Влияние засоления на некоторые показатели водного обмена у растений ячменя (Hordeum vulgare L.)
")
Автор: Таскина К.Б., Казнина Н.М.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Среда, стресс, адаптации
Статья в выпуске: 5 т.60, 2025 года.
Бесплатный доступ
Одна из основных причин снижения урожая хлебных злаков на засоленных почвах - нарушение водного и ионного баланса клеток, поэтому обязательным условием успешного роста и развития растений при засолении служит их способность поддерживать необходимый уровень водного обмена, что особенно важно на начальных этапах онтогенеза, когда растения наиболее чувствительны к воздействию стресс-факторов. Вследствие этого изучение показателей водного режима у злаков весьма актуально, однако необходимо отметить относительно низкую изученность вопросов, связанных с изменением водного обмена растений при засолении. В настоящей работе мы впервые показали, что уже у 10-суточных проростков ячменя при засолении активизируются адаптивные реакции, направленные на сохранение водного режима растений. Целью исследования было изучение влияния умеренного и сильного засоления на некоторые показатели водного обмена растений ячменя. Опыты проводили в 2023-2024 годах в Институте биологии Карельского научного центра РАН (ИБ КарНЦ РАН). Использовали ячмень (Hordeum vulgare L.) ярового сорта Нур, рекомендованный для выращивания в Нечерноземной зоне и характеризующийся пластичностью к изменению почвенных условий. В лабораторных условиях семена проращивали в темноте при 25 °С в чашках Петри (ГОСТ 12038-84. М., 2011). Наклюнувшиеся семена по 10 шт. помещали в сосуды объемом 0,5 л с крышками, в которых были проделаны отверстия. В контрольном варианте в сосуды наливали 1/2 питательный раствор Хогланда-Арнона, в опытных вариантах к питательному раствору добавляли NaCl (х.ч.) в концентрациях 100 и 150 мМ. Растения контрольного и опытных вариантов выращивали в контролируемых условиях среды под светоустановкой в течение 7 сут при температуре 20-22 °С, влажности 60-70 %, ФАР 100 мкмоль/(м2·с) и 14-часовом фотопериоде. По завершении опыта определяли содержание ионов Na+ и Cl- в побегах, сырую биомассу побегов, оводненность тканей побегов (ОВ), относительное содержание воды (RWC) в листе, общую потерю воды листом (LWL), состояние устьичного аппарата (число устьиц и диаметр устьичной щели), устьичную проводимость и интенсивность транспирации. Содержание Na+ определяли атомно-абсорбционным методом с использованием спектрофотометра АА-7000 («Shimadzu», Япония); Cl- - потенциометрическим методом с помощью потенциометра Анион 4100 («Инфраспак-Аналит НПП», Россия). Подсчет числа устьиц на абаксиальной стороне листа, измерение площади и диаметра устьичной щели осуществляли методом отпечатков с использованием светового микроскопа Микмед 2 («Ломо», Россия) и окуляр-микрометра. Устьичную проводимость и интенсивность транспирации измеряли с помощью установки для исследования CO2-газообмена и водяных паров HCM-1000 («Heinz Walz GmbH», Германия). Выявлено, что при умеренном и сильном засолении содержание ионов Na+ в побегах ячменя увеличивалось соответственно более чем в 150 и 170 раз по сравнению с контролем, а ионов Cl- - в 8 и 9 раз. При этом в опытных вариантах растения имели более низкое по сравнению с контролем RWC в листьях, но выраженного их завядания и потери тургора не наблюдалось. Оводненность тканей также сохранялась на необходимом уровне, что во многом обеспечивалось увеличением в условиях засоления водоудерживающей способности листьев, частичным закрытием устьиц и замедлением скорости транспирации. В частности, показатель LWL, характеризующий потерю воды листом, оказался при умеренном и высоком засолении меньше соответственно на 37 и 42 %, чем в контроле, что свидетельствует об увеличении водоудерживающей способности. Устьичная проводимость была снижена более чем на 40 % по сравнению с контролем, а скорость транспирации на 40 и 60 % в зависимости от концентрации соли, что свидетельствовало о частичном закрытии устьичной щели в стрессовых условиях. Обнаруженное у опытных растений снижение накопления сырой биомассы побега, очевидно, было следствием торможения роста как адаптивной реакции на стрессовое воздействие. Таким образом, уже на ранних стадиях развития ячменя сорта Нур умеренное и сильное засоление приводит к целому ряду физиолого-биохимических изменений, которые способствуют сохранению водного режима растений, что, по крайней мере отчасти, объясняет высокую солеустойчивость культуры.
Натрий-хлоридное засоление, содержание ионов, оводненность, относительное содержание воды, водоудерживающая способность, устьица, интенсивность транспирации
Короткий адрес: https://sciup.org/142247702
IDR: 142247702 | УДК: 633.16:581.1:58.04 | DOI: 10.15389/agrobiology.2025.5.866rus
Effect of salinization on some parameters of water status in barley (Hordeum vulgare L.) plants
Hordeum vulgare L.) Nur variety, recommended for cultivation in the Non-Chernozem zone and characterized by its flexibility in changing soil conditions. Under laboratory conditions, the seeds were germinated in the dark at 25 °C in Petri dishes (GOST 12038-84. Moscow, 2011). Ten hatched seeds were placed in 0.5-liter vessels with perforated lids. In the control variant, the vessels were filled with 1/2 Hoagland-Arnon nutrient solution; in the experimental variants, NaCl was added to the nutrient solution at concentrations of 100 and 150 mM. The plants of the control and experimental variants were grown in controlled environmental conditions under a light installation for 7 days at 20-22 °C, humidity of 60-70 %, PAR of 100 μmol/(m2·s) and a 14-hour photoperiod. Upon completion of the experiment, the content of Na+ and Cl- ions in shoots, shoot fresh biomass, shoot tissue water content (OB), leaf relative water content (RWC), total leaf water loss (LWL), stomatal apparatus condition (number of stomata, area and diameter of the stomatal aperture), stomatal conductance and transpiration rate were determined. The Na+ content was determined by the atomic absorption method using an AA-7000 spectrophotometer (Shimadzu, Japan); the Cl- content was measured potentiometrically using an Anion 4100 potentiometer (Infraspak-Analit NPP, Russia). The number of stomata on the leaf abaxial side and stomatal aperture diameter were measured using the indentation method on the light microscope Mikmed 2 (Lomo, Russia) and an ocular micrometer. Stomatal conductance and transpiration rate were measured using an HCM-1000 CO2 gas exchange and water vapor measurement system (Heinz Walz GmbH, Germany). It was found that under moderate and severe salinity, the Na+ ions content in barley shoots increased by more than 150 and 170 times, respectively, compared to the control, while Cl- ions content increased by 8 and 9 times. Moreover, in the experimental variants, plants had a lower RWC in the leaves compared to the control, but no significant wilting or loss of turgor was observed. Tissue water content was also maintained at the required level, largely due to an increase in leaf water-holding capacity under salinity conditions, partial closure of stomata, and a slower transpiration rate. Specifically, the LWL, which characterizes leaf water loss, was 37 % and 42 % lower under moderate and high salinity than in the control, respectively, indicating an increase in water-holding capacity. Stomatal conductance was reduced by more than 40 % compared to the control, and the transpiration rate by 40 % and 60 %, depending on the salt concentration, indicating partial closure of stomata under stressful conditions. The decrease in shoot wet biomass accumulation in the experimental plants was apparently a consequence of growth inhibition as an adaptive response to stress. Thus, already in the early stages of development of the Nur barley variety, moderate and strong salinization leads to a number of physiological and biochemical changes that contribute to the maintenance of the plant's water regime, which, at least in part, explains the high salt tolerance of this crop.
Текст научной статьи Влияние засоления на некоторые показатели водного обмена у растений ячменя (Hordeum vulgare L.)
Засоление почв — один из наиболее опасных факторов окружающей среды, отрицательно влияющих на урожай хлебных злаков. Во многом это связано с нарушением водного и ионного баланса растений из-за развития осмотического стресса и изменения содержания необходимых ионов, что негативно отражается на всех физиологических процессах и, в конечном итоге, на продуктивности (1-3). Сохранение в клетках и тканях необходимого водного баланса чрезвычайно важно для успешного роста и развития растений в условиях засоления.
Показано, что у злаков при засолении наблюдаются симптомы обезвоживания, в частности уменьшается оводненность тканей корня и побега (4), снижается относительное содержание воды в листьях и нарушается тургор клеток мезофилла (5-7), тормозится накопление сырой биомассы (8, 9). Одной из причин указанных изменений становится уменьшение поглощения воды корнем и, как следствие, снижение водного потенциала клеток (10). Однако некоторые виды (сорта, генотипы) способны довольно успешно произрастать в условиях умеренного засоления за счет функционирования механизмов адаптации, действующих на разных уровнях организации: активизации транспортных белков плазмалеммы для восстановления баланса ионов (11), усиления активности аквапоринов с целью улучшения проведения воды тканями (12), повышения содержания абсцизовой, жасмоновой, гибберилиновой кислот, накопления осмолитов в цитоплазме (13). Помимо этого, важным и эффективным механизмом сохранения оводненности тканей в условиях засоления служит закрытие устьиц и уменьшение скорости транспирации, что способствует поддержанию необходимого количества воды в тканях, а также ограничивает поступление избытка ионов солей в подземные и надземные органы (13).
Судя по данным литературы, ячмень относится к наиболее солеустойчивым зерновым культурам. Помимо перечисленных выше механизмов адаптации, у ячменя, в отличие от большинства других видов злаков, основную роль в транспорте воды к сосудам ксилемы играет симпластиче-ский путь, позволяющий в неблагоприятных условиях среды более эффективно (по сравнению с апопластическим) осуществлять контроль за поступлением воды в клетки, усиливая водопоглотительную способность корней (14). При этом разные сорта (генотипы) ячменя могут заметно различаться по солеустойчивости, что во многом связано с различиями в накоплении ионов (15, 16), способности поддерживать необходимую устьичную проводимость (17), а также с разной способностью к осморегуляции (16).
Однако, несмотря на важность водного обмена в жизнедеятельности растений, воздействие засоления на этот процесс у злаков изучено в гораздо меньшей степени, чем, например, на фотосинтез, рост или развитие.
В настоящей работе мы впервые показали, что уже у 10-суточных проростков ячменя при засолении активизируются адаптивные реакции, направленные на сохранение водного режима растений.
Целью исследования было изучение влияния умеренного и сильного засоления на некоторые показатели водного обмена растений ячменя.
Методика. Опыты проводили в 2023-2024 годах в Институте биологии Карельского научного центра РАН (ИБ КарНЦ РАН). Использовали ячмень ( Hordeum vulgare L.) ярового сорта Нур, рекомендованный для выращивания в Нечерноземной зоне и характеризующийся пластичностью к изменению почвенных условий (18). Семена были предоставлены ФГБНУ Московский НИИ сельского хозяйства Немчиновка.
В лабораторных условиях семена проращивали в темноте при 25 °С в чашках Петри (ГОСТ 12038-84. М., 2011). Наклюнувшиеся семена по 10 шт. помещали в сосуды объемом 0,5 л с крышками, в которых были проде- ланы отверстия. В контрольном варианте в сосуды наливали 1/2 питательный раствор Хогланда-Арнона, в опытных вариантах к питательному раствору добавляли химически чистого NaCl в концентрациях 100 и 150 мМ (концентрации, при которых у растений наблюдаются морфологические изменения, но не происходит их гибели, были выбраны на основании предварительных опытов). Растения контрольного и опытных вариантов выращивали в контролируемых условиях среды под светоустановкой (сборка ИБ КарНЦ РАН) в течение 7 сут при температуре 20-22 °С, влажности 60-70 %, ФАР 100 мкмоль/(м2 • с) и 14-часовом фотопериоде.
По завершении опыта определяли содержание ионов Na + и Cl - в побегах, оценивали влияние двух концентраций хлорида натрия на овод-ненность тканей побега (ОВ), относительное содержание воды (RWC, relative water content) в листе, общую потерю воды листом (LWL, leaf water loss), состояние устьичного аппарата (число устьиц и диаметр устьичной щели), устьичную проводимость и интенсивность транспирации. Содержание Na+ определяли атомно - абсорбционным методом с использованием спектрофотометра АА - 7000 («Shimadzu», Япония); Cl - — потенциометрическим методом с помощью потенциометра Анион 4100 («Инфраспак-Аналит НПП», Россия).
Накопление сырой биомассы оценивали по общепринятой методике (19). Оводненность тканей вычисляли по формуле: ОВ = ((А - В)/А) х 100 %, где A — сырая масса, мг, B — сухая масса, мг (20). Для расчета относительного водного содержания использовали формулу RWC = (А - В)/(Т - B) х 100 %, где А — сырая масса, мг, В — сухая масса, мг, Т — масса листа в тургис-центном состоянии, мг (21). Общую потерю воды листом определяли по формуле: LWL = (А - Б)/(А - В) х 100 %, где А — сырая масса, мг, Б — сырая масса после завядания, мг, B — сухая масса, мг (6). Подсчет числа устьиц на абаксиальной стороне листа, измерение площади и диаметра устьичной щели осуществляли методом отпечатков с использованием светового микроскопа Микмед 2 («Ломо», Россия) и окуляр - микрометра (22). Устьичную проводимость и интенсивность транспирации измеряли с помощью установки для исследования CO 2 -газообмена и водяных паров HCM-1000 («Heinz Walz GmbH», Германия).
Каждый вариант опыта включал 3 повторности (сосуда). Содержание ионов для каждого варианта опыта определяли в смешанной пробе, состоящей из 30 растений (навеска 0,5 г для определения Cl - и 0,2 г для Na + ). Измерения проводили в 4 аналитических повторностях ( n = 4).
При определении сырой биомассы побега и изучении показателей водного обмена биологическая повторность составляла 10 растений в каждом варианте опыта. Опыт повторяли трижды ( n = 30). Для подсчета числа и размеров устьиц в каждом варианте опыта анализировали по 9 листьев в 10 полях зрения. Опыт повторяли трижды ( n = 270). Устьичную проводимость и скорости транспирации измеряли у 6 растений в каждом варианте опыта. Опыт повторяли трижды ( n = 18).
Статистическую обработку выполняли с использованием пакета программ Microsoft Office Excel 2010 («Microsoft Corp.», США). В таблицах отражены средние значения ( M) по трем независимым опытам и стандартные ошибки средних (±SEM). Достоверность различий между вариантами опытов оценивали с помощью t -критерия Cтьюдента (при p < 0,05).
Результаты. После 7 сут воздействия хлорида натрия в умеренной (100 мМ) и высокой (150 мМ) концентрациях содержание Na+ в побегах ячменя увеличивалось соответственно более чем в 150 и 170 раз (р < 0,05) по сравнению с контролем, Cl - — соответственно в 8 и 9 раз (p < 0,05) 868
(табл. 1).
1. Содержание ионов Na+ и Cl - в побеге проростков ячменя ( Hordeum vulgare L.) ярового сорта Нур ( n = 4, M ±SEM; лабораторный опыт, 2024 год)
Увеличение содержания ионов в растениях отрицательно сказалось на накоплении ими сырой биомассы. Так, при умеренной концентрации NaCl (100 мМ) сырая биомасса побега была на 32 % меньше (p < 0,05), чем у растений контрольного варианта, а при высокой концентрации (150 мМ) — на 49 % (p < 0,05) (табл. 2). При этом значимо снижался и показатель RWC.
2. Сырая биомасса побега и показатели водного обмена у проростков ячменя ( Hordeum vulgare L.) ярового сорта Нур в зависимости от содержания в среде NaCl ( n = 30, M ±SEM; лабораторный опыт, 2023 год)
|
Вариант опыта |
|
|
контроль 100 мМ NaCl 1 150 мМ NaCl |
|
Важным показателем устойчивости растений к засолению служит водоудерживающая способность листьев, которую можно оценить на основании определения общей потери воды листом: чем меньше LWL, тем выше его водоудерживающая способность (23). В наших опытах в условиях умеренного и сильного засоления LWL оказалась меньше соответственно на 37 и 42 % по сравнению с контролем, что свидетельствует об увеличении водоудерживающей способности (p < 0,05) (см. табл. 2). Оводненность тканей побега в условиях засоления несколько снижалась по сравнению с оптимальными условиями роста, но в целом оставалась на необходимом растению уровне (см. табл. 2).
3. Показатели состояния устьичного аппарата у проростков ячменя ( Hordeum vulgare L.) ярового сорта Нур в зависимости от содержания в среде NaCl
( n = 270, M ±SEM; лабораторный опыт, 2023 год)
|
Показатель |
Вариант опыта |
||
|
контроль |
100 мМ NaCl 1 |
150 мМ NaCl |
|
|
Число устьиц, шт/мм2 328,9±3,2c Диаметр устьичной щели, мкм 8,61±0,06a a, b, c Разные латинские буквы означают статистически ными вариантами при p ≤ 0,05 ( t -критерий Стьюдента). |
358,4±4,4b 483,9±11,6a 8,11±0,07b 7,82±0,08c значимые различия между контрольным и опыт- |
||
Анализ состояния устьичного аппарата показал, что в условиях засоления у растений формируется большее число устьиц (p < 0,05) (табл. 3). Так, при умеренной концентрации NaCl число устьиц на нижней стороне листа было на 9 % больше, чем в контрольном варианте, а при высокой концентрации — на 47 % (p < 0,05). При этом уменьшались диаметр устьичной щели (на 6 и 9 %) (p < 0,05) (см. табл. 3), а также устьичная проводимость (более чем на 40 % по сравнению с контролем) (p < 0,05) (рис., А), что свидетельствовало о частичном закрытии устьиц. Уменьшение вследствие этого скорости транспирации, наблюдаемое в большей степени при концентрации 150 мМ NaCl (p < 0,05), очевидно, было направлено на уменьшение потери воды растением.
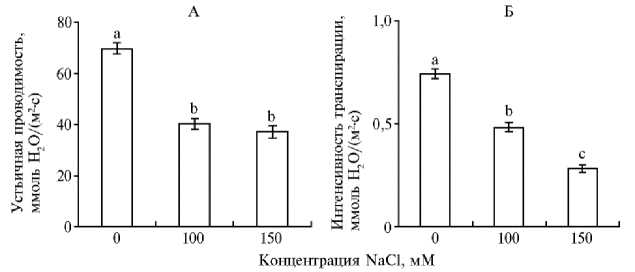
Устьичная проводимость (А) и интенсивность транспирации (Б) проростков ячменя ( Hordeum vulgare L.) ярового сорта Нур в зависимости от содержания в среде NaCl ( n = 18, M ±SEM; лабораторный опыт, 2023 год).
Известно, что на засоленных почвах злаки испытывают сильный водный дефицит вследствие нарушения водного и ионного баланса (11, 24). Повышение содержания хлорида натрия в почве снижает водный потенциал и повышает осмотическое давление раствора в прикорневой зоне, что препятствует всасыванию и транспортировке воды и приводит к обезвоживанию тканей (3). Кроме того, высокие концентрации Na+ и Cl - в растениях вызывают дисбаланс других необходимых для жизнедеятельности растений ионов. Все это приводит к снижению урожайности злаков и ухудшению качества продукции (2, 11).
В настоящем исследовании при обеих изученных концентрациях NaCl значительно возрастало содержание ионов Na+ и Cl - в побегах, что отрицательно сказалось на некоторых показателях водного обмена. В частности, в опытных вариантах растения отличались от контроля меньшим относительным содержанием воды в листьях и более низкой сырой биомассой побега. Вместе с тем выраженного визуального завядания листьев и потери тургора не наблюдалось ни в одном из вариантов, что, вероятно, связано с отсутствием сильного снижения оводненности тканей. Аналогичные результаты были получены при воздействии NaCl в концентрации 100 мМ на проростки пшеницы (25), в концентрации 120 мМ — на растения ячменя сорта Султан (26) и в концентрации 150 мМ — на некоторые генотипы сорго (27). Способность растений сохранять необходимую оводненность в условиях засоления обеспечивается наличием целого ряда механизмов со-леустойчивости. Один из важнейших — увеличение водоудерживающей способности клеток за счет синтеза осмолитов, в том числе пролина, аминокислот, cахаров (11). В проведенных нами исследованиях у ячменя с увеличением степени засоления снижалась общая потеря воды листом, что свидетельствовало о повышении водоудерживающей способности листьев.
Хорошо известен механизм солеустойчивости растений, который заключается в закрытии устьиц и замедлении скорости транспирации (28, 29). По мере увеличения степени засоления и развития водного дефицита снижается водный потенциал клеток мезофилла, возрастает концентрация АБК, что приводит к инактивации Н - помпы плазмалеммы замыкающих клеток. При этом активируются выходные К+-каналы, что сопровождается оттоком К+ из замыкающих клеток и полным или частичным закрытием устьиц. Это приводит к уменьшению интенсивности транспирации, что и обеспечивает сохранение оводненности тканей. В частности, у растений ячменя сорта Прерия при 100 мМ хлорида натрия (30), у сорго сорта Порта 870
Негро при 120 мМ (31), а у манильской травы ( Zoysia matrella (L.) Merr.) при 300 мМ соли (32) отмечалось заметное уменьшение скорости транспирации. По мнению авторов, это обеспечивало лучшую адаптацию растений к засолению. В нашем исследовании скорость транспирации также снижалась в соответствии с концентрационной зависимостью. При этом уменьшался диаметр устьичной щели, свидетельствуя о частичном закрытии устьиц. Важно отметить, что у растений в опытных вариантах на нижней поверхности листа было сформировано большее число устьиц, чем в контроле. Данных о влиянии засоления на формирование устьичного аппарата в известной нам литературе крайне мало. Однако их анализ позволяет предположить, что умеренный стресс приводит к увеличению числа и плотности устьиц, обеспечивая лучшую устьичную проводимость (33, 34). В условиях сильного засоления эти показатели снижаются, что направлено на оптимизацию эффективности использования воды (25).
Таким образом, как при умеренной (100 мМ), так и при высокой (150 мМ) концентрации хлорида натрия у растений ячменя сорта Нур активизируются адаптивные реакции, направленные на сохранение водного режима растений: устьица частично закрываются, снижается скорость транспирации, увеличивается число устьиц, возрастает водоудерживающая способность листьев. Это позволяет растениям сохранить оводненность тканей и тургор клеток листа на необходимом для нормальной жизнедеятельности уровне. Снижение в этих условиях сырой биомассы побега может быть следствием некоторого замедления роста растений, то есть адаптационной реакции на стрессовое воздействие, и позволяет растению экономить энергетические запасы, направляя их на поддержание энергетически затратных процессов. В целом, на основании проведенных исследований можно сделать вывод, что у 10-суточных проростков ячменя сорта Нур при умеренном и сильном засолении активизируются адаптивные реакции, направленные на сохранение водного режима. Способность растений уже на начальных этапах роста поддерживать водный обмен на высоком уровне в условиях засоления во многом обеспечивает их высокую солеустойчивость.
ФГБУН Институт биологии
