Внутрипопуляционная вариабельность в потомствах F3 и F4 томата, полученных от рекомбинантов F2, устойчивых к высокой температуре по признакам мужского гаметофита

Автор: Маковей Миланья Дмитриевна
Журнал: Овощи России @vegetables
Рубрика: Селекция и семеноводство сельскохозяйственных растений
Статья в выпуске: 4 (48), 2019 года.
Бесплатный доступ
Актуальность Выявление устойчивых к температурным стрессам форм важно не только на этапе поиска, но и позднее в селекционном процессе. Для оценки устойчивости растений, наряду с действием естественных факторов, необходимо их моделировать в лабораторных условиях. Материал и методика Представлены результаты изучения изменчивости признаков мужского гаметофита томата (жизнеспособность пыльцы, длина пыльцевых трубок, устойчивость пыльцы и устойчивость по длине пыльцевых трубок) в гибридных потомствах F2, F3, F4, полученных от разных комбинаций скрещивания с высокими показателями устойчивости к высокотемпературному стрессу (8 гибридных комбинаций). Результаты В расщепляющихся популяциях F2 по комплексу признаков пыльцы установлены рекомбинационные различия, выраженные в показателе средней дисперсии по семье (S2), что позволило выделить трансгрессивные рекомбинанты, получить семена для проведения следующих циклов отбора в F3 и далее. Выявлено, что в потомствах F3 и F4 сохраняется высокая внутрипопуляционная вариабельность признаков мужского гаметофита, что отражает реакцию их генотипов на высокую температуру...
Томат, гибридные популяции, признаки, изменчивость, рекомбинанты, потомства, устойчивость пыльцы, устойчивость по длине пыльцевых трубок
Короткий адрес: https://sciup.org/140240734
IDR: 140240734 | УДК: 635.64:631.526.325:631.524.85 | DOI: 10.18619/2072-9146-2019-4-37-43
Intrapopulation variability for male gametophyte traits in the tomato F3 and F4 offspring derived from high temperature resistant F2 recombinants
Relevance Identification of forms resistant to thermal stress is important not only at the stage of search, but also later in the selection process. Methods The paper presents the results obtained during the studies on the variability of tomato male gametophyte traits (pollen viability, pollen tube length, pollen resistance and resistance along pollen tubes) in the hybrid F2, F3 and F4 offspring derived from different crossing combinations strongly resistant to high temperature stress (8 hybrid combinations). Results Recombination differences expressed in the values of average variation per family (S2) have been found in the F2 populations segregated for a complex of pollen traits, which made it possible to isolate transgressive recombinants from different hybrid combinations, to produce seeds from them and to further perform selection cycles in F3. It has been found that high intrapopulation variability of male gametophyte traits is preserved in F3 and F4 offspring, which reflects the response of their genotypes to high temperature...
Текст научной статьи Внутрипопуляционная вариабельность в потомствах F3 и F4 томата, полученных от рекомбинантов F2, устойчивых к высокой температуре по признакам мужского гаметофита
В основе любых селекционных программ лежит задача улучшения наследственных свойств объекта исследований. Селекционеру для её решения необходимо на всех этапах селекционного процесса знать изменчивость признаков, в особенности под влиянием каких бы то ни было факторов внешней среды. Это стало востребованным в последние годы, когда наблюдаются значительные катаклизмы на европейской части континента, что несомненно усиливает актуальность исследований, нацеленных на селекцию форм, способных в условиях, отличных от оптимальных, реализовать генетически заложенный потенциал продуктивности. Одной из важных и сложных задач в этом контексте является устойчивость к температурным стрессам. Выявление устойчивых форм важно не только на этапе поиска, но и позднее в селекционном процессе. Как правило, чаще всего они выделяются в F 2 и в популяциях гибридов F 3 . Однако оценка уровня и выделение устойчивых форм в естественных условиях усложняется совокупностью действия комплекса стрессоров [1], что иногда делает её невыполнимой в полевых условиях. Поэтому для оценки устойчивости растений, наряду с действием естественных факторов, необходимо их моделировать в лабораторных условиях. Преимущество их состоит в возможности регулирования уровня жесткости действия неблагоприятного фактора, на который ведется селекция (высокие, низкие температуры, засуха и др.) и, таким образом интенсифицировать оценку исходного материала и ускорить процесс селекции.
В последние десятилетия интенсивно развиваются исследования реакции пыльцы на действие различных абиотических и биотических факторов среды [2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13 и др.]. Пионерами работ по искусственному отбору по пыльце были А.В Алпатьев [14], Д.В. ТерАванесян [15]. Отбор на уровне мужского гаметофита впервые был теоретически обоснован только в 1971 году [16]. В основе этого метода селекции лежит предложение, что отбор на устойчивость к конкретному экстремальному фактору на гаплоидной фазе (популяции гамет) может обеспечить появление устойчивого споро-фитного поколения. Особое значение придается именно возможностям мужского гаметофита растений, потому что мужские гаметы эволюционно менее защищены, чем женские, закрытые от непосредственных воздействий толстыми слоями соматических тканей. Кроме того, растения продуцируют значительно большее количество мужских гамет, но в формировании плодов участвуют только десятки, сотни пыльцевых зерен, преодолевших непосредственное влияние всевозможных стрессовых факторов. Значит, мужские гаметы вовлечены в своеобразные природные селективные механизмы. Способность пыльцы прорастать и расти на искусственной питательной среде [17] дает возможность быстро и эффективно оценить, как жизнеспособность пыль- цы, так и реакцию на действие конкретного стрессового фактора. Мужской гаметофит, имея два преимущества перед спорофитным поколением – микроскопические размеры и гаплоидный генотип, позволяет проводить широкие исследования, в том числе, в лабораторных условиях, где анализу может быть подвергнуто большое количество генотипов. Эти характеристики пыльцы можно использовать с целью создания генофондов, обладающих высоким потенциалом устойчивости, или непосредственно в селекционном процессе при отборе устойчивых форм томата из гибридных популяций. С использованием гаметофитных отборов появляется возможность значительно ускорить селекционный процесс за счет неограниченного увеличения объема выборок и провести оценку и отборы на фонах любой степени жесткости. Такие отборы недоступны на других фазах жизненного цикла взрослых растений при выращивании их на больших площадях в связи с отсутствием возможности моделирования стрессовых фонов.
Цель исследований – оценить перспективность гибридных популяций томата в потомствах F 3 и F 4 , полученных из устойчивых к высокой температуре рекомбинантов по признакам мужского гаметофита (жизнеспособность пыльцы, длина пыльцевых трубок, устойчивость пыльцы и устойчивость по длине пыльцевых трубок), во взаимосвязи с ответной реакцией вегетирующего растения на отбор по ряду количественных признаков (количество соцветий на растении, число цветков на соцветии, число завязавшихся плодов, масса плода, количество семян в плодах).
Материал и методы
Экспериментальным материалом служили девять гибридных комбинаций томата (Л5 x Церос, Л5 x Лидер, Л5 x Л324, Л7 x Лидер, Л7 x Л126, Л7 x Церос, Л7 x Л305, Л7 x Л327, Л7 x Л324). Они получены от скрещивания форм с высокой устойчивостью пыльцы и пыльцевых трубок к высокой температуре. Для установления температурного режима и временной экспозиции воздействия на пыльцу томата, которая бы позволила идентифицировать и классифицировать изучаемые исходные формы по характеру и степени проявления их устойчивости к высокотемпературному фактору, использовали пять режимов: 35 ° С; 40 ° С; 45 ° С; 50 ° С с разной временной экспозицией воздействия на пыльцу – 3, 6 и 8 час.
Полученные результаты показали, что температурный режим 45°С c временной экспозицией воздействия на пыльцу в течение 8 часов позволяет наиболее четко дифференцировать образцы как по прорастанию пыльцевых зерен, так и способности их формировать пыльцевые трубки достаточной для оплодотворения длины (три диаметра пыльцевого зерна). Следовательно, для оценки и анализа гибридов F1 и определению доли влияния родительских компонентов скрещивания на характер проявления в гибридном потомстве устойчивости к повышенной температуре использо- вали именно этот режим (45°С/8 часов) [18]. Далее изучали потомства F2, F3 и F4 (отсчет поколений ведется по растению, выделенному в F1).
Гибриды выращивали на опытном поле ИГФиЗР на двухрядковых делянках в трех повторностях по единой схеме в разные годы. В качестве стандарта служили родительские формы. Оценку расщепляющихся популяций F 2 от каждой комбинации проводили индивидуально на 15-20 растениях. Каждое растение изучали по комплексу признаков пыльцы: жизнеспособность свежесобранной пыльцы и длина пыльцевых трубок (контроль); способность пыльцевых зерен прорастать, а проросших формировать длинные пыльцевые трубки после высокотемпературной обработки (45 ° С) в течение 8 часов (опыт). Длину пыльцевых трубок измеряли в делениях окуляр-микрометра.
Устойчивость признаков определяли по степени уменьшения показателей в опытном варианте по отношению к контролю [4].
Полученные результаты обрабатывали в двухфакторном дисперсионном комплексе с тремя повторностями и четырьмя градациями второго фактора. Мерой изменчивости изучаемых признаков в F 2 служит показатель средней дисперсия (S2) по семье.
По аналогичной схеме обрабатывали результаты, полученные в F 3 с определением доли влияния генотипа на характер проявления 2-х разных признаков – устойчивость пыльцы и устойчивость по длине пыльцевых трубок. Реакцию гибридных популяций на отбор (R) определяли по разнице между средним значением признаков всей популяции (Хп) и средним его значением у генотипов, выделенных в предыдущем поколении, а также с показателем лучших рекомбинантов в F 2 к F 3 и F 4 .
Потомства F 4 изучали и по ряду количественных признаков: высота закладки первого соцветия, число соцветий на растении, число цветков на соцветии, количество завязавшихся плодов на соцветии и число семян в плодах. Степень внутрипопуляцион-ной вариабельности их отражает коэффициент вариации (cv, %) [19]. Таким образом, определяли связь между уровнем устойчивости рекомбинантных форм, выделенных в F 2 по комплексу признаков пыльцы, и формированием из них устойчивых потомств в F 3 и F 4 с более высокими показателями.
Результаты и их обсуждение
Анализ расщепляющихся популяций F2, от каждой гибридной комбинации по комплексу признаков пыльцы (жизнеспособность свежесобранной пыльцы и длина пыльцевых трубок, устойчивость пыльцы и устойчивость по длине пыльцевых трубок) выявил высокую гетерогенность между растениями в популяциях по изученным признакам: уступающие обоим родительским формам; имеющие средние значения между двумя родительскими формами; превышающие лучшего родителя или на его уровне. Это позволило выделить лучшие растения (трансгрессивные рекомбинанты), получить с них семена для следующего цикла отборов в F3, и далее.
Например, в популяции гибрида F 2 (Л7 x Лидер) практически все генотипы имели значения признаков ниже худшего и среднего двух родителей. Высокая дисперсия по семье (S2 – 4620) между средними одного признака и по всем изученным позволила выделить только 4 генотипа, у которых значения признаков были на уровне родителя с меньшей устойчивостью. Практически такой же уровень изменчивости (S2 = 4502) выявлен и в популяции от комбинации Л5 x Л324, но здесь выделяются три генотипа, у которых значения по комплексу признаков выше, чем у лучшей родительской формы (табл. 1).
Из комбинаций гибридов F 2 – Л7 x Церос и Л7 x Л327 на фоне высокой внутрипопуляционной гетерогенности выделились два и один генотип, соответственно, со значением признаков превышающих среднюю между их родительскими формами. Из других двух популяций – Л7 x Л324 и Л5 x Церос выделено только по 2 растения с показателями ниже или на уровне среднего их родителей. При анализе средних дисперсий по семьям видно, что изменчивость в популяциях по анализируемым признакам на фоне высокой температуры достаточно высокая (S2 = 5764; S2 = 5035; S2 = 5848 и S2 = 4645). Они отличаются наличием большого числа генотипов с крайне низкими и средними значениями признаков и лишь незначительным количеством с высокими показателями по отдельным признакам.
Низкий спектр изменчивости (S2 = 2647), относительно других выделяет комбинацию, скрещивания Л7 x Л126. В популяции частота встречаемости генотипов, превышающих среднее значении 2-х родительских форм по изученным признакам пыльцы высокая. Выделено 4 растения с показателем признаков выше, чем у лучшего родителя (Л126).
Очень широкий спектр изменчивости (S2 = 8015) отмечен у гибрида F 2 от комбинации (Л7 x Л305). Популяцию отличает наличие большого количества генотипов с низкими и средними значениями исследованных признаков, но одновременно отмечается присутствие и с достаточно высокими показателями, что позволило выделить три рекомбинанта (табл.1).
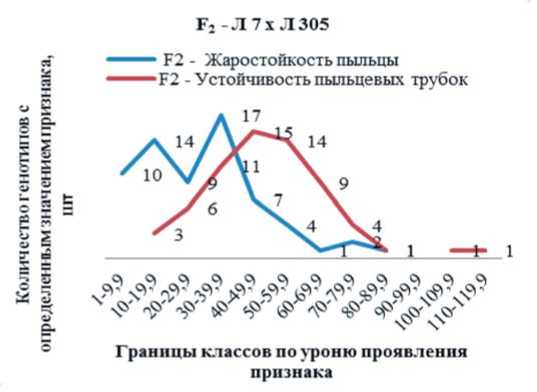
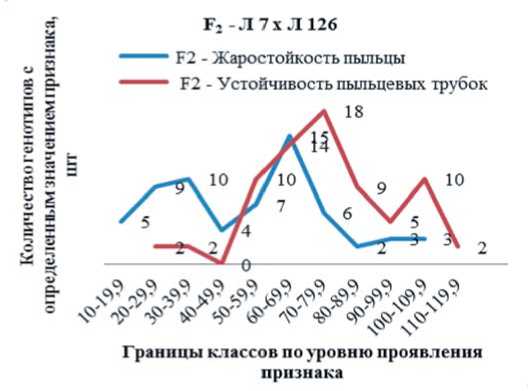
Исходя из характера внутрипопуля-ционной вариабельности, было определено число классов и выявлены их границы по степени выраженности признаков «жаростойкость пыльцы и устойчивость по длине пыльцевых трубок», что демонстрируется на диаграмме вариационного распределения для 2-х гибридных комбинаций: Л7 x Л126 и Л7 x Л305, которые в 2-х факторном комплексе дисперсионного анализа демонстрируют контрастный размах изменчивости (рис. 1 а, б). В комбинации Л7 x Л305, средняя дисперсия по семье (S2 = 8015) в 3 раза выше, чем в Л7 x Л126 (S2 = 2647) (табл. 1).
В популяции гибрида Л5 x Л305 выявлены генотипы с крайне низкими и крайне высокими значениями по признаку «жаростойкость пыльцы», которые варьируют от 1,0-9,9% до 80,0-89,9%. Большая часть их (50,8%) характеризуются низкими (1,0-29,9%) показателями. Средние значения (30,0-49,9%) имели 32,3% генотипов, и лишь 16,9% проявляют высокую жаростойкость пыльцы (50,0-89,9%). Генотипов, относящихся к классу с очень высокими значениями (90,0-
109,9%), не выявлено (рис. 1а).
Аналогичные результаты получены по признаку «устойчивость по длине пыльцевых трубок». У 53,8% генотипов показатель этого признака падает на классы с низким (20-29,9%) и средним (30,0-49,9%) значением. Выделяются два генотипа, которые после термообработки пыльцы формируют очень длинные трубки, устойчивость которых составляет 100119,9%, что выше, чем в контроле. Остальные 43,1% относятся к классам с высоким показателем этого признака (50,0-89,9%) (рис. 1а).
Несколько иной характер внутрипо-пуляционной изменчивости наблюдается у гибрида F 2 от комбинации Л7 x Л126 по обоим признакам. Из диаграммы 1 б видно, что кривая распределения генотипов внутри популяции смещена в сторону классов с высокой жаростойкостью пыльцы (50,089,9%), что составляет 47,7%. Высокоустойчивую пыльцу (90,0109,9%) имели 9,2% генотипов. Остальные (23,0%) относятся к классам со средним (30,0-49,9%) и (20,1%) низкими (10,0-29,9%) значениями признаков.
Здесь важно отметить, что генотипов с крайне низкими значениями (1,0-9,9%) не выявлено.
Анализ характера распределения генотипов внутри популяции по признаку «устойчивость по длине пыльцевых трубок» показывает, что 61,2% относятся к классам с высокими значениями (50,0-89,9%). К классам с крайне высокими показателями признака (90,0-119,9%) относятся 26,1% генотипов и только 12,7% характеризуются низким и средним уровнем проявления признака. Генотипов, относящихся к классам с крайне низкими (1-9,9%) и низкими (10-19,9%)
Таблица 1. Результаты дисперсионного анализа и характеристика расщепляющихся гибридных популяций F 2 по комплексу признаков пыльцы
Table 1. Results of analysis of variance and characterization of populations segregated hybrid F 2 by a complex of pollen traits
|
Номер гибридной семьи |
Гибридная комбинация |
Изучено растений |
№ выделившегося растения |
Средний показатель по всем признакам*, % |
Средняя дисперсия по семье (S2) |
|
29 |
Л5 x Церос |
15 |
5 4 |
46,6 44,9 |
4645 |
|
31 |
Л5 x Л324 |
20 |
9 14 20 |
64,7 W 70,8 WV 66,2 W |
4502 |
|
33 |
Л7 x Церос |
15 |
7 9 |
58,6 V 51,4 V |
5764 |
|
34 |
Л7 x Лидер |
20 |
14 16 18 19 |
44,6 41,8 44,2 41,3 |
4620 |
|
35 |
Л7 x Л126 |
20 |
4 8 9 12 |
61,5 W 60,7 W 69,0 W 64,5 W |
2647 |
|
37 |
Л7 x Л305 |
20 |
8 12 15 |
55,5 V 61,9 W 53,1 V |
8015 |
|
38 |
Л7 x Л324 |
15 |
6 2 |
58,3 V 42,4 |
5848 |
|
39 |
Л7 x Л327 |
20 |
5 |
52,0 V |
5035 |
Примечание: 1.* – жизнеспособность, жаростойкость пыльцы, длина пыльцевых трубок устойчивость по длине пыльцевых трубок к высокотемпературному фактору стресса.
-
2. V – устойчивый генотип, W – с высокой устойчивостью и WV – сверхустойчивые генотипы
а
Рис. 1 (а, б). Генотипическая структура расщепляющихся популяций F 2 от разных гибридных комбинаций по частоте встречаемости генотипов с определенным уровнем значения признаков: жаростойкость пыльцы и устойчивость пыльцевых трубок
б
Fig. 1 (a, b). The genotypic structure of the segregated F2 populations from different hybrid combinations on frequency of occurrence of genotypes with the certain level of value of traits: heat-tolerance of pollen and resistance along pollen tubes значениями этого признака, не выделено (рис. 1б). По способности проросшей пыльцы формировать длинные пыльцевые трубки после её обработки высокой температурой (45°С/8 час), показатели выше в популяциях обоих гибридных комбинаций.
Как результат, из гибридных семей F 2 удалось отобрать 20 генотипов, в геноме которых сочетаются генетические факторы, обеспечивающие на фоне стрессовой температуры высокую устойчивость пыльцы, а также отвечающие за рост и формирование длинных трубок. Но только 14 выделенных из разных гибридных семей имели значения признаков, превышающие среднюю между родителями или лучшего из них. Они характеризуются как устойчивые и высокоустойчивые. Показатели на уровне среднего двух родителей имели 6 генотипов. Лучшие рекомбинанты выделены из комбинаций: Л5 x Л324, Л7 x Л126, Л7 x Л305, Л7 x Л324 и Л7 x Церос. Из представленных данных видно, что частота образования устойчивых по комплексу признаков пыльцы трансгрессивных форм со значительным превышением значений их исходных форм в популяциях F 2 не так высока, даже если их родительские формы характеризуются достаточно высокими показателями анализируемых признаков. И всё же, несмотря на это, разработанный метод с использованием комплекса признаков мужского гаметофита позволяет начать отбор в F 2 , что очень важно при селекции на устойчивость к стрессовым абиотическим факторам.
О селекционной ценности выще-пившихся трансгрессивных форм можно судить только по анализу характера проявления признаков пыльцы в более поздних поколениях [20]. Для этого популяции F3 формировали из семян лучших рекомбинантов, выделенных в F2. Оценка и анализ их потомств в следующей генерации F3, а затем и F4, позволит определить эффективность используемого метода отбора и его влияние на характер проявления не только признаков пыльцы, но и ряда других количественных признаков.
Дисперсионный анализ результатов оценки исследуемых признаков пыльцы по всем комбинациям в F 3 показывает, что действие изучаемых факторов и их взаимодействие значимо на самом высоком уровне (P<0,001). Анализ взаимодействия факторов «генотип x признак» и «генотип x температура» показывает, что у генотипов в популяциях F 3 показатели изученных признаков ниже, чем у растения, от которого получено это потомство (табл. 2).
Смещение показателей анализируемых признаков пыльцы в сторону их уменьшения в пределах семьи F3 имеет место внутри каждой из изученных комбинаций (табл. 2). Например, в популяции гибрида F3 (Л7 x Л305), полученного от растения №8 (F2), которое имело среднее значение (55,5%) по комплексу признаков, формируется наиболее устойчивое потомство. При анализе взаимодействия факторов (генотип x температура) выделяется 5 растений – № 3; 5; 8; 9; 11. Значения показателей признаков пыльцы у данных растений после воздействия на нее температурой 45°С в течение 8 часов достаточно высокие (табл. 2). Доля влияния генотипа на проявление признаков: устойчивость пыльцы и устойчивость трубок у вышеприведенных растений, как видно из таблицы 2, очень высокая. Причем доля влияния генотипа на признак «устойчивость пыльцевых трубок» значительно выше, чем на «жаростойкость пыльцы». Отличные от этих получены результаты в популяциях потомств F3 от лучших рекомбинантов №12 и №15, генотипы демонстрируют показатели значительно ниже, чем в F2 – 41,3% и 40,0% против 61,9% и 53,1% – по жаростойкости пыльцы и 35,1% и 43,4% против 72,3% и 66,7% – по устойчивости пыльцевых трубок, соответственно. Из этих семей не удалось получить растения, которые бы имели показа- тель по комплексу анализируемых признаков на уровне среднего их родителей или выше.
Несколько иные результаты получены по комбинации Л7 x Л126, где в F 2 частота встречаемости растений, относящихся к классам с более высокими показателями, была выше (рис. 1 б). Лучшие растения 4, 8, 9 и 12, выделенные в F 2 из этой комбинации, формируют в F 3 более однородные популяции, но при этом средние значения по комплексу признаков пыльцы смещены в сторону классов с низкими и средними показателями. Из потомств растений № 4, 9 и 12 наиболее высокими значениями в F 2 (№4 – 61,5%; №9 – 69,0%; №12 – 64,5%) в F 3 не удается выделить ни одного генотипа с равнозначным показателем или выше среднего по популяции. Потомство F 3 , полученное от растения №8 (F 2 – 60,7%), характеризуется наличием в популяции большего числа генотипов со средним и высокими значениями изученных признаков. Это дает возможность выделить пять растений (табл. 2). Лучшие рекомбинанты F 2 – №7 и №20, выделенные из комбинаций Л7 x Церос и Л5 x Л324, в F 3 дают потомства, устойчивость которых ниже, чем у генотипов, из которых они были получены. Дисперсионный анализ взаимодействия факторов «генотип x температура» и «генотип x признаки» показывает, что в популяциях F 3 не обнаруживается растений с более высокой выраженностью показателей анализируемых признаков, чем у растений F 2 (табл. 1 и 2).
Идентичные результаты получены в F 3 и по комбинациям Л5 x с. Церос, Л7 x с. Лидер.
В целом анализ потомств гибридов F 3 , полученных из лучших рекомбинантов, выделенных в F 2 , показывает, что индивидуальный отбор по комплексу признаков пыльцы приводит к изменению структуры гибридных популяций F 3 , которое выражается в уменьшении или полном отсутствии генотипов с крайними значениями
Таблица 2. Характеристика гибридных потомств F 3 , полученных от лучших рекомбинантов выделенных в F 2 по комплексу признаков пыльцы на высокотемпературном фоне
Table 2. Characterization of hybrid offspring F 3 obtained from the best recombinants isolated in
F 2 on the complex of pollen traits on a high-temperature background
Наиболее высокая степень совпадения выявлена у 5 растений от комбинации Л7 x Л305, хотя при сравнительном анализе мы всё же видим, что доля влияние генотипа на устойчивость трубок выше (табл. 2). У растений, выделенных из популяций F 3 других комбинаций, наблюдается как совпадения, так и значительные различия по доле влияния генотипа, как, казалось бы, на два взаимозависимых признака. Например, у растения №12 (Л7 x с. Церос), доля влияния генотипа на проявление признака «жаростойкость пыльцы» составляет 38,3%, тогда как «устойчивость пыльцевых трубок» определяется генотипом на 60,9%. Эти различия выше в 1,8 раза и у растения №13 (Л5 x Л324). Аналогичные результаты получены и по растению №2 от комбинации Л5 x с. Церос. Сильно выражены (2,2 раза) эти различия и у растения №3, полученного из комбинации Л7 x с. Лидер (табл. 2).
Выявленные особенности доли влияния генотипа на характер проявления признаков «устойчивости пыльцы и устойчивость по длине пыльцевых трубок» показывают, что в популяции одной конкретной комбинации появляются генотипы, у которых доля влияния на характер проявления двух признаков неоднозначна. Можно предположить, что на характер их проявления влияют наследственные факторы в сочетании с фактором высокотемпературного стресса, посредством изменения различных биохимических и физиологических процессов.
Оценка популяций поколения F 4 , полученных из лучших растений по характеру проявления устойчивости по обоим признакам пыльцы у всех гибридных комбинаций, не подтверждает предположение получения более устойчивых потомств. Анализ селекционной ценности их по реакции гибридных популяций на отбор (R) показал, что фактический сдвиг по жаростойкости пыльцы в F 3 относительно F 2 невелик (0,92), и ещё ниже в F 4 к F 2 – 0,65. Несколько выше сдвиг показателей в динамике поколений F 2 , F 3 и F 4 от разных гибридных комбинаций был по устойчивости пыльцевых трубок 0,99 (F 3 к F 2 ) и 1,15 (F 4 к F 2 ). Наиболее выраженный положительный сдвиг в сторону высоких значений по данному признаку в F 3 и F 4 относительно F 2 отмечен только у трех комбинаций – Л7 x Л305, Л7 x сорт Церос, Л7 x сорт Лидер (R – 1,91, 1,29 и 1,08 соответственно).
В целом, селекционная ценность потомств, полученных из трансгре-сивных рекомбинантов F2 по признакам «жаростойкость пыльцы и устой- чивость пыльцевых трубок», не столь выражена, как ожидалось (табл. 3). Различия, установленные при оценке расщепляющихся популяций F2 по уровню и спектру изменчивости признаков пыльцы, не подтвердились в F3 и F4. Можно предположить, что связь между конкурентоспособностью рекомбинантов и разнообразием их потомств носит модификационный характер.
Оценка и анализ характера проявления количественных признаков и уровня их изменчивости (выраженный коэффициентом вариации) в потом-ствах F 4 , полученных от генотипов с разным уровнем устойчивости по признакам пыльцы 64,2% (Л7 х Л 305), 42,2% (Л7 х 126) и 34,7 (Л7 х Лидер), также выявляет высокую вариабельность. Установлена тенденция в направлении большего выхода растений с равнозначным значением изученных признаков в потомствах F 4 , полученных от растений F 3 со средним показателем устойчивости по признакам пыльцы. В то время как потомство, полученное из растений с более высоким показателем по комплексу признаков пыльцы, демонстрирует высокую вариабельность с наличием генотипов с меньшим значением количественных признаков в пределах популяции. На растениях завязалось меньше плодов, существенные различия имели место и по массе плода, а также их осемененности. В потомстве F 4 , полученном от растения с низким показателем, также наблюдается высокая изменчивость (cv, %) (табл. 4). Здесь следует отметить, что
Таблица 3. Реакция гибридных популяций F 3 , F 4 на отбор в F 2 по признакам пыльцы Table 3. The reaction of hybrid populations F 3 , F 4 on selection in F 2 on the traits of pollen
Вероятно, характер проявления этих признаков является отражением совокупности многих генотипических и фенотипических черт, проявляющихся на разных стадиях роста и развития растений. Выявленные особенности проявления количественных признаков в потомствах F4, полученных от лучших рекомбинантов, выделенных в F2, с учетом только характеристик проявления признаков мужского гаметофита на искусственно смоделированном фоне, является недостаточным критерием для отбора устойчивых генотипов, сочетающих и высокие стабильные показатели по ряду изученных селекционно ценных признаков.
Выводы
-
1. Результатами исследований показано, что в динамике гибридных поколений F 2 , F 3 и F 4 сохраняется высокая внутрипопуляционная вариабельность по комплексу признаков пыльцы (жизнеспособность свежесобранной пыльцы, длина пыльцевых трубок, устойчивость пыльцы и устойчивость по длине пыльцевых трубок), которые отражают реакцию генотипов на действие высокотемпературного стрессового фактора.
-
2. Установленные рекомбинационные при оценке популяций F 2 различия, выраженные в показателе средней дисперсии по семье (S2) при оценке комплекса признаков пыльцы выявили
значительные различия по частоте встречаемости устойчивых генотипов в популяциях следующих гибридных генераций:
-
- потомства, полученные из рекомбинантов выделенных из комбинаций с широким спектром изменчивости (S2 = 8015), по комплексу признаков пыльцы, в поколениях F 3 и F 4 формируют популяции, в которых выявляется большее число генотипов с равными или значениями показателей выше, чем у их родительских форм;
-
- больше растений с высоким и стабильным проявлением как по комплексу признаков пыльцы, так и количественным отмечается и в популяциях потомств поколений F 3 и F 4 , полученных от рекомбинантов, выделенных из комбинаций, где спектр изменчивости признаков средний (S2 = 4620);
-
- потомства лучших рекомбинантов, выделенных из комбинации где пока-
- Таблица 4. Изменчивость количественных признаков в потомствах F4, полученных от растений F3 с разным уровнем устойчивости к высокой температуре по комплексу признаков пыльцы Table 4. The variability of quantitative traits in F4 offspring obtained from F3 plants with different levels of resistance to high temperature to the complex of pollen traits
-
3. Показано, что потомства, полученные из лучших рекомбинантов F 2 в F 3 и F 4 , не сохраняют высокий уровень устойчивости. По жаростойкости пыльцы фактический сдвиг показателя признака в F 3 относительно F 2 составил 0,92, а в F 4 к F 2 – 0,65. Несколько выше сдвиг показателей в динамике поколений F 2 , F 3 и F 4 от раз-
- ных гибридных комбинаций отмечен по устойчивости пыльцевых трубок – 0,99 (F3/F2) и 1,15 (F4/F2). Наиболее выраженный положительный сдвиг в сторону высоких значений по данному признаку в F3 и F4 относительно F2 выявлен только у трех комбинаций – Л7 x Л305, Л7 x сорт Церос, Л7 x сорт Лидер (R – 1,91, 1,29 и 1,08, соответственно). При проведении отборов по признакам пыльцы с использованием искусственно смоделированных фонов, способность проросших пыльцевых зерен формировать длинные пыльцевые трубки под давлением высокотемпературного стрессового фактора (45°С/8 часов) может служить критерием отбора наиболее устойчивых генотипов.
Высокая изменчивость в популяциях потомств F 4 отмечается и по количественным признакам: число соцветий на растении, число цветков на соцветии, количество завязавшихся плодов на соцветии и число семян в плодах. Различия, выявленные при оценке расщепляющихся популяций F 2 , полученных от разных комбинаций скрещивания по спектру изменчивости, не подтвердились в F 3 , F 4 . Из высокоустойчивых по признакам пыльцы генотипов формируются потомства с большей изменчивостью количественных признаков, в то время как из генотипов со средними показателями получены более выровненные популяции с меньшей изменчивостью этих признаков.
Об авторе:
Milania D. Маkоvеi – Ph D of agricultural sciences, Leading Researcher, Associate Professor
-
• Литература
-
1. Кошкин Е.И. //Физиология устойчивости сельскохозяйственных культур. Москва, 2010. –638 с.
-
2. Тараканов Г.И., Андреева Е.Н., Морев В.В. К вопросу оценки реакции пыльцы на внешние условия //Разработка методов селекции и семеноводства в плодо-овощеводстве: //Сб. научн. тр. ТСХА. М.: 1986. – С.65-69.
-
3. Кравченко А.Н., Лях В.А., Тодераш Л.Г. и др. Методы гаметной и зиготной селекции томатов. /Под редакцией А.А. Жученко // Мн: Кишинев, 1988. – 140 с.
-
4. Маковей М.Д. Применение метода пыльцевой оценки в селекции тепличного томата на устойчивость к стрессовым абиотическим факторам. // Дис… канд. с.-х. наук. 06.01.05. 1992. –197 с.
-
5. Балашова Н.Н., Валеева З.Т., Игнатов А.Н. и др. К вопросу о роли микрогаметофита в адаптации растений к эконише возделывания. // Сельскохоз. биология. – 1994. – №3. – С.59-64.
-
6. Игнатова С.И., Маковей М.Д., Горшкова Н.С. Селекция томата на устойчивость к пониженной освещенности с использованием гаметофитного отбора //Эффективные приемы выращивания овощных культур (Научные труды ВНИИО). Москва,1998. – С.252-256.
-
7. Фотев Ю.В., Юрлова Е.В. Показатель прорастания пыльцы как критерий оценки адаптивной способности гибридов томата. //Сельскохоз. биология. – 1996. – №3. – С.46-51.
-
8. Пивоваров В.Ф. Методические указания по гаметной селекции сельскохозяйственных растений (методология, результаты и перспективы). – Москва (ВНИИСОК), 2001. – 386 с.
-
9. Коваль В.С., Макарова Н.Н. Использование гаметного отбора в селекции на адаптивность // Задачи селекции и пути их решения в Сибири. (Докл. сообщ. генетико-селекционной школы). Новосибирск, 2000. – С.227-231.
-
10. Кильчевский А.В. Генетико-экологические основы селекции растений // Вестник ВОГиС. – 2005. – Т.9. – №4. – С.518-526.
-
11. Бухарова А., Бухаров А. Элементы гаметной и зиготной селекции в практике работ по отдаленной гибридизации томата и перца. // Матер. научно-прак-тич. конф. «Перспективы развития садоводства и овощеводства на Южном Урале». – Уфа, 2005. – С.101-104.
-
12. Anokhina V. The use of gametophytic selection for differentiation of lupin tolerance to stress /V. Anokhina, V. Duksina, I Sauk // Lupin crops: an opportunity for today, a promise for the future. Proceedings of the 13th International Lupin Conference (Poznan, Polan, 6-10 June 2011). – P.285-287.
-
13. Yeun-Kyung Chang, Leslie A. Blischak, Richard E. Veilleux, Muhammad J. Iqbal Effect of temperature on gametophytic selection in a Phalaenopsis F 1 population //Euphytica. 2010. – Vol.171. – Iss. 2. – P.251-261.
-
14. Алпатьев А.В. Мичуринские основы селекции томатов //Сад и огород. – 1947. – №1.
-
15. Тер-Аванесян Д.В. Роль количества пыльцевых зерен цветка в оплодотворении растений. //Труды по прикл. ботанике, генетике и селекции. Л.: 1949. – Т. 28. – Вып.2. – С.119.
-
16. Mulcahy D.L. A correlation between Gametophytic and sporophytic characteristics in Zea mays L. // Science. – 1971. – No171. – Р.1155-1156.
-
17. Голубинский И.Н. //Биология прорастания пыльцы. Киев. 1974. – 464 с.
-
18. Маковей М.Д. Доля влияния родительских компонентов скрещивания на наследуемость устойчивости к температурным стрессам у томатов. //Известия Академии наук Молдовы. Науки о жизни. – 2011. – №1(313). – С.88-94.
-
19. Доспехов Б.А. // Методика опытного дела. – М.: Агропромиздат, 1979. – 416 с.
-
20. Михайленко И.М., Драгавцев В.А. Математические модели в селекции растений. Сообщение I. Теоретические основы идентификации генотипов по фенотипам при отборе в расщепляющихся поколениях. //Сельскохозяйственная биология. – 2013. – №1. – С.26-34.
-
-
• References
-
1. Koshkin E.I. // Physiology of agricultural crop stability. Moscow, 2010. – 638 p. (In Russ.)
-
2. Tarakanov G.I., Andreeva E.N., Morev V.V. On the question of pollen response to external conditions // Development of methods for breeding and seed production in fruit and vegetable farming: // Collection of scientific papers TSHA. Moscow, 1986. – P.65-69. (In Russ.)
-
3. Kravchenko A.N., Lyakh V.A., Toderash L.G. and others. Methods gamete and zygote selection of tomatoes. / Edited by A.A. Zhuchenko // Mn: Chisinau, 1988. – 140 p. (In Russ.)
-
4. Makovei M.D. Application of the pollen assessment method in the selection of greenhouse tomato for resistance to abiotic stress factors. //Dis...PhD of agric. sciences. 01.06.05. 1992. – 197 p. (In Russ.)
-
5. Balashova N.N., Valeeva Z.T., Ignatov A.N. and others. On the question of the role of microgametophyte in plant adaptation to econe-mode cultivation. // J. Agricultural Biology. – 1994. – №3. – P.59-64. (In Russ.)
-
6. Ignatova S.I., Makovei M.D, Gorshkova N.S. Tomato selection for resistance to low light using gametophyte selection // Effective methods of growing vegetable crops (Scientific works of VNIIO). Moscow, 1998. – P.252-256. (In Russ.)
-
7. Fotev Yu.V., Yurlova E.V. Pollen germination rate as a criterion for assessing the adaptive capacity of tomato hybrids. // J. Agricultural Biology. – 1996. – №3. – P.46-51. (In Russ.)
-
8. Pivovarov V.F. Guidelines for gamete selection of agricultural plants (methodology, results and prospects) Moscow (VNIISSOK), 2001. – 386 p. (In Russ.)
-
9. Kovali V.S., Makarova N.N. Using gamete selection in breeding for adaptability // Problems of selection and ways to solve them in Siberia. (Reported by the message of the genetic-breeding school). Novosibirsk, 2000. – P.227-231. (In Russ.)
-
10. Kilchevsky A.V. Genetic-ecological bases of plant breeding // VOGiS-Bulletin. – 2005.
– V.9, №4. – P.518-526. (In Russ.)
-
-
11. Bukharova A., Bukharov A. Elements of gamete and zygote selection in the practice of works on remote hybridization of tomato and pepper. // Collection papers of scientific and practical conf. "Prospects for the development of horticulture and vegetable production in the Southern Urals." Ufa, 2005. – P.101-104. (In Russ.)
-
12. Anokhina V. The use of gametophytic selection for differentiation of lupin tolerance to stress / V. Anokhina, V. Duksina, I Sauk //Lupin crops: an opportunity for today, a promise for the future. Proceedings of the 13th International Lupin Conference (Poznan, Polan, 6-10 June 2011). – P. 285-287.
-
13. Yeun-Kyung Chang, Leslie A. Blischak, Richard E. Veilleux, Muhammad J. Iqbal Effect of temperature on gametophytic selection in a Phalaenopsis F 1 population //Euphytica. – 2010. – Vol.171. – Iss. 2. – P.251-261.
-
14. Alpatyev A.V. Michurin Basics of Tomato Breeding // Garden and Garden. – 1947. – №1. (In Russ.)
-
15. Ter-Avanesyan D.V. The role of the amount of pollen grains of a flower in the fertilization of plants. // Works on botany, genetics and breeding. Leningrad. – 1949. – V.28. – Issue 2. – P.119. (In Russ.)
-
16. Mulcahy D.L. A correlation between Gametophytic and sporophytic characteristics in Zea mays L. // Science. – 1971. – No171. – P.1155-1156.
-
17. Golubinsky I.N. //The biology of germination of pollen. – Kiev, 1974. – 464 p. (In Russ.)
-
18. Makovei M.D. The proportion of the influence of the parental components of the cross on the heritability of resistance to thermal stress in tomatoes. //Journal of Academy of Sciences of Moldova. Life Sciences. 2011. №1 (313). – P.88-94.
-
19. Dospehov B.A. //Methods of experimental work. Moscow: Agropromizdat, 1979. – 416 p. (In Russ.)
-
20. Mikhaylenko I.M., Drahavtsev V.A. Mathematical models in plant breeding. Report I. Theoretical basis for the identification of genotypes by phenotypes in the selection of fissile generations. //Journal Agricultural Biology. – 2013. – №1. – P.26-34. (In Russ.)
Список литературы Внутрипопуляционная вариабельность в потомствах F3 и F4 томата, полученных от рекомбинантов F2, устойчивых к высокой температуре по признакам мужского гаметофита
- Кошкин Е.И. //Физиология устойчивости сельскохозяйственных культур. Москва, 2010. -638 С.
- Тараканов Г.И., Андреева Е.Н., Морев В.В. К вопросу оценки реакции пыльцы на внешние условия // Разработка методов селекции и семеноводства в плодо-овощеводстве: // Сб. научн. тр. ТСХА. М.: 1986. - С. 65-69.
- Кравченко А.Н., Лях В.А., Тодераш Л.Г. и др. Методы гаметной и зиготной селекции томатов. /Под редакцией А.А. Жученко // Мн: Кишинев, 1988. - 140 С.
- Маковей М.Д. Применение метода пыльцевой оценки в селекции тепличного томата на устойчивость к стрессовым абиотическим факторам. // Дис.. канд. с.-х. наук. 06.01.05. 1992. -197 С.
- Балашова Н.Н., Валеева З.Т., Игнатов А.Н. и др. К вопросу о роли микрогаметофита в адаптации растений к эконише возделывания. // Сельскохоз. биология. - 1994. - № 3. - С. 59-64.
- Игнатова С.И., Маковей М.Д., Горшкова Н.С. Селекция томата на устойчивость к пониженной освещенности с использованием гаметофитного отбора//Эффективные приемы выращивания овощных культур (Научные труды ВНИИО). Москва,1998. -С. 252-256.
- Фотев Ю.В., Юрлова Е.В. Показатель прорастания пыльцы как критерий оценки адаптивной способности гибридов томата.//Сельскохоз. биология. -1996. -№ 3. -С. 46-51.
- Пивоваров В.Ф. Методические указания по гаметной селекции сельскохозяйственных растений (методология, результаты и перспективы). -Москва (ВНИИСОК), 2001. -386 С.
- Коваль В.С., Макарова Н.Н. Использование гаметного отбора в селекции на адаптивность//Задачи селекции и пути их решения в Сибири. (Докл. сообщ. генетико-селекционной школы). Новосибирск, 2000. -С. 227-231.
- Кильчевский А.В. Генетико-экологические основы селекции растений//Вестник ВОГиС. -2005. -Т.9. -№ 4. -С. 518-526.
- Бухарова А., Бухаров А. Элементы гаметной и зиготной селекции в практике работ по отдаленной гибридизации томата и перца.//Матер. научно-практич. конф. «Перспективы развития садоводства и овощеводства на Южном Урале». -Уфа, 2005. -С. 101-104.
- Anokhina V. The use of gametophytic selection for differentiation of lupin tolerance to stress/V. Anokhina, V. Duksina, I. Sauk//Lupin crops: an opportunity for today, a promise for the future. Proceedings of the 13th International Lupin Conference (Poznan, Polan, 6-10 June 2011). -P. 285-287.
- Yeun-Kyung Chang, Leslie A. Blischak, Richard E. Veilleux, Muhammad J. Iqbal Effect of temperature on gametophytic selection in a Phalaenopsis F1 population//Euphytica. 2010. -Vol. 171. -Iss. 2. -P. 251-261.
- Алпатьев А.В. Мичуринские основы селекции томатов//Сад и огород. -1947. -№ 1.
- Тер-Аванесян Д.В. Роль количества пыльцевых зерен цветка в оплодотворении растений.//Труды по прикл. ботанике, генетике и селекции. Л.: 1949. -Т. 28. -Вып.2. -С. 119.
- Mulcahy D.L. A correlation between Gametophytic and sporophytic characteristics in Zea mays L.//Science. -1971. -No171. -Р. 1155-1156.
- Голубинский И.Н.//Биология прорастания пыльцы. Киев. 1974. -464 С.
- Маковей М.Д. Доля влияния родительских компонентов скрещивания на наследуемость устойчивости к температурным стрессам у томатов.//Известия Академии наук Молдовы. Науки о жизни. -2011. -№ 1(313). -С. 88-94.
- Доспехов Б.А.//Методика опытного дела. -М.: Агропромиздат, 1979. -416 С.
- Михайленко И.М., Драгавцев В.А. Математические модели в селекции растений. Сообщение I. Теоретические основы идентификации генотипов по фенотипам при отборе в расщепляющихся поколениях.//Сельскохозяйственная биология. -2013. -№ 1. -С. 26-34.
