Внутрипопуляционная генетическая дифференциация быков холмогорской породы Республики Коми по микросателлитам

Автор: Матюков В.С., Жариков Я.А., Николаев С.В.
Журнал: Известия Коми научного центра УрО РАН @izvestia-komisc
Рубрика: Растениеводство
Статья в выпуске: 1 (77), 2025 года.
Бесплатный доступ
Исследовали полиморфизм микросателлитов у 85 быков- доноров семени следующих пород и типов: холмогорской (38 гол.), голштинской (10), печорского типа холмогорской породы (17), и кроссированных печороского типа с классической холмогорской породой (20 гол.). Генотипировали особей по 15 локусам микросателлитов. Наибольшее число специфических аллелей выявили в группе быков холмогорской породы. Максимальную генетическую дистанцию установили между печоро-холмогорскими быками и быками голштинской породы (DN 0,237, FST 0,045). Подразделение всего массива на два кластера в программе Strukture подтвердило высокую генетическую дифференциацию кроссированных и чистолинейных быков печорского типа холмогорской породы с голштинской. Генетическое различие голштинской породы с классической холмогорской было ниже. Средняя вероятность членства в первом кластере быков голштинской породы составила 0,107±0,039, во втором - 0,893±0,039, быков печорского типа холмогорской породы соответственно 0,828±0,052 и 0,172±0,052, печоро-холмогорских быков - 0,860±0,019 и 0,140±0,019, быков классической холмогорской породы соответственно 0,633±0,046 и 0,367±0,046. Сведения о числе эффективных аллелей на локус, гетерозиготности и индексах фиксации в группах животных разной генеалогии и породности не дали значимой информации об их внутрипопуляционной генетической структуре и дифференциации, которую можно было бы использовать в практической работе по поддержанию генетического разнообразия и воспроизводству сохраненного генофонда породы. Кластерный анализ при k=3 и k=4 позволил выявить и детализировать структуру и степень генетического различия между группами и, таким образом, получить дополнительную информацию для планирования и реализации генофондосохранных мероприятий.
Холмогорская порода, печорский тип, кластер, вероятность, генетическая дифференциация, микросателлиты
Короткий адрес: https://sciup.org/149147832
IDR: 149147832 | УДК: 575.22+599.735.3 | DOI: 10.19110/1994-5655-2025-1-75-82
Intrapopulation genetic differentiation of bulls of the Kholmogory breed in the Komi Republic by microsatellites
The polymorphism of microsatellites was studied in 85 bulls - semen donors of the following breeds and types: the Kholmogory breed (38), the Holstein breed (10), the Pechora type of the Kholmogory breed (17), and the type obtained through crossing the Pechora type with the typical Kholmogory breed (20). Individuals were genotyped by 15 microsatellite loci. The largest number of specific alleles was revealed in the group of the Kholmogory breed bulls. The maximum genetic distance was identified between the Pechora-Kholmogory bulls and the Holstein bulls (DN 0.237, FST 0.045). The division of the entire array into two clusters in the program Strukture confirmed the high genetic differentiation between the crossed and purebred Pechora type bulls of the Kholmogory breed and the Holstein breed. The genetic difference between the Holstein and the typical Kholmogory breed was as high. The average probability of membership in the first cluster was 0.107±0.039, in the second cluster - 0.893±0.039 for Holstein bulls, 0.828±0.052 and 0.172±0.052 for Pechora type bulls of the Kholmogory breed, 0.860±0.019 and 0.140±0.019 for Pechora-Kholmogory bulls, and 0.633±0.046 and 0.367±0.046 for bulls of the typical Kholmogory breed, respectively. Information on number of effective alleles per locus, heterozygosity and fixation indices in animal groups of different genealogy and breeds did not provide any reliable and significant information on their intrapopulation genetic structure and differentiation that could be used in practical work to maintain the genetic diversity and reproduce the conserved gene pool of the breed. The cluster analysis at k=3 and k=4 allowed to identify and detail the structure and degree of genetic difference between groups and, thus, obtain additional information for planning and implementing gene pool conservation measures.
Текст научной статьи Внутрипопуляционная генетическая дифференциация быков холмогорской породы Республики Коми по микросателлитам
Выживание человека в неблагоприятных природно-климатических и социально-экономических условиях всегда было связано с использованием традиционных способов природопользования и разведением аборигенных пород сельскохозяйственных животных. Сохранение исторически приобретенных коренными народами навыков получения животноводческого сырья для производства продуктов питания особенно актуально для территорий рискованного земледелия, к которым, в первую очередь, относятся заселенные человеком пространства высоких широт, в частности Республики Коми. На территории современной Республики Коми с момента ее заселения в оседлом животноводстве разводили аборигенный северный комолый крупный рогатый скот, северную грубошерстную короткотощехвостую овцу и северную упряжную лошадь. Вплоть до 1960-х гг. эти породы можно было встретить в коллективных и индивидуальных хозяйствах населения. Однако со второй половины ХХ в. породное преобразование, механизация и специализация аграрной отрасли привели к практически полной утрате аборигенных генофондов. В молочном скотоводстве северный комолый скот был поглощен и вытеснен холмогорским, а с 1980-х гг. в ходе технологической революции в молочном скотоводстве холмогорский скот вытесняется и поглощается голштинским. По данным инвентаризации 2020 г. по Российской Федерации, общая численность чистопородного холмогорского скота составила всего 1170 гол. [1]. Более половины этого поголовья сохранялось в хозяйствах Республики Коми, расположенных в районах Крайнего Севера и Приполярья с неблагоприятными условиями кормления и содержания животных. Для воспроизводства холмогорского скота на Головном племпредприятии РГУСП «Коми» по племенной работе сохранили банк глубокозаморожен-ного семени чистопородных и помесных быков-производителей холмогорской породы, уничтоженный в большинстве регионов России.
Холмогорский скот в хозяйствах республики по генеалогической структуре, некоторым экстерьерным и хозяйственным признакам отличался от заводской популяции, поскольку был получен на основе местного северного комолого скота. В начале процесса породного преобразования в хозяйствах Крайнего Севера, Приполярья и Заполярья Коми АССР для скрещивания использовали быков-производителей холмогорской породы, выводных из хозяйств соловецкой (Беломоро-Балтийской) популяции, которая была уничтожена во время Великой Отечественной войны. В дальнейшем на базе массива холмогорского скота северных районов Республики Коми был создан и официально признан зональный печорский тип холмогорской породы. В условиях Приполярья и Заполярья животные печорского типа по продуктивности не уступали классической холмогорской породе, характеризовались более продолжительным сроком хозяйственного использования и устойчивостью к ряду заболеваний, в том числе лейкозу крупного рогатого скота. Холмогорская порода в целом при уровне удоев около 5 тыс. кг молока за лак- тацию до 2000-х гг. по продолжительности хозяйственного использования, воспроизводительным способностям, пожизненной молочной продуктивности коров и в расчете на месяц жизни оставалась конкурентоспособной относительно голштинской [2]. В настоящее время в связи с малочисленностью сохранившегося поголовья воспроизводство холмогорского скота требует проведения ряда мероприятий, важнейшими из которых являются генетический мониторинг и поддержание генетического разнообразия [3]. Основными инструментами, с помощью которых оценивают генетическое разнообразие и идентифицируют породную принадлежность животных являются биохимические и молекулярные маркеры [4, 5]. Для решения этих задач наиболее доступными и эффективными маркерами служат микросателлиты [6, 7].
Цель настоящего исследования – изучение микросател-литного профиля быков-доноров сохраненного криокон-сервированного семени и оценка внутрипопуляционного индивидуального и группового генетического разнообразия сохраненной популяции холмогорского скота.
Материалы и методы
Исследовали полиморфизм микросателлитов у быков-доноров криоконсервированного семени, хранящегося в банке племпредприятия РГУСП «Коми» по племенной работе. Данные о генотипах быков по локусам микросателлитов предоставлены для генетического анализа руководством РИСЦ. Генотипирование быков по локусам микросателлитов на коммерческой основе проведено лабораторией ДНК-технологий Всероссийского научно-исследовательского института племенного дела. ДНК для исследования выделяли из криоконсервированного семени быков.
Общая численность выборки генотипированных чистопородных холмогорских быков составила 75 гол., в том числе печорского типа холмогорской породы (П) – 17, холмогорских быков классических линий (Х) – 38 гол., кроссированных печоро-холмогорских (ПХ) – 20. Для межпородного сравнения использовали результаты генотипирования 10 гол. чистопородных голштинских быков (Гл). Результаты генотипирования быков по STR-локусам обработали с помощью программы GenAlEx 6.5 [8]. По разным генеалогическим группам и породам рассчитали следующие параметры: N – число животных в группе; Na – среднее число аллелей на локус; Ne – среднее эффективное число аллелей на локус; Ho – оценка средней наблюдаемой (фактической) гетерозиготности; He – оценка средней ожидаемой гетерозиготности на локус; uHe – несмещенная оценка средней ожидаемой гетерозиготности на локус; FIS – индекс фиксации (инбридинга). Парные генетические дистанции между группами быков рассчитали следующими методами: DN – генетическая дистанция по Nei М.; FST – генетическая дистанция по Wright S. и оценки Fst, G’ST (Nei), G’st(Hed), G’’ST(Hed) Dest , полученные с использованием метода генерации случайных выборок в программе GenAlEx 6.5.
В программе Structurе [9] рассчитали апостериорную вероятность (Q) членства отдельных STR-генотипов в различных кластерах без введения предварительной информации об их классификации по генеалогии. Членство отдельных генотипов (особей) в кластерах вычислили при разном их числе (k=2–4). По каждой группе рассчитали средние вероятности членства особей в k-м кластере начиная с k=2 до k=4. Распределение средних вероятностей по кластерам свели в таблицы и визуализировали графически (рис. 1–3). Для детализации генетических различий особей, отнесенных к различным кластерам при k=3 и k=4, в пределах каждого кластера рассортировали особей по вероятности членства и с порогом вероятности 0,75 сформировали в группы. Затем в программе GenAlEx 6.5 по сформированным группам рассчитали их генетико-популяционные характеристики и генетические дистанции между ними. Статистическую обработку данных и графики выполнили в программе Excel.
Результаты и их обсуждение
Исследование показало, что разные внутрипородные группы быков достоверно не различались и были близ ки по параметрам генетического разнообразия (табл. 1). Хотя недостоверная дифференциация по некоторым показателям прослеживалась. Так, кроссбредные печоро-холмогорские быки при численности выборки в два раза большей, по сравнению с чистопородными голштинскими быками, уступили им по эффективному числу аллелей. В то же время по наблюдаемой гетерозиготности при высоких показателях абсолютного и эффективного числа аллелей на локус голштинские быки уступали остальным группам по уровню наблюдаемой гетерозиготности. Кроме того, в группе голштинских быков наблюдалась редукция гетерозигот, в то время как по группам быков печорского типа, печоро-холмогорским и холмогорским прослежи- вался небольшой избыток гетерозигот. Генетические дистанции между кроссированными печоро-холмогорскими быками и голштинской породой были максимальными.
Несколько ниже по величине была дифференциация с голштинской породой быков печорского типа и чистопородных холмогорских быков классических линий (табл. 2). В целом значения межпородных генетических дистанций были ниже минимальных значений, опубликованных в обзорной работе Холла [10].
При попарном сравнении групп быков разной породности коэффициенты корреляции оценок генетических дистанций, определенных разными методами, были выше 0,9.
Сравнение аллелофондов отдельных групп быков выявило 12 приват-аллелей, в том числе в группе быков
Таблица 1
Table 1
Характеристика групп быков-производителей по генетико-популяционным показателям (Х±sx)
Characteristics of groups of seed bulls by the genetic-population indicators (Х±sx)
|
Группы |
N |
Na |
Ne |
Ho |
He |
uHe |
Fis |
|
Гл |
10 |
5,333±0,422 |
3,813±0,385 |
0,627±0,052 |
0,691±0,037 |
0,727±0,039 |
0,087±0,061 |
|
П |
17 |
5,867±0,435 |
3,747±0,391 |
0,702±0,026 |
0,698±0,026 |
0,719±0,027 |
-0,019±0,041 |
|
ПХ |
20 |
5,867±0,389 |
3,579±0,283 |
0,724±0,030 |
0,698±0,021 |
0,715±0,022 |
-0,040±0,034 |
|
Х |
38 |
6,800±0,490 |
3,814±0,279 |
0,733±0,024 |
0,717±0,021 |
0,727±0,021 |
-0,024±0,026 |
Таблица 2
Генетические дистанции, рассчитанные разными способами при попарном сравнении выборок быков-производителей разной породности
Table 2
Genetic distances calculated using different methods for pairwise comparison between seed bulls of different types and breeds
|
Оценки Группы |
DN |
uDN |
Fst |
Fst’ |
G’st (Nei) |
G’st (Hed) |
G’’st |
Dest |
|
Гл*П |
0,188 |
0,089 |
0,035 |
0,036 |
0,028 |
0,090 |
0,103 |
0,071 |
|
Гл*ПХ |
0,237 |
0,145 |
0,045 |
0,045 |
0,049 |
0,152 |
0,173 |
0,130 |
|
П*ПХ |
0,047 |
0,000 |
0,010 |
0,010 |
-0,008 |
-0,018 |
-0,027 |
-0,019 |
|
Гл*Х |
0,185 |
0,106 |
0,034 |
0,035 |
0,034 |
0,111 |
0,126 |
0,095 |
|
П*Х |
0,079 |
0,026 |
0,015 |
0,016 |
0,005 |
0,030 |
0,033 |
0,024 |
|
ПХ*Х |
0,063 |
0,016 |
0,013 |
0,013 |
0,007 |
0,020 |
0,024 |
0,017 |
Примечание. Полужирным шрифтом выделены достоверные значения генетических дистанций при уровне значимости p ≤ 0,05.
Note. Reliable values of genetic distances at a significance level of p ≤ 0.05 are semi-bolded.
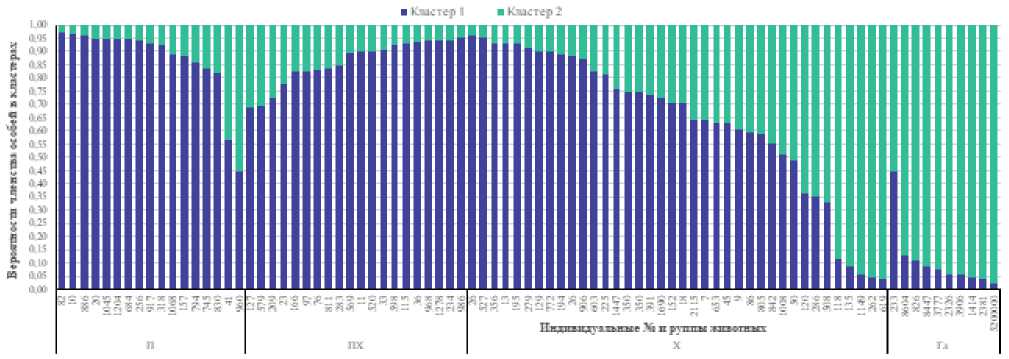
Рисунок 1. Вероятности членства особей в кластерах при k=2.
Figure 1. Membership probabilities of individuals in clusters at k=2.
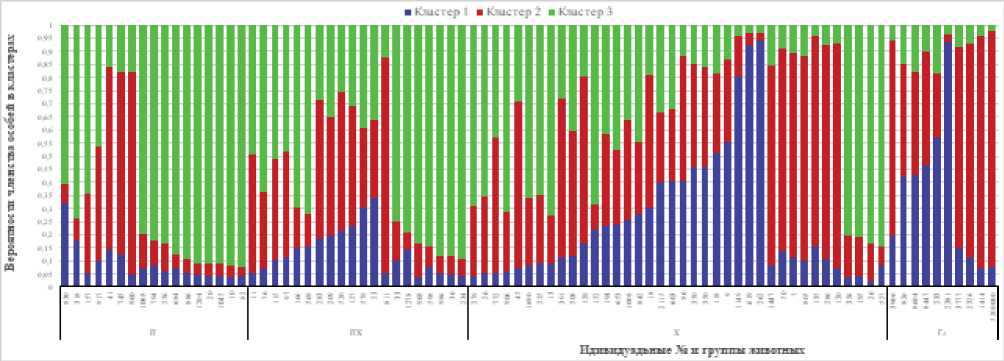
Рисунок 2. Вероятности членства особей в кластерах при k=3.
Figure 2. Membership probabilities of individuals in clusters at k=3.
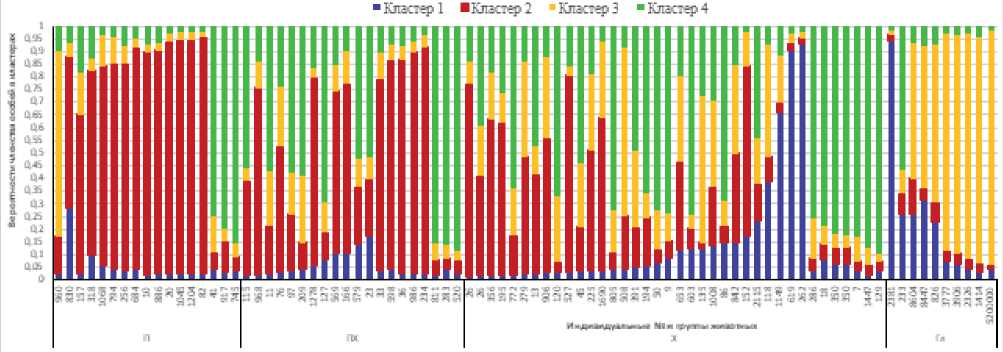
Рисунок 3. Вероятности членства особей в кластерах при k=4.
Figure 3. Membership probabilities of individuals in clusters at k=4.
Таблица 3
Исходные данные для построения диаграммы к рисунку 2
-
Table 3
Initial data to draw diagram in Figure 2
|
Группы быков разной породности |
Кластер |
|
|
1 |
2 |
|
|
Гл 100 |
0,107 |
0,893 |
|
П |
0,828 |
0,172 |
|
ПХ |
0,860 |
0,140 |
|
Х |
0,633 |
0,367 |
Таблица 4
Исходные данные для построения диаграммы к рисунку 3
-
Table 4
Initial data to draw diagram in Figure 3
Анализ вероятности членства быков печорского типа в определенных кластерах при k=2 показал, что у 88,2 % животных вероятность членства в первом кластере была равна или превысила порог 0,75. У двух быков (11,8 %) вероятность членства в первом или втором кластере была
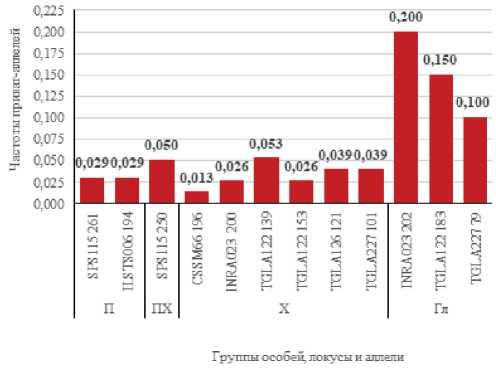
Рисунок 4. Приват–аллели в группах быков.
Figure 4. Private alleles in the bull groups.
Таблица 5
Средняя вероятность принадлежности групп различной породности к кластерам при k=3 и k=4 (Х±sx)
Table 5
A verage affiliation probability of groups of different breeds and types to clusters at k=3 and k=4 (Х±sx)
|
Группа |
N |
k=3 |
k=4 |
|||||
|
1 |
2 |
3 |
1 |
2 |
3 |
4 |
||
|
Гл |
10 |
0,35±0,087 |
0,56±0,095 |
0,09±0,019 |
0,23±0,086 |
0,05±0,012 |
0,62±0,105 |
0,10±0,052 |
|
П |
17 |
0,09±0,030 |
0,22±0,060 |
0,69±0,069 |
0,05±0,015 |
0,65±0,080 |
0,10±0,040 |
0,20±0,071 |
|
ПХ |
20 |
0,13±0,020 |
0,30±0,047 |
0,57±0,055 |
0,06±0,010 |
0,45±0,072 |
0,10±0,015 |
0,39±0,068 |
|
Х |
38 |
0,25±0,039 |
0,39±0,040 |
0,36±0,044 |
0,14±0,036 |
0,24±0,038 |
0,20±0,024 |
0,42±0,049 |
членства в каком-либо кластере не достигала порогового значения. В группе печоро-холмогорских быков из 20 особей пять (25 %) отнесли ко второму кластеру и три (15 %) – к четвертому. Остальные 60 % не достигли порога вероятности членства в одном из кластеров. В выборке холмогорской породы семь особей с вероятностью членства 0,75 и выше отнесли к четвертому кластеру, два – к первому ниже 0,75. По группе печоро-холмогорских быков три особи (15 %) не достигли порогового значения для отнесения их к первому кластеру. Вероятность их членства в кластере сохранялась на уровне 0,688–0,723. У остальных быков вероятность членства в кластерах была равна или превысила установленное пороговое значение. У животных классической холмогорской породы 36,8 % особей с вероятностью 0,75 и выше относились к первому кластеру, 13,2 % – ко второму. Остальные особи не достигли пороговых значений вероятности для отнесения их к одному из кластеров (см. рис. 1).
В выборке голштинской породы у девяти быков (90 %) вероятность членства во втором кластере превысила пороговое значение, у одного быка (№ 233) вероятность отнесения к первому или второму кластеру распределилась примерно поровну.
При подразделении экспериментальной выборки на три кластера в группе печорских быков 10 (58,9 %) особей достигли и превысили пороговое значение вероятности членства в третьем кластере, одна особь отнесена ко второму кластеру (0,6 %).
В группе печоро-холмогорских быков семь (35 %) особей отнесены к третьему кластеру, одна – ко второму (0,5 %). Максимальная вероятность отнесения особей к первому кластеру в этих группах не превышала соответственно 0,321 и 0,342.
По голштинской породе 40 % особей отнесены ко второму кластеру со значением Q ≥ 0,75 и одна особь (10 %) – к первому.
В группе быков классической холмогорской породы величина Q в третьем кластере варьировала по отдельным животным от 0,027 до 0,847, при этом только у четырех особей (10,5 %) значение Q составило 0,75 и выше. У 18,4 % особей вероятность их членства во втором кластере достигла 0,75 и выше, в первом – у трех (7,9 %) (рис. 2, табл. 3). Из этих данных видно, что классическая холмогорская порода занимала промежуточное положение между печорским типом и голштинской породой.
Подразделение массива данных на четыре кластера по существу не изменило интерпретации полученных данных (рис. 3, табл. 4). Десять печорских чистолинейных быков с вероятностью 0,75 и выше отнесли ко второму кластеру и трех – к четвертому. Суммарно кластерная принадлежность определена для 76,5 % особей. В группе голштинских быков 50 % особей отнесено к третьему кластеру и одна особь – к первому. У остальных 40 % особей вероятность и два – ко второму. Суммарно у 11 (28,9 %) быков вероятность членства в одном из кластеров достигла или превысила пороговое значение.
Из представленных в табл. 5 данных видно, что по кластерной структуре аллелофонда микросателлитов при k=3 и k=4 сходство с голштинской породой возрастает от минимума у быков печорского типа до максимума у быков классической холмогорской породы.
Таким образом, по общим параметрам генетической изменчивости достоверных различий между группами не установили (см. табл. 1). Дифференциация холмогорской породы c голштинской по некоторым оценкам была на одном уровне или несколько выше, чем у быков печорского типа холмогорской породы. Между парой П*ПХ генетические дистанции, оцененные разными способами, в большинстве случаев были близки к нулю (см. табл. 2). Редукция гетерозигот в группе чистопородных голштинских быков, вероятно, объясняется инбредностью использованных голштинских производителей. Полученные нами результаты лишь частично согласуются с опубликованными данными Волковой с соавт [11], которые установили, что холмогорская порода, по сравнению с голштинской, обладала более высоким уровнем аллельного разнообразия по STR-локусам: по среднему числу аллелей на локус, количеству информативных аллелей и эффективному числу аллелей. Оценки уровня аллельного разнообразия, гетерозиготности, индексов фиксации, генетической дистанции между печорским типом холмогорской породы, холмогорской с голштинской оказались близкими. Эти сведения не дали практически значимой информации о внутрипопуля-ционной генетической структуре сравниваемых групп, которую можно было бы использовать в практической работе по поддержанию генетического разнообразия и воспроизводству сохраненного генофонда породы.
Планируя эксперимент, мы полагали, что использование для генетического анализа микросателлитов, вследствие их высокого полиморфизма [12] в сочетании с кластерным анализом, позволит детализировать информацию об индивидуальной и групповой структуре, генетической изменчивости и дифференциации внутрипопуляционных субъединиц.
Результаты исследования подтвердили это предположение. Последовательный кластерный анализ генетической структуры популяции показал, что при k=2 уровень дифференциации и консолидированности быков голштинской породы, кроссированных печоро-холмогорских производите- лей и чистолинейных быков печорского типа холмогорской породы был высоким. Консолидированность быков холмогорской породы оказалась гораздо ниже. Высокая консоли-дированность быков печорского типа холмогорской и голштинской пород была подтверждена и при подразделении массива на три и четыре кластера. При k=3 в генетической структуре печорского типа превалировал компонент третьего и отчасти второго кластеров, последний был основным у голштинской породы. Суммарная доля генетического компонента второго и первого кластеров у голштинской породы была почти в три раза выше, чем у печорского типа – 0,91 к 0,31, у печоро-холмогорских быков – 0,43, у классической холмогорской породы – 0,64 (см. табл. 3). Таким образом, классическая холмогорская порода заняла промежуточное положение между печорским типом и голштинской породой. С увеличением числа кластеров величины и соотношения вероятностей членства особей в соответствующих кластерах менялись, детализировалась генетическая структура отдельных особей и их групп. Однако выводы о консолиди-рованности и дифференциации отдельных внутрипопуля-ционных групп животных кардинально не менялись.
Кластерный анализ массива, генотипированных по маркерам, животных без учета их породы и особенностей генеалогии, позволил выявить и детализировать генетическую оригинальность отдельных особей и/или их групп, структуру и степень генетического различия между ними и, таким образом, получить недостающую информацию для планирования и реализации генофондосохранных мероприятий. В частности, по сравнению с классической холмогорской породой была показана более высокая дифференциация печорского типа с голштинской породой. Визуализация соотношения у отдельных особей вероятности их членства в отдельных кластерах дает возможность без опоры на генеалогию планировать предварительную систему закрытого воспроизводства генофондного поголовья при условии одновременной аттестации самцов и самок по микросателлитам.
