Воробьиные птицы из бинагадинских плейстоценовых асфальтов. I. Вводная часть

Автор: Пантелеев А.В., Бурчак-абрамович Н.И.
Журнал: Русский орнитологический журнал @ornis
Статья в выпуске: 112 т.9, 2000 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/140149078
IDR: 140149078
Passerine birds from binagady pleistocene asphalts. I. Introduction
Текст статьи Воробьиные птицы из бинагадинских плейстоценовых асфальтов. I. Вводная часть
Асфальтовые местонахождения с остатками флоры и фауны довольно редки. Все они приурочены к районам выхода нефти на земную поверхность и имеют, чаще всего, четвертичный возраст. Как правило, в подобных местонахождениях органика накапливается на протяжении длительного времени, видовой состав ископаемых очень разнообразен, а сохранность остатков очень хорошая. Нередки находки полных сочленованных скелетов, иногда сохраняются даже связки и мышцы. Кости, пропитанные битумом, приобретают бурый или чёрный цвет.
Наиболее известным асфальтовым местонахождением является Ранчо ла Бреа в южной Калифорнии. Первое сообщение о его ископаемых относится к 1875 г. В дальнейшем, во время раскопок, вскрылись огромные залежи костей. К настоящему времени оттуда извлечено более 2 млн. костей, 100 тыс. насекомых, 40 тыс. моллюсков, большое количество микроорганизмов и растительных остатков, определено более 600 форм растений и животных. Возраст остатков колеблется от 4 до 38 тыс. лет (Stock, Harris 1992). Два других местонахождения (Старунь на Западной Украине и Нижние Кармалки в Татарстане) относительно небольшие. Они пока точно не датированы, но их наиболее вероятный возраст — конец позднего плейстоцена (Лукашевич 1914; Верещагин 1953).
Несколько асфальтовых местонахождений имеется в Восточном Азербайджане. В районе Дивичинского лимана расположены огромные залежи битума с обильной фауной. Н. И. Бурчак-Абрамович, дважды посетивший эти места, собрал там очень много костей. Он оценивает возраст нижнего пласта как верхнеплиоценовый.
На Апшеронском полуострове известны 5 битумных местонахождений с фауной (Кирмаку, Бинагады, Хурдаланы, Бабазанан и о-в Артёма), но исследованы пока только два из них — Кирмаку и Бинагады. В Кирмаку с давних времен велась добыча кира, поэтому к началу палеонтологических раскопок (в 1941 г.) вся площадка была сильно перекопана, и кости извлекались не из пласта коренного залегания. По фауне (бык Bos primi-genius, лошадь, сайгак, волк, лисица) возраст предполагается позднеплейстоценовый (Бурчак-Абрамович, Джафаров 1955), моложе бинагадин-ского. Бинагадинское местонахождение ещё слабо изучено, хотя по обилию остатков и видовому разнообразию, вероятно, не уступает Ранчо ла Бреа и значительно древнее.
Краткая характеристика бинагадинского местонахождения
Рис. 1.
Бинагадинское местонахождение (обведено кружком).
В феврале 1938 студент Ахат Мастанзаде открыл местонахождение четвертичной фауны и флоры у с. Бинагады. Предварительный осмотр местности выявил в Кировых отложениях огромное количество ископаемых остатков прекрасной сохранности. Большое научное значение этого открытия было очевидным, поэтому в декабре того же года начались планомерные раскопки под руководством директора Естественноисторического музея Азербайджанского филиала Академии наук СССР Р.Д.Джафарова. Работы на местонахождении проводились в 1938-1942, 1947-1949 и 1951-1954 гг.
Бинагадинское местонахождение расположено в Азербайджане, почти в самом центре западной половины
Апшеронского полуострова и определяется координатами 40°30' с.ш. и 49°05' в.д. (рис. 1). В настоящее время это северная окраина города Баку. Современный климат полуострова — сухой субтропический с мягкой зимой и сухим жарким летом. Устойчивого снежного покрова не образуется. Годовое количество осадков около 200 мм. Ландшафты полупустынные (Гвоздецкий и др. 1966). На месте выходов бинагадинского кира природа "уныла и однообразна: ни кустика, ни деревца; только изредка верблюжья колючка и аналогичные ей сухолюбивые травы" (Бурчак-Абрамович, Джафаров 1955).
Геология района Бинагадов изучалась с конца XIX в., и этому вопросу посвящено большое число публикаций. Геологическое описание собственно местонахождения наиболее детально сделано И.М.Громовым (1952).
Широко распространено мнение, что животные гибли только в озере, попав в плавающую нефтяную пленку или увязая в топком иле, пропитанном мазутом (Бурчак-Абрамович 1972 и др.). Однако тщательный тафономический анализ захоронения позволяет предполагать, что часть насекомых, птиц и большинство грызунов сносились в озеро рекой и отлагались в её дельте (Громов 1952).
Возраст местонахождения активно обсуждался в конце 1930-х - начале 1950-х, когда проводилось интенсивное изучение органических остатков и геологии. Разногласия были довольно значительными — от минде-ля до "послеледниковья", но многие склонялись к рисс-вюрму (см. обзор: Бурчак-Абрамович, Джафаров 1955).
Исследователи, датировавшие данное местонахождение концом среднего плейстоцена, основывались, главным образом, на следующих об- стоятельствах: 1) костеносный слой залегает непосредственно над морской террасой с Didacna surachanica Andrus., относящейся к верхнехазарскому подгоризонту, который включали в средний плейстоцен; 2) истечение нефти связывалось с "калинской" фазой складкообразования, отвечающей нововалашинской складчатости альпийской системы (средний плейстоцен) (Богачев 1938, 1939); 3) среди млекопитающих присутствуют хазарские элементы: Bos mastanzadei Bur. (близкий к Bos namadicus Falc.), Equus hydruntinus Reg., Dicerorhinus binagadensis (Dz.) (типа носорога Мерка) и др.
Позднее стали появляться данные об абсолютном возрасте подстилающих и перекрывающих отложений. Радиоуглеродное датирование позднехазарских раковин из Дагестана дало возраст более 47 тыс. л.н. (ЛГ-129-А), с р. Урал — 248901170 (ЛГ-148), но последнюю датировку сами авторы считают омоложенной, т.к. было удалено лишь 30% внешнего слоя раковины (Бадинова и др. 1978).
Абсолютные датировки, полученные уран-иониевым методом по раковинам из верхнехазарских отложений Дагестана, содержащих Didacna surachanica, показали возраст 81+2 тыс. л.н. (ЛУ-400 В) по внутренней фракции раковин и 7614 тыс. л.н. (ЛУ-432 В) по полной раковине (Арсланов и др. 1978).
А.А.Свиточ и его соавторы (1985) оценивают возраст нижнехвалын-ских отложений по радиоуглероду в 11-20 тыс. л.н.. Ионий-урановый метод для тех же отложений показал близкий возраст, для верхнехазарских — 75-115 тыс. л.н.; термолюминесцентный метод: хазарская трансгрессия — 90-340 тыс., хвалынская — 15-70 тыс. л.н..
Вообще, радиоуглеродный метод показывает более молодой возраст, чем другие методы. ВАЗубаков, сопоставивший результаты радиоуглеродного, ториево-уранового и термолюминесцентного методов в определении возраста каспийского плейстоцена с трековыми и палеомагнитны-ми данными, пришел к выводу, что наиболее достоверны термолюминесцентные датировки. В приведенной им таблице (Зубаков 1986, табл. 3.2) слои с бинагадинской фауной помещены выше хазарских слоев с Didacna surachanica (датировки: KTL 130±15 тыс. лет, Th/U 8812 тыс. лет) и ниже раннехвалынской террасы (возраст KTL 5316, 6418 тыс. лет), при этом верхняя часть бинагадинских отложений сопоставляется с мустье Волгограда.
Позднейшие исследования позволили В.А.Зубакову уточнить последовательность событий и датировки. Так, мамонтовая фауна культурного горизонта Волгоградской позднемустьерской стоянки оказалась древнее ательских отложений и коррелятна гирканским слоям и первой половине изотопной подстадии 5d (120-113 тыс. л.н.). А начало позднехазарской-гирканской регрессии точно совпадает с г4 экскурс Блейк (113 тыс. л.н.) и серединой изотопной подстадии 5d. Ательская же регрессия коррелятна изотопной подстадии 5с, 106-96 тыс. л.н. (Зубаков 1999).
Для бинагадинской фауны пока можно принять широкий временной интервал 120-96 тыс. л.н., который можно будет сузить при дальнейших исследованиях.
Материал и методика
Птиц из бинагадинского местонахождения изучал сначала П.В.Серебров-ский (в 1939-1941), а после — Н.И.Бурчак-Абрамович (с 1948). Изучение извлекаемого материала шло одновременно с раскопками.
П.В.Серебровский в конце 1939 получил из Естественно-исторического музея АзФ АН СССР для обработки 659 костей птиц, в т.ч. более 150 черепов. Из этого материала ему удалось определить 65 видов (Серебровский 1945, 1948), причем 3 вида оказались новыми, 4 — представленными особыми подвидами, и ещё для 4 указывались отличия от современных форм, предполагающие выделение новых подвидов. Из-за фрагментарности ископаемого материала и неполноты сравнительной коллекции 13 видов были определены условно. Воробьиные оказались представленными 6 формами: Corvus corax (L.), Corvus (?cornix) subsp. nova?, Corvus sp. (ruficollis Less.?), Pica pica (L.), Melano-corypha bimaculata (Menetr.) или M. calandra (L.), Calandrella rufescens (Vieill.).
Н.И.Бурчак-Абрамович, продолживший изучение бинагадинских птиц, увеличил список видов до 108 (Бурчак-Абрамович, Бурчак 1986). Среди воробьиных появились Corvus frugilegus L., Turdus iliacus L., T. merula L. (Бурчак-Абрамович 1949), определённые по цевкам. Однако значительное количество костей воробьиных птиц (особенно мелких) до сих пор оставалось необработанным.
Наша работа основана, главным образом, на материале, собранном после 1939 г. и хранившемся у Н.И.Бурчака-Абрамовича. Кроме того, использовались коллекции костей бинагадинских птиц, переданных в Зоологический институт АН СССР Естественно-историческим музеем АзФ АН СССР в июле 1941, после определения П.В.Серебровским, и в 1950-х — Н.И.Бурчаком-Абрамовичем, после определения им бедренных костей.
Сборы до 1942 г. не этикетированы, послевоенный материал имеет привязку к площадке, квадрату и глубине.
Определение ископаемых костей проводилось с помощью остеологической коллекции Зоологического института Российской Академии наук (ЗИН РАН) и личной коллекции Н.И.Бурчака-Абрамовича.
Для измерения костей использовали промеры, предложенные С.Морер-Шевирэ (Mourer-Chauvire 1975), с некоторыми изменениями (рис. 2). Крупные кости измеряли штангенциркулем, мелкие — под стереоскопическим микроскопом МБС-9. Названия костной морфологии даны по: Baumel (1979), Hamon (1964) и Howard (1929). Статистические расчеты выполнены в программе STATGRAPHICS (версия 5.0); значимость отличий оценивали по критерию Стьюдента (Ивантер, Коросов 1992).
Используемые сокращения названий костей:
|
clav clavicula cmc carpometacarpus cor coracoideum cran cranium fem femur hum humerus mand mandibulae phal phalanx premax premaxillare |
quadr quadratum rad radius scap scapula stern sternum syns synsacrum tib tibiotarsus tmt tarsometatarsus vert vertebra |
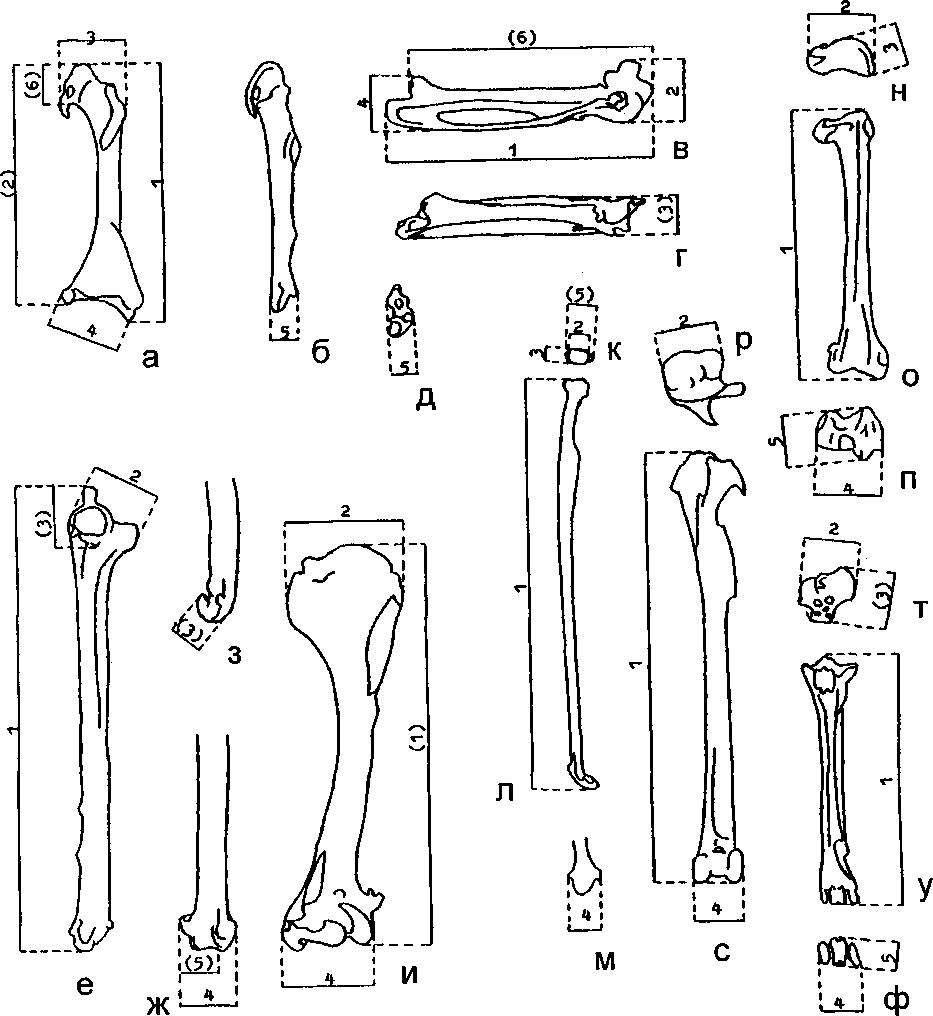
Рис. 2. Используемые промеры костей
(по: Mourer-Chauvire 1975, с изменениями).
Примечание: у промеров, не соответствующих С.Mourer-Chauvire, номера заключены в скобки.
