2013. V. 124. P. 163-170.
- Гераськин С.А. Хроническое радиационное воздействие на популяции растений //Биологические эффекты малых доз ионизирующей радиации и радиоактивное загрязнение среды: матер. межд. конф., Сыктывкар, Республика Коми, Россия, 17-21 марта 2014 г. Сыктывкар, 2014. С. 45-46.
- Удалова А.А., Ульяненко Л.Н., Алексахин Р.М., Гераськин С.А., Филипас А.С. Методология оценки допустимого воздействия ионизирующих излучений на агроценозы //Радиационная биология. Радиоэкология. 2010. Т. 50, № 5. С. 572-581.
- Larionova N.V., Lukashenko S.N., Lyakhova O.N., Aidarkhanov A.O., Subbotin S.B., Yankauskas A.B. Plants as indicators of tritium concentration in ground water at the Semipalatinsk test site //J. Environ. Radioact. 2017. V. 177. P. 218-224.
- Сыроватко В.А. Тритийсодержащая вода в процессах водообмена растений: дис... канд. биол. наук. Киев: Институт физиологии растений, 1984.
- Melintescu A., Galeriu D.A. Versatile model for tritium transfer from atmosphere to plant and soil //Radiopro-tection. 2005. V. 40. P. 437-442.
- Choi Y.H., Lim K.M., Lee W.Y., Diabate S., Strack S. Tissue free water tritium and organically bound tritium in the rice plant acutely exposed to atmospheric HTO vapor under semi-outdoor conditions //J. Environ. Radioact. 2002. V. 58, N 1. P. 67-85.
- Keum D., Lee H., Kang H., Jun I., Choi Y., Lee C. Prediction of tritium level in agricultural plants after short term exposure to HTO vapor and its comparison with experimental results //Health Phys. 2006. V 90, N 1. P. 42-55.
- Pointurier F., Baglan N., Alanic G. A method for the determination of low-level organic-bound tritium activities in environmental samples //Appl. Radiat. Isot. 2004. V. 61, N 2-3. P. 293-298.
- Calmon P., Garnier-Laplace J. Tritium and the environment. Radionuclide fact sheet. Institute of radiopro-tection, 2010. 26 p.
- Шестаков Ю.Г. Математические методы в геологии. Учебное пособие для студентов геологических специальностей. Красноярск, 1988. 54 c.
- Horowitz C.T., Schock H.H., Horowitz-Kisimova L.A. The content of scandium, thorium, silver, and other trace in different plant species //Plant Soil. 1974. V. 40. P. 397-403.
- Laul J.C., Weimer W.C., Rancitelli L.A. Biogeochemical distribution of rare earths and other trace elements in plants and soils //Origin and Distribution of the Elements, Vol. 11 /Ed.: L.H. Ahrens. Oxford: Pergamon Press, 1979. P. 819-827.
- Губанов И.А. Тростник обыкновенный, или южный. Иллюстрированный определитель растений Средней России. В 3 т. Т. 1. Папоротники, хвощи, плауны, голосеменные, покрытосеменные (однодольные). М.: Т-во науч. изд. КМК, Ин-т технолог. иссл., 2002. 285 c.
