Внутрипопуляционное разнообразие и внутрипопуляционная дифференциация (на примере пяти видов отряда Carnivora)
")
Автор: Кораблв Мирослав Павлович, Кораблв Павел Николаевич, Кораблв Николай Павлович
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Статья в выпуске: 4, 2014 года.
Бесплатный доступ
Рассматривается внутрипопуляционное разнообразие и дифференциация популяций пяти видов хищных млекопитающих: американской норки Neovison vison, европейской норки Mustela lutreola, лесного хоря Mustela putorius, лесной куницы Martes martes и енотовидной собаки Nyctereutes procyonoides. Показана взаимосвязь данных характеристик. Виды с высоким уровнем разнообразия (американская норка, хорь, енотовидная собака) характеризуются большей внутрипопуляционной структурированностью. Степень различий между группировками не глубока, не зависит от разделяющего их расстояния и определяется сочетанием эндогенных и абиотических факторов.
Хищные млекопитающие, внутрипопуляционное разнообразие, внутрипопуляционная дифференциация
Короткий адрес: https://sciup.org/146116529
IDR: 146116529 | УДК: 599.74:591.4:575.17
Intrapopulation variability and intrapopulation differenciation (an example of five species of the orger Carnivora)
Intrapopulation variability and intrapopulation differentiation of five carnivoran species: American mink Neovison vison, European mink Mustela lutreola L., polecat Mustela putorius L., pine marten Martes martes L., and raccoon dog Nyctereutes procyonoides were investigated. It is shown that species with high level of variability (American mink, polecat, raccoon dog) are characterized by better intrapopulation structuring. The degree of differences between groups is not deep, and does not depend on the distance; it is determined by the combination of endogenic and abiotic factors.
Текст научной статьи Внутрипопуляционное разнообразие и внутрипопуляционная дифференциация (на примере пяти видов отряда Carnivora)
Введение. В морфологии млекопитающих, построенной на изучении краниологического материала, сложилась парадоксальная ситуация: при наличии большого объема исследований по внутривидовой изменчивости чрезвычайно мало работ по внутрипопуляционной дифференциации. В значительной степени это можно объяснить тем, что размеры популяций млекопитающих, особенно с широким радиусом репродуктивной активности, определить очень трудно. В результате, сравнительный анализ выборок часто трактуется как межпопуляционные исследования, что не всегда верно. При этом близко собранные выборки, как правило, объединяются в одну совокупность, хотя материал, собранный на малой географической шкале, позволяет проводить корректный внутрипопуляционный анализ.
Методика. Авторами статьи на протяжении ряда лет собирались черепа промысловых видов млекопитающих в Тверской обл. и на прилегающих к ней территориях. В результате были получены репрезентативные выборки черепов американской норки Neovison vison,
Исследования проведены при поддержке РФФИ (проект № 14-04-97510).
европейской норки Mustela lutreola, лесного хоря Mustela putorius, лесной куницы Martes martes и енотовидной собаки Nyctereutes procyonoides, собранные на удалении друг от друга от 60 до 250 км. Учитывая плотность населения видов и реальную экологическую ситуацию, можно с уверенностью утверждать, что выборки принадлежат одним метапопуляциям указанных видов. Этот материал послужил основой для изучения внутрипопуляционного полиморфизма и дифференциации популяционных группировок отдельных видов с использование морфометрического, фенетического и молекулярногенетического подходов. Всего изучено 828 черепов, в том числе: европейская норка - 103 черепа (две выборки), лесной хорь - 250 черепов (4 выборки), лесная куница - 270 черепов (3 выборки), енотовидная собака - 205 черепов (3 выборки). Результаты исследований по отдельным видам частично опубликованы (Кораблёв и др., 2011, 2012а, 20126, 2013). В этих публикациях подробно описаны методы исследований и дается характеристика материала. Целью настоящей работы является попытка предварительного обобщения данных и выявления некоторых закономерностей внутрипопуляционной дифференциации.
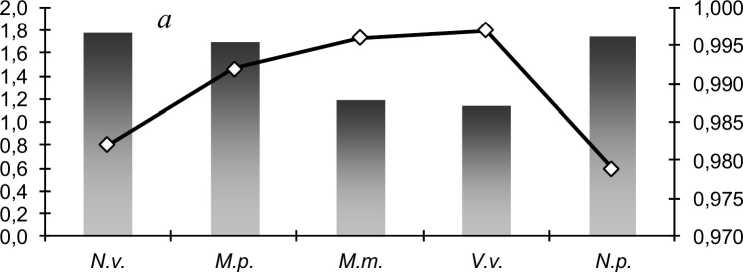
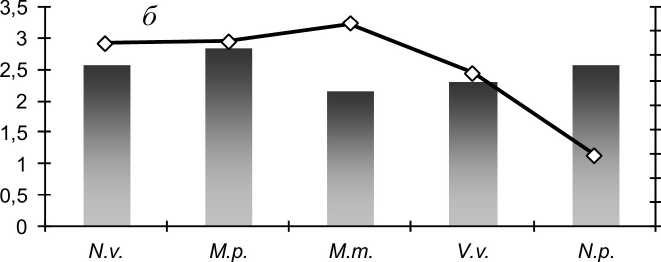
Результаты и обсуждение. Изучение краниометрической изменчивости хищных млекопитающих на основе анализа частот проявления неметрических вариаций (фенов) показывает, что их популяции не мономорфны; внутри них существуют группировки, обладающие морфологическим своеобразием. Прослеживается хорошо выраженная связь степени полиморфизма с уровнем внутрипопуляционной структуры. Сравнение (рис. 1) наглядно показывает, что полиморфные виды отличаются большей внутрипопуляционной структурированностью. Эта тенденция выражена для одонтологических и краниологических признаков, проявляясь ярче в первом случае. Наименьшие различия между районами сбора отмечены для лесной куницы и лисицы, что вполне согласуется с экологическими и морфологическими особенностями видов. В ходе эволюционного развития у куницы выработалось экологически экономное передвижение в условиях заснеженного леса. За счет большой площади опоры при почти такой же массе тела весовая нагрузка на площадь опоры у лесной куницы вдвое меньше, чем у черного хоря (Машкин, 2007). Как следствие этого, средняя длина суточного хода и площадь индивидуального участка у лесной куницы больше, чем у черного хоря (Данилов, Туманов, 1976). Относительно низкая территориальная активность черного хоря, не в последнюю очередь, определяется его синантропизацией. Эта своеобразная экологическая ниша, не характерная для других рассматриваемых видов куньих, обеспечивает хорю хорошую кормовую базу за счет высокой численности грызунов при отсутствии серьезной конкуренции.
1,000
0,990
0,980
0,970
0,960
0,950
0,940
0,930
0,920
0,910
вид
1,000
0,990 о
0,980 “ о
0,970 о
0,960 °
0,950
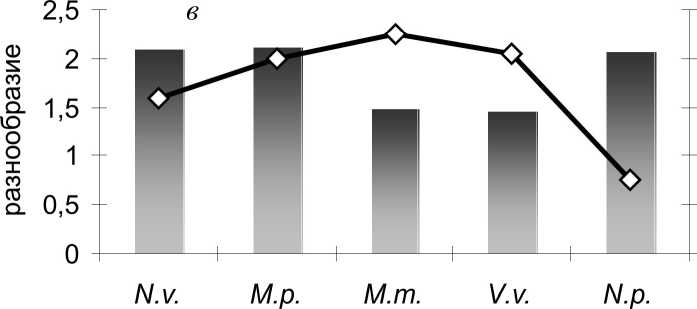
Рис. 1 . Степень внутрипопуляционного разнообразия (ц) и сходства (г) популяционных группировок хищных млекопитающих: а - одонтологические признаки; б - краниологические признаки; в - все признаки. N.v. - американская норка, М.р. - лесной хорь, М.т. - лесная куница, V.v. - лисица, N.p. - енотовидная собака
Тот факт, что выборки черепов енотовидной собаки различаются сильнее, чем лисицы на той же географической дистанции кажется нелогичным с позиции истории формирования группировок, но -116- '
объясним с экологической точки зрения. История появления енотовидной собаки в Тверской обл. началась в 1934 году, когда 25 пар зверей с юга Дальнего Востока были выпущены в Вышневолоцком р-не (Павлов, Корсаков, Лавров, 1974). Всеядность енотовидной собаки и ее способность впадать в зимнюю спячку позволяют зверям обходиться относительно небольшим индивидуальным участком. На Северо-Западе России площадь участка обитания енотовидной собаки составляет от 1 до 12 км2, в то время как у лисицы в зимнее время она варьирует от 20 до 70 км2 (Данилов и др., 1979).
Большая территориальная активность, в особенности, больший радиус репродуктивной активности куницы и лисицы, способствует более интенсивному обмену генетической информацией на большей территории, что и обеспечивает меньшее различие фенофонда между внутрипопуляционными группировками указанных видов в сравнении с другими хищниками на одинаковой географической шкале.
Морфологические различия между внутрипопуляционными группировками прослеживаются и по результатам краниометрического анализа. Как и в случае анализа фенетической изменчивости, степень структурированности популяций у видов различна. Однако, если сопоставить итоги изучения внутрипопуляционной дифференциации видов хищных млекопитающих (табл. 1), то становится очевидным, что нет согласованности не только между результатами, полученными при использовании различных методических подходов (фенетического и морфометрического), но и результатами разных методов статистического анализа пластических признаков.
Таблица 1
Результаты оценки степени внутрипопуляционной дифференциации, полученные разными методами
|
Вид |
Пол |
Дисперсионный анализ* |
Дискриминантный анализ** |
Фенетический анализ (г) |
|
|
min - max |
среднее |
||||
|
Американская норка |
7 |
2,1 |
50,0 - 80,0 |
68,6 |
0,982 |
|
$ |
3,6 |
50,0-100 |
77,5 |
||
|
Европейская норка |
1,0 |
12,5-79,2 |
62,5 |
0,967 |
|
|
? |
0 |
- |
- |
||
|
Черный хорь |
3 |
3,8 |
25,0-91,4 |
65,7 |
0,990 |
|
Лесная Куница |
3 |
0,7 |
80,0-100 |
89,3 |
0,995 |
|
? |
2,0 |
100 |
100 |
||
|
Енотовидная собака |
3 |
2,0 |
38,1 -73,5 |
61,2 |
0,965 |
|
2,0 |
27,3-77,8 |
69,7 |
|||
Примечание. * - среднее число достоверно различающихся признаков, приходящихся на одно парное сравнение; ** - процент корректной классификации.
Отсутствие согласованности между результатами, полученными при использовании различных краниометрических подходов, объясняется разными принципами оценки различий между размерными характеристиками. Некоторые признаки, достоверно различаясь по средним значениям с учетом статистической ошибки, не различаются по
Вестник ТвГУ. Серия "Биология и экология". 2014. № 4 дисперсиям. Трансгрессия признаков отразилась на качестве распознавания выборок в ходе дискриминантного анализа.
Различия в результатах, полученных при использовании фенетических и количественных признаков, более ожидаемы; они формируются разными полигенными системами и определяются большим числом генов, чем дискретные (Майр, 1974). Указанные признаки подвержены сильному модифицирующему влиянию среды (Шмальгаузен, 1983), причем их изменения могут происходить быстро и не всегда иметь микроэволюционное значение, отражая порой неблагоприятные условия онтогенетического развития. Среди фенетических признаков можно выделить относительно моногенные и полигенные, в разной степени реактивные на изменения экологических факторов. Характер формирования разнообразия, а также проявления нарушений онтогенетического развития, позволяют считать, что зубные признаки более моногенны и в большей степени, чем краниологические, генетически детерминированы, что подтверждается данными классической одонтологии (Зубов, Халдеева, 1989).
N.v. M.I. М.р. М.т. М.р.
Вид
Рис. 2 . Связь фенетических и краниометрических показателей внутрипопуляционного разнообразия хищных млекопитающих. Обозначения видов аналогично рис. 1.
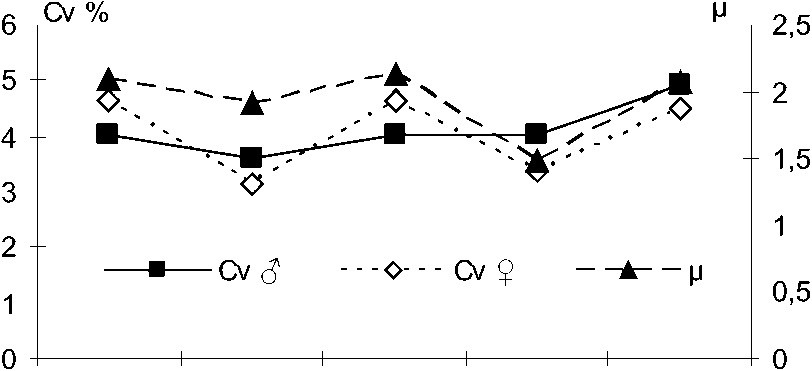
Поскольку нами была показана связь внутрипопуляционного разнообразия с различиями между внутривидовыми группировками по неметрическим вариациям краниологических признаков, представляет значительный интерес обнаружение ассоциаций фенетических признаков с пластическими. Сопоставление результатов изучения внутрипопуляционного разнообразия хищных млекопитающих позволяет отметить тенденцию изменения значений фенетических
Вестник ТвГУ. Серия "Биология и экология". 2014. № 4 характеристик и коэффициентов вариации (рис. 3), причем, если у самцов она лишь слегка обозначается, то у самок выражена вполне отчетливо.
Таким образом, вырисовываются две закономерности: (1) виды, чьё внутрипопуляционное разнообразие (ц) выше, отличаются большей структурированностью популяций (г) и (2) виды с более высокой степенью фенетического разнообразия (ц) более полиморфны по метрическим признакам (Cv). Мы вправе ожидать, что полиморфность и структурированность популяций по метрическим признакам также имеют положительную корреляцию.
Сравнительный анализ степени дифференциации группировок хоря и американской норки свидетельствует, что у хоря высокая степень структурированности по краниометрическим признакам не подтверждается на основе анализа фенетических признаков, а у американской норки различия фенотипов между группировками весьма убедительны. Это может свидетельствовать о том, что различия между группировками американской норки в большей степени, чем у хоря, генетически обусловлены. Различия пластических признаков у последнего, как следует из нашего материала, могут в значительной мере объясняться индивидуальной изменчивостью размеров под влиянием временных экологических факторов. Если это предположение верно, то факторы, оказывающие столь явное влияние на формирование признаков в онтогенезе животных, должны вызывать нарушение стабильности индивидуального развития и отражаться на уровне флуктуирующей асимметрии (Захаров, 1987). Именно это мы и наблюдаем при сравнительном анализе данного показателя у хищных млекопитающих. Среднее значение уровня флуктуирующей асимметрии одонтологических и собственно краниологических признаков у черного хоря занимает максимальное положения (рис. 3). Наивысшее для всех выборок пяти видов млекопитающих значение уровня асимметрии краниометрических признаков равное 41,8 % также отмечено у хоря из Торопецкого р-на. Известно, что асимметрия билатеральных структур минимальна у «средних особей» и максимальна в группах с «крайними» вариантами распределения признаков (Soule, Cuzin-Roudy, 1982). Поскольку именно присутствие крайних вариантов признаков определяют значение коэффициента вариации, то его величина должна быть связана положительной корреляцией с уровнем флуктуирующей асимметрии; эта связь подтверждается при сравнении данных показателей у разных видов (рис. 3).
В то же время имеются данные о том, что у разных животных существует отрицательная корреляция между средней гетерозиготностью полиморфных локусов и флуктуирующей асимметрией меристических признаков. Эту закономерность Л.А. Животовский (1984) считает естественной, полагая, что большая -119- гетерозиготность особей сопутствует большей стабильности их развития. При этом, чем выше гетерозиготность популяции, тем выше концентрация в ней стабильно развивающихся особей. Обнаруженная нами положительная связь, противоречащая приведенным выше примерам, лишь подчеркивает, что разнообразие морфометрических признаков определяется не только генетическим полиморфизмом, но и в значительной степени факторами внешней среды.
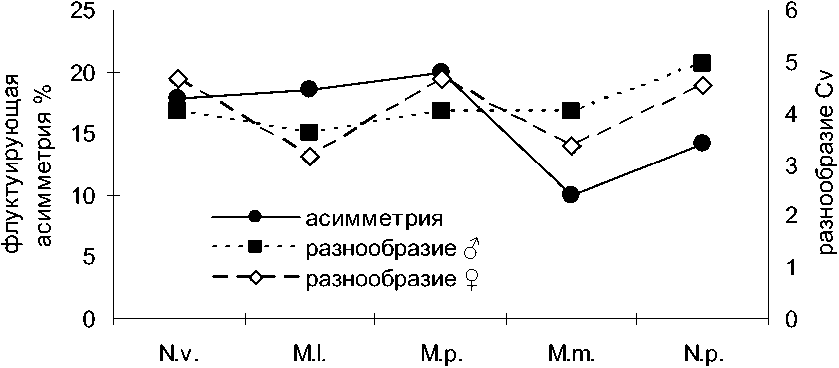
ВИД
Рис. 3 . Связь разнообразия морфометрических признаков с уровнем флуктуирующей асимметрии (средней по группам признаков) билатеральных структур.
Заключение. При формировании фенооблика популяционных группировок биологические особенности видов имеют большее значение, чем длительность их существования. Популяции всех видов в той или иной мере дифференцированы. Степень различий между группировками неглубока, не зависит от разделяющего их расстояния и определяется сочетанием эндогенных и абиотических факторов. Учитывая отсутствие каких-либо экологических или иных барьеров, можно предположить, что внутрипопуляционные группировки не являются устойчивыми и представляют собой временные интеграции полигенных систем (Животовский, 1984), сохраняющие относительную самостоятельность на протяжении жизни немногих поколений. Механизмами, способствующими возникновению внутрипопуляционных группировок, могут быть временные нарушения панмиксии в результате соматического ассортивного спаривания (Ли, 1978 по Егоров, 1983) и зоосоциальное доминирование (Кайданов, 1996). Морфологические различия между популяционными группировками могут не иметь очевидного адаптивного характера, хотя, повышая общепопуляционное разнообразие, являются материалом для адаптациогенеза в условиях постоянной трансформации среды обитания. Наибольшие отличия от остальных группировок отмечены у американской норки Полистово-Ловатской низменности и енотовидной собаки района Дарвинского государственного заповедника. Эти группы особей испытали сильное опосредованное влияние антропогенных факторов: направленную искусственную селекцию на протяжении многих поколений и глобальное изменение ландшафта с возникновением экологических барьеров, ограничивающих панмиксию.
Список литературы Внутрипопуляционное разнообразие и внутрипопуляционная дифференциация (на примере пяти видов отряда Carnivora)
- Данилов П.И., Русаков О.С., Туманов И.Л. 1979. Хищные звери Северо-Запада СССР. Л.: Наука. 163 с.
- Данилов П.И., Туманов И.Л. 1976. Куньи Северо-Запада СССР. Ленинград: Наука. 256 с.
- Егоров Ю.Е. 1983. Механизмы дивергенции. М.: Наука. 172 с.
- Животовский Л.А. 1984. Интеграция полигенных систем в популяциях. Проблемы анализа комплекса признаков. М.: Наука. 183 с.
- Захаров В.М. 1987. Асимметрия животных. М.: Наука. 216 с.
- Зубов А.А., Халдеева Н.И. 1989. Одонтология в современной антропологии. М.: Наука. 231 с.
- Кайданов Л.З. 1996. Генетика популяций. М.: Высшая школа. 320 с.
- Кораблёв М.П., Кораблёв П.Н., Кораблёв Н.П., Туманов И.Л. 2013. Характеристика полиморфизма исчезающей популяции европейской норки (Mustela lutreola, Carnivora, Mustelidae) в районе ЦентральноЛесного заповедника//Зоологический журнал. Т. 9. № 10. С. 1259-1268.
- Кораблев М.П., Кораблев Н.П., Кораблев П.Н. 2012. Морфо-фенетический анализ популяций американской норки (Neovison vison) Каспийско-Балтийского водораздела//Российский журнал биологических инвазий. № 4. С. 36-56.
- Кораблев Н.П., Кораблев М.П., Кораблев П.Н. 2012. Краниометрическая изменчивость енотовидной собаки -Nyctereutes procyonoides Gray (Carnivora, Canidae) Тверской области: от интродуцентов до современных популяций//Бюллетень МОИП. Отделение биологическое. Т. 117. Вып. 1. С. 16-25.
- Кораблев П.Н., Кораблев М.П., Кораблев Н.П. 2011. Оценка состояния популяций хищных млекопитающих в зоне влияния Калининской АЭС на основе анализа их фенофонда//Экология. № 4. С. 272-279.
- Майр Э. 1974. Популяции, виды и эволюция. М.: Мир. 460 с.
- Машкин В.И. 2007. Биология промысловых зверей России. Киров. 423 с.
- Павлов М.П., Корсакова И.Б., Лавров Н.П. 1974. Акклиматизация охотничье-промысловых зверей и птиц в СССР. Ч. II. Киров. Волго-Вятское книжное издательство. 459 с.
- Шмальгаузен И.И. 1983. Пути и закономерности эволюционного процесса. М.: Наука. 360 с.
- Soule M.E., Cuzin-Roudy J. 1982. Allometric variation. 2. Developmental instability of extreme phenotypes//Amer. Natur. V. 120. № 6. P. 765-786.
