Внутрипопуляционная структура Helicopsis striata (Gastropoda, Pulmonata, Hygromiidae) в условиях лесостепного ландшафта юга Среднерусской возвышенности
 в условиях лесостепного ландшафта юга Среднерусской возвышенности")
Автор: Сычев А.А., Снегин Э.А.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Экология
Статья в выпуске: 9, 2013 года.
Бесплатный доступ
В статье описываются особенности внутрипопуляционной структуры наземного моллюска Helicopsis striata на примере модельной изолированной популяции, обитающей на юге Среднерусской возвышенности. Выявлена морфологическая дифференциация и определены демографические характеристики субпопуляций. На основе анализа изоферментов изучена структура генофондов исследуемых групп. Вычислена эффективная численность и дается прогноз времени дальнейшего существования популяции.
Особо охраняемый вид, наземный моллюск, лесостепь, внутрипопуляционная структура
Короткий адрес: https://sciup.org/14083233
IDR: 14083233 | УДК: 575.22;
Intra-population structure of Helicopsis striata (Gastropoda, Pulmonata, Hygromiidae) in the forest-steppe landscape conditions of the Mid-Russia upland south
The intra-population structure peculiarities of terrestrial mollusk Helicopsis striata on the example of the model isolated population located in the south of Mid-Russian Upland are described in the article. The morphological differentiation is revealed and the demographic characteristics of subpopulations are defined. The gene pool structure of the researched groups is studied on the basis of the isoenzyme analysis. The effective number is calculated and the forecast of the population further existence term is given.
Текст научной статьи Внутрипопуляционная структура Helicopsis striata (Gastropoda, Pulmonata, Hygromiidae) в условиях лесостепного ландшафта юга Среднерусской возвышенности
В этой связи весьма актуальным является анализ внутрипопуляционной структуры особо охраняемых видов с целью охраны и восстановления их популяций в естественных биотопах.
Цель исследований . Определить степень пространственной структурированности и ее значение в обеспечении жизнеспособности модельной популяции наземного моллюска Helicopsis striata (Müller, 1774) в условиях лесостепи Среднерусской возвышенности.
Данный вид является индикатором реликтовых лугостепных экосистем, называемых «сниженные Альпы», занесен в охранный список МСОП [3] и Красную книгу Белгородской области [4].
Стоит также отметить, что южная лесостепь Среднерусской возвышенности является восточной границей видового ареала H. striata , где он часто формирует сильно изолированные локальные популяции. Особенности адаптаций таких групп в условиях малой подвижности улиток, безусловно, заслуживают внимание.
Материалы и методы исследований. В качестве модели нами выбрана изолированная популяция H. striata , обитающая в окрестностях г. Белгорода (урочище «Белая гора»). Группа расположена на правом крутом берегу реки Северский Донец и представлена цепью из пяти субпопуляций или демов (рис. 1; координаты 50º37’28,66’’ с.ш., 36º37’15,97’’в.д.). Общая длина занимаемого биотопа составляет около 700 м. Субпопуляции в различной степени изолированы друг от друга оврагами, пешеходными дорожками, дачными застройками, меловыми обнажениями.
Материал собирался в 2010–2011 гг. вручную. В каждой субпопуляции на участке площадью 25 м2 проводилось описание биотопа, определялась плотность населения живых особей H. striata. На собранных особях подсчитывалось количество оборотов с округлением возраста до четверти. Для упрощения анализа собранных особей разбивали на четыре условные возрастные группы: I – 2,25–3,0 оборота раковины; II – 3,25–4,0; III – 4,25–5,0; IV – 5,25–6,0 оборотов. После определения возраста моллюсков возвращали обратно в биотоп. На каждом участке случайным образом отбирались живые особи с пятью оборотами на раковине для промера основных морфометрических параметров и расчета стандартных морфометрических индексов [5, 6]. Промеры проводились на бинокулярном микроскопе МБС-10 с окулярмикрометром. Промерено было 223 раковины.

Рис. 1. Схема расположения субпопуляций H. striata в биотопе «Белая гора»
Также в пределах обозначенных участков проводились выборки живых особей H. striata для анализа генетической структуры. В качестве генетических маркеров были использованы двухаллельные мономеры неспецифических эстераз ( EST7 и EST8 ) и двухаллельный димер супероксиддисутазы ( SOD2 ), полученные с помощью диск-электрофореза в вертикальных пластинах 10 % полиакриламидного геля (камера VE-3, «Heli-con»). Генотипы определены у 198 особей. Кроме этого, в субпопуляции №2 для определения особенностей распределения особей внутри группы было сформировано 28 пробных площадок размером 1 м2, где также определялась плотность поселения и проводился отбор улиток для морфометрического анализа. Данные обрабатывались в пакетах статистических программ Excel, Statistica 6, GenAlEx и TFPGA.
Результаты исследований и их обсуждение. Согласно полученным данным, демографическая структура изучаемой популяции выглядит неоднородной (табл. 1). При этом наблюдается прямая корреляция между увеличением плотности особей в субпопуляциях и повышением долей неполовозрелых особей I и II возрастов (R=0,99±0,05; P=0,001). Наибольшие размеры особей, а соответственно наименьшая плотность, отмечены в группе №1 (табл. 2). Аналогичную картину мы наблюдали при распределении особей внутри субпопуляций №2, где размеры особей также во многом определяются плотностью населения (рис. 2). Однофакторный дисперсионный анализ морфометрических признаков подтвердил наш вывод о неоднородно- сти внутрипопуляционной структуры – по всем конхиологическим признакам отмечена достоверная дифференциация (табл. 3).
Таблица 1
Плотность населения особей и демографическая структура в субпопуляциях
|
Cубпопуляция |
Число особей на 1 м2 |
Доли особей различных возрастных классов |
|||
|
I |
II |
III |
IV |
||
|
1 |
20 |
0,000 |
0,067 |
0,600 |
0,333 |
|
2 |
352 |
0,057 |
0,239 |
0,701 |
0,004 |
|
3 |
131 |
0,041 |
0,122 |
0,786 |
0,051 |
|
4 |
483 |
0,116 |
0,312 |
0,566 |
0,006 |
|
5 |
195 |
0,110 |
0,116 |
0,753 |
0,021 |
Таблица 2
Средние значения морфометрических признаков раковины (мм) в субпопуляциях H. striata (M±Δ, P =0,95)
|
N |
БД |
ВР |
ВЗ |
ШУ |
ВУ |
V р (мм3) |
Sу (мм2) |
|
36 |
11,8±0,2 |
8,2±0,2 |
3,1±0,2 |
5,7±0,1 |
5,5±0,1 |
572,1±32,1 |
24,9±1,1 |
|
48 |
9,7±0,2 |
7,0±0,1 |
2,9±0,1 |
4,6±0,1 |
4,5±0,1 |
338,7±18,8 |
16,3±0,8 |
|
42 |
10,1±0,2 |
7,2±0,1 |
3,0±0,1 |
4,8±0,1 |
4,7±0,1 |
368,6±17,3 |
17,8±0,7 |
|
52 |
8,9±0,2 |
6,6±0,1 |
3,0±0,1 |
4,2±0,1 |
4,0±0,1 |
263,9±13,2 |
13,3±0,5 |
|
45 |
10,5±0,2 |
7,1±0,1 |
2,8±0,1 |
5,0±0,1 |
4,8±0,1 |
397,1±17,7 |
18,8±0,7 |
Примечание: N – объем выборки, %; БД – большой диаметр раковины; ВР – высота раковины; ВЗ – высота завитка; ШУ – ширина устья; ВУ – высота устья; V р – объем раковины; Sу – площадь устья.
Стоит отметить, что аналогичную картину мы наблюдали при сопоставлении конхиологических признаков между популяциями, обитающими на юге Среднерусской возвышенности [6], то есть межпопуляционные различия по размерным характеристикам вполне сопоставимы с внутрипопуляционной дифференциацией. Такая картина, вероятно, обусловлена микроклиматическими особенностями биотопов, формирующих конституцию животных в ходе онтогенеза, а также намечающейся генетической дивергенцией между субпопуляциями.
Таблица 3
Результаты однофакторного дисперсионного анализа (ANOVA) морфометрических признаков в субпопуляциях
|
Показатель |
Источник вариации |
SS |
df |
MS |
F |
Fst |
|
БД |
Между группами |
192,8 |
4 |
48,202 |
128,7 |
2,4 |
|
Внутри групп |
81,7 |
218 |
0,375 |
|||
|
ВР |
Между группами |
51,8 |
4 |
12,962 |
62,8 |
|
|
Внутри групп |
45,0 |
218 |
0,206 |
|||
|
ВЗ |
Между группами |
1,5 |
4 |
0,376 |
2,7 |
|
|
Внутри групп |
30,4 |
218 |
0,139 |
|||
|
ШУ |
Между группами |
50,8 |
4 |
12,708 |
109,5 |
|
|
Внутри групп |
25,3 |
218 |
0,116 |
|||
|
ВУ |
Между группами |
51,3 |
4 |
12,820 |
122,4 |
|
|
Внутри групп |
22,8 |
218 |
0,105 |
|||
|
V |
Между группами |
2127102,5 |
4 |
531775,630 |
120,5 |
|
|
Внутри групп |
962094,9 |
218 |
4413,279 |
|||
|
S |
Между группами |
3004,1 |
4 |
751,023 |
121,1 |
|
|
Внутри групп |
1352,0 |
218 |
6,202 |
Б
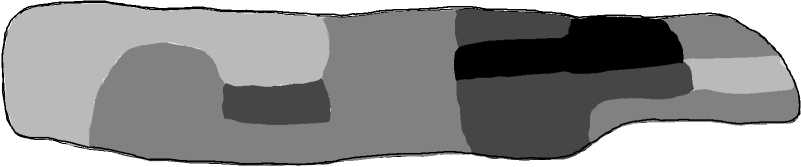
Низкая плотность (100 особей на 1 м2)
Средняя плотность (100250 особей на 1 м2)
Высокая плотность (250-400 особей на 1 м2) Очень высокая плотность (>400 особей на 1 м2)
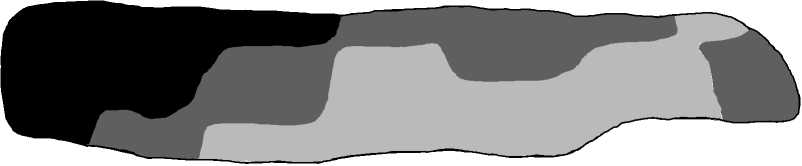
Мелкие особи
(300 мм3)
Особи средних размеров (300-400 мм3)
Крупные особи (>400 мм3)
Рис. 2. Пространственная дифференциация субпопуляции № 2 по плотности населения особей (А) и объему раковины (V р ) (Б)
Частоты аллелей и показатели генетической изменчивости приведены в табл. 4. Согласно полученным данным, отмечается варьирование показателей генетической изменчивости между субпопуляциями как по отдельным локусам изоферментов, так и по усредненным значениям. Самой мономорфной оказалась субпопуляция №5, где отмечена полная гомозиготность по аллелю EST7-1 . Тем не менее уровень генетической дифференциации между субпопуляциями, оцениваемый по коэффициенту инбридинга F st =0,090 (табл. 5), демонстрирует незначительную разобщенность исследуемых демов, особенно при сопоставлении с показателем генетической дифференциации популяций H. striata , полученным нами ранее для всего юга Среднерусской возвышенности ( F st =0,356) [6]. Таким образом, генетическая структура, оцениваемая с помощью полиморфных изоферментных локусов, больше соответствует пространственному расположению субпопуляций, чем морфологические параметры раковины, подверженные модификационной изменчивости. Этот тезис подтверждается построенной дендрограммой генетических расстояний между субпопуляциями по Нею [7] (рис. 3), которая указывает на прямую зависимость генетического родства изучаемых демов от степени их пространственной удаленности, что соответствует модели изоляции расстоянием.
Таблица 4
Частоты аллелей и показатели генетической изменчивости в субпопуляциях H. striata
|
Субпопуляция |
Локус |
Аллель |
A |
Ae |
I |
Ho |
He |
F |
|
|
1 |
2 |
||||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
|
1 ( N =40) |
EST7 |
0,675 |
0,325 |
2 |
1,782 |
0,631 |
0,550 |
0,439 |
-0,254 |
|
EST8 |
0,45 |
0,55 |
2 |
1,980 |
0,688 |
0,400 |
0,495 |
0,192 |
|
|
SOD2 |
0,925 |
0,075 |
2 |
1,161 |
0,266 |
0,150 |
0,139 |
-0,081 |
|
|
Среднее |
2 |
1,641 |
0,528 |
0,367 |
0,358 |
-0,048 |
|||
|
2 ( N =40) |
EST7 |
0,8 |
0,2 |
2 |
1,471 |
0,500 |
0,300 |
0,320 |
0,062 |
|
EST8 |
0,538 |
0,463 |
2 |
1,989 |
0,69 |
0,625 |
0,497 |
-0,257 |
|
|
SOD 2 |
0,788 |
0,213 |
2 |
1,503 |
0,517 |
0,375 |
0,335 |
-0,120 |
|
|
Среднее |
2 |
1,654 |
0,569 |
0,433 |
0,384 |
-0,105 |
|||
Окончание табл. 4
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
|
3 ( N =38) |
EST7 |
0,908 |
0,092 |
2 |
1,201 |
0,307 |
0,184 |
0,167 |
-0,101 |
|
EST8 |
0,289 |
0,711 |
2 |
1,699 |
0,602 |
0,579 |
0,411 |
-0,407 |
|
|
SOD2 |
0,579 |
0,421 |
2 |
1,951 |
0,681 |
0,526 |
0,488 |
-0,080 |
|
|
Среднее |
2 |
1,617 |
0,53 |
0,430 |
0,355 |
-0,196 |
|||
|
4 ( N =40) |
EST7 |
0,8 |
0,2 |
2 |
1,471 |
0,500 |
0,350 |
0,320 |
-0,094 |
|
EST8 |
0,212 |
0,788 |
2 |
1,503 |
0,517 |
0,325 |
0,335 |
0,029 |
|
|
SOD2 |
0,788 |
0,213 |
2 |
1,503 |
0,517 |
0,375 |
0,335 |
-0,120 |
|
|
Среднее |
2 |
1,492 |
0,512 |
0,350 |
0,33 |
-0,062 |
|||
|
5 ( N =40) |
EST7 |
1 |
0 |
1 |
1,0 |
0,000 |
0,000 |
0,000 |
- |
|
EST8 |
0,563 |
0,437 |
2 |
1,969 |
0,685 |
0,525 |
0,492 |
-0,067 |
|
|
SOD2 |
0,563 |
0,438 |
2 |
1,969 |
0,685 |
0,425 |
0,492 |
0,137 |
|
|
Среднее |
1,7 |
1,646 |
0,457 |
0,317 |
0,328 |
0,035 |
|||
|
Общее среднее |
1,9 |
1,61 |
0,519 |
0,379 |
0,351 |
-0,083 |
|||
Примечание: N – число особей в выборке; A – наблюдаемое число аллелей; Ae – эффективное число аллелей; I – индекс Шеннона; Ho – средняя наблюдаемая гетерозиготность; He – средняя ожидаемая гетерозиготность; F – коэффициент инбридинга.
В заключение нами был проведен расчет эффективной численности как отдельных демов, так и популяции в целом, с целью оценки потенциального времени их дальнейшего существования. Срок жизни определялся по формулам t=1,5 Ne , где t – количество поколений [8]; Ne=(4N-2)/(V+2 ), где N – численность половозрелых особей в популяции; V – дисперсия индивидуальной плодовитости [9].
Таблица 5
Коэффициенты инбридинга на разных уровнях иерархической структуры популяции H. striata по результатам изоферментного анализа
|
Локус |
Популяция |
Субпопуляция №2 |
||||
|
F is |
F it |
F st |
F is |
F it |
F st |
|
|
EST7 |
0,11 |
0,012 |
0,089 |
-0,07 |
-0,047 |
0,022 |
|
EST8 |
0,1 |
0,014 |
0,078 |
0,115 |
0,124 |
0,009 |
|
SOD2 |
0,04 |
0,064 |
0,097 |
-0,024 |
-0,015 |
0,009 |
|
Среднее |
0,08 |
0,013 |
0,090 |
0,007 |
0,021 |
0,013 |
Примечание: F it – коэффициент инбридинга особи относительно популяции; F is – коэффициент инбридинга особи относительно субпопуляции; F st – коэффициент инбридинга субпопуляции относительно популяции .
1 .075 .05 .025 0,000
------------- 1
----------- 4
---------------------------------- 5
Рис. 3. Дендрограмма генетических расстояний субпопуляций (UPGMA)
Для определения дисперсии индивидуальной плодовитости было изучено 83 кладки H. striata . Кладки яиц были получены в период массового спаривания H. striata по методике, описанной нами ранее [6, 10]. Согласно данным, представленным в табл. 6, размер кладки прямо пропорционален размеру животных ( R =0,97±0,08; P =0,001). Тем не менее дисперсия количества отложенных яиц уменьшается параллельно с уменьшением размеров половозрелых особей и увеличения их численности. А это в свою очередь значительно повышает срок дальнейшего существования популяции. Самой уязвимой группой, по нашим данным, является субпопуляция №1, где в условиях малой плотности обитают наиболее крупные улитки, обладающие наибольшими размерами кладок и наибольшим значением дисперсии индивидуальной плодовитости. Предполагаемый срок жизни этой субпопуляции оценивается 160 годами.
При этом стоит отметить, что усредненные показатели потенциальной продолжительности жизни всей популяции имеют более низкую оценку (84088 лет), чем некоторых субпопуляций. Самым жизнеспособным демом, по нашим данным, является группа №2, предполагаемый срок жизни которой равен 421131 год.
Данные по кладкам яиц, эффективная численность субпопуляций
H. striata и прогноз времени их дальнейшего существования
Таблица 6
|
Cубпопуляция |
Число кладок |
Среднее количество яиц в кладке |
U |
N |
V |
Ne |
Ne/N (%) |
Т |
|
1 |
10 |
31,6±11,6 |
19 |
4675 |
348,9 |
53 |
1,1 |
160 |
|
2 |
16 |
12,9±1,4 |
248 |
372000 |
8,6 |
140377 |
37,7 |
421131 |
|
3 |
19 |
12,4±3,1 |
109 |
874400 |
47,0 |
71380 |
8,2 |
214139 |
|
4 |
14 |
9,9±1,2 |
276 |
110400 |
4,9 |
64000 |
58,0 |
191999 |
|
5 |
24 |
19±2,8 |
151 |
150700 |
47,9 |
12080 |
8,0 |
36240 |
|
Популяция |
83 |
14,2±1,5 |
136 |
1512175 |
107,9 |
28029 |
1,8 |
84088 |
Примечание: U – плотность половозрелых особей (м2); N – общая предполагаемая численность половозрелых особой; V – дисперсия индивидуальной плодовитости; Ne – эффективная численность; Т – предполагаемое время существования группы в годах, которое рассчитывалось исходя из того, что срок жизни одного поколения H. striata составляет два года.
Заключение. Таким образом, на основании полученных данных о связи плотности населения особей их средних размеров и плодовитости можно сделать вывод о действии внутрипопуляционных механизмов регуляции функционального состояния популяции в колеблющихся условиях среды. При этом средние значения демографических и морфофизиологических параметров в большей степени определяются экологическими факторами. В связи с этим пространственная дифференциация изолированной популяции H. striata происходит в основном по количественным признакам раковины, плотности населения особей в биотопе и их плодовитости. При этом по частотам аллелей генетических маркеров такой степени подразделенности нами не выявлено.
Стоит отметить также, что морфофизиологическое своеобразие локальных групп на фоне общего генофонда значительно повышает уровень жизнеспособности популяции в целом за счет увеличения приспособленности наиболее «успешных» субпопуляций. .
