Влияние условий подготовки и проведения эксперимента на показатели функционирования изолированной перфузируемой печени крысы

Автор: Шадрин К.В., Пахомова В.Г., Рупенко А.П., Моргулис И.И.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Экология
Статья в выпуске: 3, 2013 года.
Бесплатный доступ
В настоящем исследовании определяли влияние условий перфузии (длительность подготовительного периода, норадреналин и молочная кислота) на показатели функционирования изолированной печени крысы.
Перфузия изолированной печени, метаболизм, норадреналин, глюконеогенез, гипоксия
Короткий адрес: https://sciup.org/14082967
IDR: 14082967 | УДК: 577.359
The influence of preparation and experiment conducting conditions on the functioning characteristics of the isolated perfused rat liver
The effect of the perfusion conditions (duration of the preparatory period, noradrenaline and lactic acid) on the characteristics of isolated rat liver functioning is determined in the present research.
Текст научной статьи Влияние условий подготовки и проведения эксперимента на показатели функционирования изолированной перфузируемой печени крысы
Введение. Исследование метаболизма печени и нахождение пределов функционирования этого органа – очень важная задача современных исследований в фармакологии, физиологии и медицине. Для решения таких задач подходит метод перфузии изолированных органов [1, 6]. Он позволяет в реальном времени отслеживать реакцию органа на различные воздействия, а также дает уникальную возможность поддерживать его гомеостаз [8]. При этом оценивают содержание различных метаболитов [10, 11], медиаторов [7, 9], факторов, которые регулируют накопление гликогена и выброс глюкозы в среду [4, 5]. Однако не уделено достаточно внимания функционированию печени как самостоятельного целостного органа, в частности тому, как влияет длительность ишемического периода на качество перфузии, как осуществляется метаболизм печени при стимуляции и как этот орган в условиях изолированной перфузии выполняет одну из присущих ему метаболических функций, например производство глюкозы из молочной кислоты [2, 3]. Несо- мненно, параметры функционирования изолированного органа зависят от условий перфузии, в частности, от состава перфузионной среды [12, 13]. При этом технологии самой операции и процессы выделения и размещения органа в термостатируемой камере (т.е. подготовительный период) вызывают в органе состояние гипоксии, что может также отражаться на его функционировании в дальнейшем. Поэтому очень важно знать, как длительность подготовительного периода влияет на метаболизм изолированных органов.
Целью работы было сравнить функционирование изолированной перфузируемой печени при различной длительности подготовительного периода и оценить влияние добавок в перфузионную среду, при использовании которых орган лучше сохраняет функциональную активность. В качестве стресс-воздействия использовали введение в перфузионную систему норадреналина. В качестве функциональной нагрузки на печень использовали лактат.
Методика исследования. Эксперименты проводили на крысах-самках Вистар массой 200–260 г. У наркотизированных (натрий-тиопентал, 100 мг/кг, в/б) животных стабилизировали гемостаз (гепарин, 1000 ед./кг, в/в); канюлировали воротную вену, инфузировали охлажденным раствором (50% – среда199, 50% – солевой раствор Кребса-Хенселейта с добавлением 80 г/л полиглюкина); канюлировали верхнюю полую вену, после чего орган выделяли и помещали в установку для культивирования изолированных органов мелких лабораторных животных «Гомеостат-3М». Печеночную артерию и нижнюю полую вену лигировали. Таким образом, вход перфузионной жидкости в печень осуществлялся через воротную вену, а выход – через верхнюю полую вену.
Перфузию печени крыс проводили в нормотермических условиях при постоянной скорости тока перфузата ( ~ 3-3,5 млтъ мин -1 ).
По длительности подготовительного периода животных разделили на три группы: 1-я группа – 25–35 мин, 2-я группа – 36–45, 3-я группа – 46–55 мин. Всего в эксперименте использовано 30 животных (по 10 в каждой группе). Длительность перфузии составляла 60 мин.
В экспериментах, где исследовали влияние добавок в перфузионную среду, использовали 22 животных. Все перфузируемые органы были разделены на 3 группы: 1 – контроль (органы 12 животных), без введения норадреналина; 2 – органы, в перфузионную систему которых вводили норадреналин (5) (вводили c 60 по 70 мин перфузии в концентрации 1мкМ); 3 – органы, в перфузионную систему которых добавляли лактат (5) (вводили с 70 по 80 мин перфузии в концентрации 10М). Длительность перфузии составляла 120 мин.
В ходе перфузии определяли следующие показатели жизнедеятельности органа: уровень давления в воротной вене, скорость желчетока, удельные скорости потребления кислорода и выделения углекислого газа, отношение удельной скорости потребления кислорода к удельной скорости выделения углекислого газа, концентрации молочной, пировиноградной кислот, глюкозы, отношение концентраций молочной и пировиноградной кислот в оттекающей от органа жидкости, скорости изменения содержаний молочной и пировиноградной кислот в оттекающей от органа жидкости.
Давление в воротной вене определяли манометрическим методом (см водн. ст.), удельные скорости потребления кислорода и выделения углекислого газа (мл/мин/г) определяли по формуле dVO2 /dt = Qt (Pa — Pv ) - где Qt – минутный объем кровообращения;
( P a — P v ) — разность напряжений кислорода в артериальной и венозной ветвях циркуляции.
Концентрации глюкозы, лактата и пирувата (мМ) определяли спектрофотометрически. Скорости изменения содержаний молочной и пировиноградной кислот (мМ/мин) определяли по формуле dC/dt = (C„1 - C^/At, где C – концентрация в момент i-го измерения;
C i + 1 - концентрация в момент измерения i+1 ;
A t - интервал между забором соответствующих проб, мин.
Результаты были статистически обработаны c использованием t-критерия Стьюдента.
Результаты исследования
Влияние длительности подготовительного периода на показатели жизнедеятельности изолированной перфузируемой печени крысы
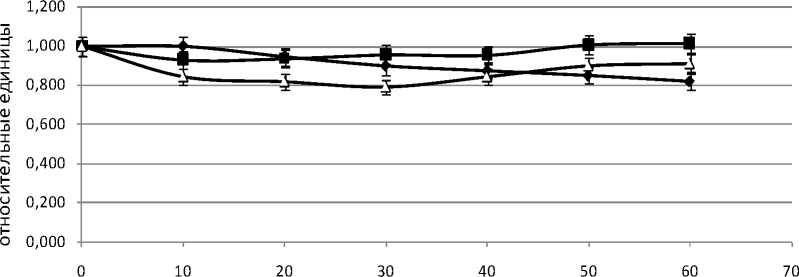
Данные свидетельствуют о сохранении в условиях перфузии функциональной активности печени: во всех экспериментальных группах изолированный орган хорошо выделяет желчь и стабилизирует уровни давления в воротной вене. Наиболее близкий к исходному уровень давления в воротной вене наблюдали для органов животных группы 2 (рис. 1).
время, мин
1 —■— 2 —2s— 3
Рис.1. Динамика давления в воротной вене в системе изолированной перфузируемой печени крыс при различной длительности подготовительного периода (1, 2, 3 – номер экспериментальной группы)
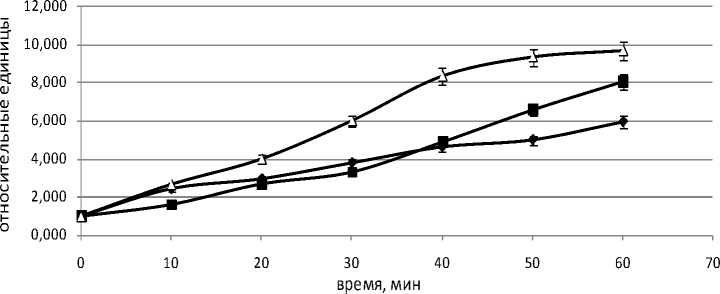
На рисунке 2 представлено изменение скорости продукции желчи для различной длительности подготовительного периода. В группах 1 и 2 скорость выделения желчи остается стабильной в течение всего эксперимента, в то время как в группе 3 после 50 мин она заметно падает.
Для удельной артерио-венозной разницы по кислороду (рис. 3) наиболее стабильными были значения показателя для органов первой группы, а для органов животных второй и третьей групп измеряемые показатели достаточно заметно отклонялись от исходного значения.
—♦— 1 —■— 2 —.J— 3
Рис. 2. Изменение скорости выделения желчи изолированной перфузируемой печенью при различной длительности подготовительного периода (1, 2, 3 – номер экспериментальной группы)
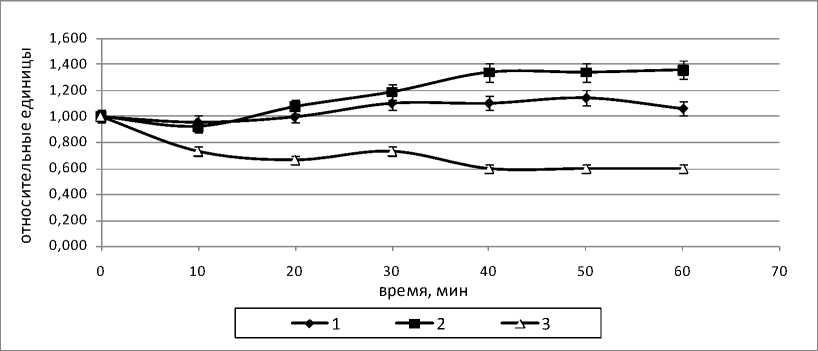
Рис. 3. Динамика удельной артерио-венозной разницы по кислороду для изолированной перфузируемой печени при различной длительности подготовительного периода
(1, 2, 3 – номер экспериментальной группы)
В целом наиболее стабильную по всем показателям динамику наблюдали для органов животных первой группы. Динамика показателей для органов второй и третьей групп была менее стабильной: наблюдались колебания значений исследуемых величин.
Реакция изолированной перфузируемой печени крысы на введение норадреналина
Из таблицы 1 можно видеть, что с момента введения норадреналина уровень давления в воротной вене при перфузии органов животных первой группы начинает отличаться от контроля. В течение 10 мин после введения норадреналина уровень давления в воротной вене для органов группы 2 достоверно ниже, чем в контроле. Начиная с 70 мин, уровень давления в воротной вене для органов группы 2 повышается до контрольных значений и сохраняется таковым до конца перфузии.
Таблица 1
Динамика уровня давления в воротной вене в группах 1 и 2, относительные единицы, в системе изолированной перфузируемой печени крыс при введении норадреналина
|
Время, мин |
30 |
40 |
50 |
60 |
70 |
80 |
90 |
100 |
110 |
120 |
|
Группа 1 |
1,000 ±0,050 |
0,982 ±0,049 |
0,994 ±0,050 |
1,113 ±0,056 |
1,029 ±0,051 |
1,064 ±0,053 |
1,072 ±0,054 |
1,119 ±0,056 |
1,164 ±0,058 |
1,226 ±0,061 |
|
Группа 2 |
1,000 ±0,050 |
0,907 ±0,045 |
0,837 ±0,042 |
0,728 ±0,036* |
0,745 ±0,037* |
1,125 ±0,056 |
0,983 ±0,049 |
0,921 ±0,046 |
0,881 ±0,044* |
0,854 ±0,043* |
* Обозначает P<0,05 в сравнении со значениями для группы 1.
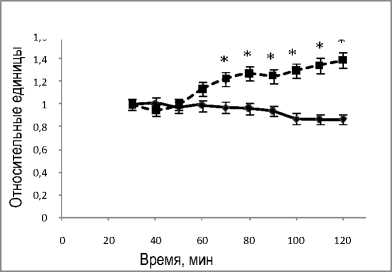
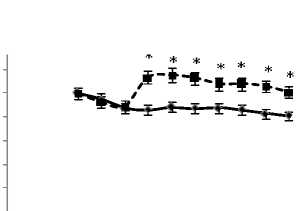
На рисунке 4 представлены изменения скоростей потребления кислорода и выделения углекислого газа, а также отношения скорости выделения углекислого газа к скорости потребления кислорода. Как видно, в группе 1 как скорость потребления кислорода, так и скорость выделения углекислого газа увеличиваются, а в группе 2 уровни этих показателей остаются неизменными. Коэффициент дыхания (рис. 4,в) и в 1, и во 2 группах остается неизменным до последних 20 мин перфузии, после чего в группе 1 заметно понижается, оставаясь прежним при добавлении норадреналина.
0 20 40 60 80 100 120
В емя, мин
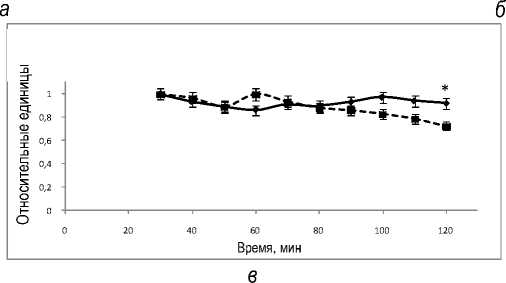
Рис. 4. Изменения скорости потребления кислорода (а) выделения углекислого газа (б) и отношения скорости выделения углекислого газа к скорости потребления кислорода (в) в группах 1 (пунктирная кривая) и 2 (сплошная кривая) в системе изолированной перфузируемой печени крыс при введении норадреналина.* обозначает P<0,05 в сравнении со значениями для группы 1
На рисунке 5 представлена динамика уровней лактата, пирувата и отношения лактат/пируват в течение перфузии. Уровень лактата снижается в течение перфузии, в то время как содержание пирувата в оттекающей от органа жидкости остается постоянным. Отношение содержаний лактата и пирувата (рис. 5,в) уменьшается так же, как и снижается уровень лактата, что согласуется с данными рисунка 4,а.
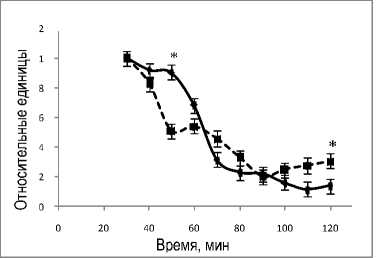
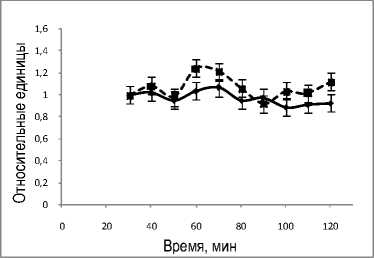
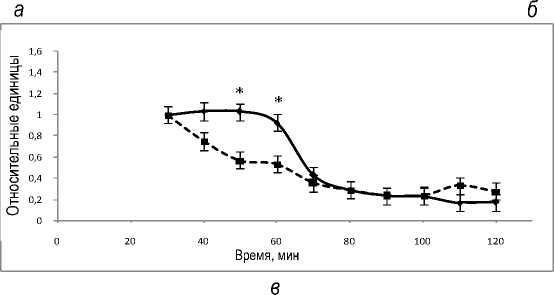
Рис. 5. Динамика уровней лактата (а), пирувата (б) и отношения лактат/пируват (в) в группах 1 (пунктирная кривая) и 2 (сплошная кривая) в системе изолированной перфузируемой печени крыс при введении норадреналина. *обозначает P<0,05 в сравнении со значениями для группы 1
Влияние молочной кислоты на показатели жизнедеятельности изолированной перфузируемой печени крысы
На рисунке 6, а представлена динамика содержаний молочной и пировиноградной кислот в венозной ветви циркуляции. В контроле – перфузия без введения пирувата и лактата в среду.
Из рис. 6, б следует, что введение молочной кислоты приводит к повышению содержания глюкозы, в то время как в контроле содержание глюкозы относительно стабильно. Увеличение содержания глюкозы в этих условиях, возможно, означает участие лактата в метаболическом пути глюконеогенеза.
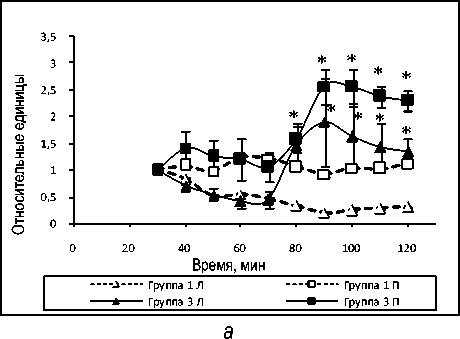
Рис.6. Динамика содержаний молочной и пировиноградной кислот (а) и глюкозы (б) в венозной ветви циркуляции при перфузии изолированной печени крысы: * обозначает P<0,05 в сравнении со значениями для группы 1
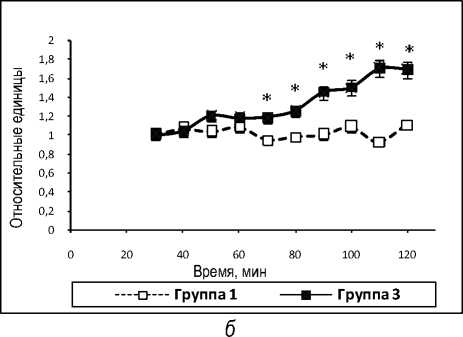
Исходя из данных, представленных в таблице 1, можно считать, что изолированный орган адекватно реагирует на введение норадреналина повышением уровня давления в воротной вене. Полученные данные также могут свидетельствовать о том, что в системе изолированной перфузируемой печени идет нормальный кислородный метаболизм.
По полученным данным можно судить также о том, что изолированная перфузируемая печень может продолжать выполнять одну из своих уникальных метаболических функций – превращение лактата в глюкозу.
Представленные данные свидетельствуют о том, что во время подъема содержания молочной кислоты и пирувата скорость прироста содержания молочной кислоты меньше скорости прироста пировиноградной (табл. 2). Это может означать, что активируется не только кислородный метаболизм, например, цикл Кребса, дыхание в митохондриях, но и другой путь метаболизма лактата, присущий интактной печени, глюконеогенез.
Таблица 2
Динамика скорости изменения содержаний лактата и пирувата в группах 1 и 3, относительные единицы, в системе изолированной перфузируемой печени крыс при введении лактата
|
Время, мин |
30 |
40 |
50 |
60 |
70 |
80 |
90 |
100 |
110 |
120 |
|
Лактат |
||||||||||
|
Группа 1 |
1,000 ±0,050 |
-1,843 ±0,092 |
0,188 ±0,009 |
0,502 ±0,025 |
0,751 ±0,038 |
0,751 ±0,038 |
-0,260 ±0,013 |
-0,182 ±0,009 |
-0,177 ±0,009 |
-0,184 ±0,009 |
|
Группа 3 |
1,000 ±0,050 |
-1,448 ±0,072 |
-0,704 ±0,035* |
-0,128 ±0,006* |
6,472 ±0,324* |
3,864 ±0,193* |
-2,080 ±0,104* |
-1,544 ±0,077* |
-1,160 ±0,058* |
-0,584 ±0,029* |
|
Пируват |
||||||||||
|
Группа 1 |
1,000 ±0,050 |
-1,259 ±0,063 |
3,250 ±0,163 |
-0,345 ±0,017 |
-1,905 ±0,095 |
-1,741 ±0,087 |
1,405 ±0,070 |
-0,241 ±0,012 |
1,310 ±0,066 |
-0,655 ±0,033 |
|
Группа 3 |
1,000 ±0,050 |
-1,360 ±0,068 |
-1,800 ±0,090* |
-0,200 ±0,010* |
6,740 ±0,337* |
16,700 ±0,835* |
-0,360 ±0,018* |
-3,200 ±0,160* |
-2,320 ±0,116* |
-1,320 ±0,066* |
* Обозначает P<0,05 в сравнении со значениями для группы 1.
Выводы
Таким образом, результаты исследования свидетельствуют о том, что норадреналин нормализует физиологическую и метаболическую активности изолированной печени при перфузии и может быть использован для стабилизации параметров гомеостаза органа.
Протекание глюконеогенеза в изолированной перфузируемой печени наряду с обычным аэробным метаболизмом углеводов может свидетельствовать о том, что функционирование органа в условиях искусственного гомеостаза протекает адекватно условиям in vivo. Нагрузка молочной кислотой, в свою очередь, может служить удобным и надежным тестом на адекватность перфузии.
Результаты исследования свидетельствуют также о том, что жизнеспособность изолированной перфузируемой печени зависит от длительности подготовительного периода, во время которого орган находится в состоянии гипоксии. Чем короче период между выделением органа и подсоединением к системе искусственного жизнеобеспечения, тем стабильнее его функционирование.
