Воробьиные птицы из бинагадинских плейстоценовых асфальтов. II. Мелкие врановые

Автор: Пантелеев А.В., Бурчак-абрамович Н.И.
Журнал: Русский орнитологический журнал @ornis
Статья в выпуске: 115 т.9, 2000 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/140149089
IDR: 140149089
Passerine birds from binagady pleistocene asphalts. II. Small corvids
Текст статьи Воробьиные птицы из бинагадинских плейстоценовых асфальтов. II. Мелкие врановые
Систематическая часть*
Отряд Passeriformes (Linnaeus, 1758)
Подотряд Passeres (Linnaeus, 1758)
Семейство Corvidae Vigors, 1825
Подсемейство Garrulinae (Boie, 1825)
Род Garrulus Brisson, 1760
Сойка Garrulus glandarius (Linnaeus, 1758)
Современный ареал сойки занимает обширную территорию Палеарктической и Индо-Малайской зоогеографических областей. Внутри вида выделяют более 30 подвидов, объединяемых в 6-8 подвидовых групп. На Апшеронском полуострове в настоящее время встречается кавказская сойка G. g. krynicki Kaleniczenko, 1839.
Для сравнения использованы скелеты современных соек четырех подвидов (по: Степанян 1990) из 3 подвидовых групп: G. g. glandarius (15 экз.), G. g. Iphigenia (4 экз.), G. g. krynicki (1 экз.), G. g. brandtii (4 экз.) и 3 скелета неопределённой подвидовой принадлежности. Кроме того, были промерены и объединены в таблицах и на рисунках с современным материалом 10 фрагментов костей соек из позднеголоценовых пещерных отложений Кавказа, Красноярского и Приморского краёв и Монголии.
Garrulus glandarius assiduus ssp. nov.
Этимология. От assiduus (лат.) — оседлый.
Голотип — ЗИН N РО 4599; левый коракоид (рис., 1, 2); Азербайджан, Бинагады (пл. 1, кв. 1-1, гл. 7 см); поздний плейстоцен.
Материал. Кроме голотипа, имеются правый коракоид (возможно, оба коракоида от одной особи), правая плечевая кость (с обломанными tuberculum ventrale и crista pectoralis), две целые правые локтевые кости от взрослых птиц и проксимальная половина правой локтевой кости молодой птицы.
Размеры голотипа, мм: промер 1 — 32.8; 2 — 30.9; 3 — 6.6; 4 — 8.0; 5 - 2.5; 6 - 5.0.
Описание и сравнение. Coracoideum . В средней части внутреннего края головки коракоида между вершиной акрокоракоида и brachial tu-
Таблица 1. Промеры (мм) coracoideum ископаемых и современных соек Garrulus glandarius
F -G. g. assiduus ssp. n.; GGG - G. g. glandarius; GGI - G. g. Iphigenia;
GGK - G. g. krynicki; GGB - G. g. brandtii
По морфологии коракоида G. g. assiduus ssp. nov. ближе всего к G. g. krynicki и G. g. glandarius.
Бинагадинские коракоиды очень длинные: наибольшая длина кости (промер 1) и длина от int. distal angle до макушки акрокоракоида (промер 2) превышают размеры современных, не перекрываясь. По остальным промерам эти кости лежат у верхнего предела значений для наиболее крупных современных подвидов (glandarius и iphigenia) (табл. 1).
Humerus . На дистальном эпифизе tuberculum supracondylare ventrale более высокий и резче отделен от fossa m. brachialis. С каудальной стороны между crista pectoralis и margo caudalis имеется глубокая ямка. У современных соек эта ямка либо совсем отсутствует (у 9 из 18 экз.), либо очень пологая, слабо выраженная.
Размеры лежат в пределах современных изученных форм вида, наиболее соответствуя G. g. glandarius (табл. 2).
Ulna . Основание olecranon более массивное, чем у современных костей. Размеры ископаемых локтевых костей совпадают с размерами G. g. glandarius (табл. 2).
Замечания. У нас не было возможности сравнить G. g. assiduus ssp. nov. co скелетом G. g. hyrcanus, но промеры шкурок из коллекции Зоологического института РАН показали, что талышская сойка сравнительно мелкая, близка к G. g. brandtii. Бинагадинская сойка по размерам была крупной птицей (как наиболее крупные экземпляры самых крупных подвидов и даже немного крупнее) с относительно короткими крыльями. Полёт такой птицы должен быть более тяжёлым. На это указывает и более массивный локтевой отросток (olecranon), за который тянет оканчивающаяся на нём трёхглавая мышца плеча, ответственная за разгибание (т.е. выпрямление) локтя (Ромер, Парсонс 1992). В связи с этим можно предположить большую оседлость ископаемой формы по сравнению с современными G. g. glandarius, G. g. Iphigenia и G. g. krynicki.
Garrulus glandarius assiduus ssp. nov. могла быть предковой формой для G. g. glandarius и/или G. g. krynicki.
Таблица 2. Промеры (мм) humerus и ulna ископаемых и современных соек Garrulus glandarius
F -G. g. assiduus ssp. n.; GGG - G. g. glandarius; GGI - G. g. Iphigenia;
GGK - G. g. krynicki; GGB - G. g. brandtii
|
Промер |
n M + m |
lim |
|
Humerus |
||
|
1 F |
1 42.0 |
42.0 |
|
GGG |
14 40.64±0.250 |
39.2-42.5 |
|
GGI |
4 42.45±0.155 |
42.0-42.7 |
|
GGK |
2 41.10+0.700 |
40.4-41.8 |
|
GGB |
3 38.77±0.517 |
38.2-39.8 |
|
3 F |
1 4.6 |
4.6 |
|
GGG |
14 4.49±0.025 |
4.4-4.7 |
|
GGI |
4 4.63±0.048 |
4.5-4.7 |
|
GGK |
4 4.68±0.103 |
4.5-4.9 |
|
GGB |
4 4.25±0.029 |
4.2-4.3 |
|
4 F |
1 9.8 |
9.8 |
|
GGG |
14 10.09±0.097 |
9.2-10.5 |
|
GGI |
4 10.38±0.111 |
10.2-10.7 |
|
GGK |
3 10.23±0.240 |
9.9-10.7 |
|
GGB |
4 9.68±0.085 |
9.5-9.Э |
|
Ulna |
||
|
1 F |
2 48.70±0.300 |
48.4-49.0 |
|
GGG |
14 49.29±0.379 |
46.7-52.2 |
|
GGI |
4 51.30±0.493 |
50.2-52.6 |
|
GGK |
1 49.8 |
49.8 |
|
GGB |
4 46.80±0.652 |
45.9-48.7 |
|
2 F |
3 6.90±0.000 |
6.9;6.9 |
|
GGG |
14 6.84±0.050 |
6.5-7.0 |
|
GGI |
4 6.95±0.029 |
6.9-7.0 |
|
GGK |
1 6.9 |
6.9 |
|
GGB |
5 6.40±0.105 |
6.2-6.7 |
|
3 F |
3 6.63±0.033 |
6.6-6.7 |
|
GGG |
14 6.43±0.035 |
6.2-6.6 |
|
GGI |
4 6.73+0.085 |
6.5-6.Э |
|
GGK |
1 6.7 |
6.7 |
|
GGB |
5 5.96±0.068 |
5.8-6.1 |
|
4 F |
2 6.30+0.100 |
6.2-6.4 |
|
GGG |
14 6.13±0.040 |
5.Э-6.4 |
|
GGI |
4 6.35±0.096 |
6.2-6.6 |
|
GGK |
1 5.9 |
5.9 |
|
GGB |
4 5.70±0.108 |
5.5-6.0 |
Наиболее древние остатки соек известны из Puspokfurdo (Румыния) и датируются второй половиной позднего плиоцена (Lambrecht 1933; Ja-nossy 1981) или концом раннего плейстоцена (Janossy 1979). Кости из этого местонахождения определены В.Чапеком как Garrulus aff. glandarius (Janossy 1979). Из нижнего плейстоцена Тюрингии по дистальному фрагменту карпометакарпуса известна Garrulus sp. (Janossy 1965).
Сойки из раннеплейстоценовых местонахождений Чехии и Австрии (Mlikovsky 1996, 1998) определяются уже достаточно уверенно, а из среднего плейстоцена Франции и островов Средиземноморья (Alcover et al. 1992; Mourer-Chauvire 1975) и более поздние морфологически неотличимы от современных. Плейстоценовые (верхнеплейстоценовые?) остатки этого вида с острова Крит морфологически идентичны современным, но по размерам ископаемые кости крыла примерно на 10% короче, хотя размеры костей ноги равны у обеих форм (Weesie 1988).
В восточной части современного ареала самые древние остатки соек найдены в отложениях верхнего(?) плейстоцена Японии (Rich et al. 1986).
Подсемейство Corvinae (Vigors, 1825)
Род Pica Brisson, 1760
Сорока Pica pica (Linnaeus, 1758)
Вид состоит из 12 современных и 1 ископаемого (Р. р. major Janossy, 1972) подвидов. На Апшеронском п-ове сейчас обитает Р. р. pica (Linnaeus, 1758). Ископаемый материал сравнивался с костями 4 современных подвидов: Р. р. pica (5 экз.), Р. р. bactriana (3 экз.), Р. р. camtschatica (1 экз.), Р. р. jankowskii (1 экз.) и 5 скелетами неопределённой подвидовой принадлежности. В таблицах и рисунках использованы промеры 8 позднеголоценовых костей из Красноярского края, объединённые с современными.
Pica pica ssp.
Материал. 1 надклювье, 6 коракоидов (рис., 7-10), 18 плечевых, 13 локтевых, 2 лучевые кости, 2 карпометакарпуса, 5 бедренных костей, 7 голеней, 7 цевок.
Сравнение. Кости бинагадинских сорок морфологически мало отличаются от современных. Premaxillare . Ископаемое надклювье более низкое, имеет более острый конёк и более узкую межноздревую перегородку. По длине оно примерно равно современным.
Coracoideum . На бинагадинских костях facies artic. clavicularis более узкая и выпуклая. По очертаниям головки коракоида ближе всего к Р. р. bactriana. Размеры очень близки к современным костям (табл. 3).
Humerus . Ulna . Radius . Carpometacarpus . Femur . Tibiotarsus . Эти кости (кроме бедренной) в среднем меньше современных. Причём, отличия по всем промерам humerus и некоторым ulna статистически значимы (табл. 3, 4 и 5).
Таблица 3. Промеры (мм) coracoideum и humerus ископаемых (F; Бинагады) и современных (R) сорок Pica pica
|
Промер |
п |
М + т |
Нт |
Р |
|
|
Coracoideum |
|||||
|
1 |
F |
3 |
34.40±0.603 |
33.2-35.1 |
|
|
R |
11 |
34.40±0.283 |
33.0-35.8 |
> 0.05 |
|
|
2 |
F |
4 |
32.63±0.485 |
32.5-33.4 |
|
|
R |
12 |
32.56±0.324 |
31.2-34.5 |
>0.05 |
|
|
3 |
F |
4 |
7.08±0.103 |
6.9-7.3 |
|
|
R |
12 |
6.98±0.064 |
6.7-7.3 |
> 0.05 |
|
|
4 |
F |
3 |
8.13±0.120 |
7.9-8.3 |
|
|
R |
11 |
8.28±0.083 |
7.7-8.6 |
> 0.05 |
|
|
5 |
F |
4 |
2.43±0.155 |
2.0-2.7 |
|
|
R |
13 |
2.39±0.045 |
2.1-2.7 |
>0.05 |
|
|
6 |
F |
6 |
5.70±0.058 |
5.5-5.8 |
|
|
R |
11 |
5.55±0.058 |
5.2-5.8 |
>0.05 |
|
|
Humerus |
|||||
|
1 |
F |
12 |
43.57±0.357 |
41.9-45.2 |
|
|
R |
12 |
45.88±0.560 |
43.1-49.9 |
<0.01 |
|
|
2 |
F |
8 |
13.20±0.134 |
12.7-13.6 |
|
|
R |
12 |
13.89±0.144 |
13.3-14.9 |
< 0.01 |
|
|
3 |
F |
14 |
4.86±0.020 |
4.7-5.0 |
|
|
R |
17 |
5.18±0.047 |
5.0-5.6 |
<0.001 |
|
|
4 |
F |
14 |
10.71±0.083 |
10.2-11.2 |
|
|
R |
16 |
11.20±0.134 |
10.2-12.6 |
<0.01 |
|
Tarsometatarsus . Практически не отличимы от современных цевок Р. р. bactriana. Наибольшие различия наблюдаются с сороками из Приморья: у Р. р. jankowskii гипотарсус повернут более латерально, a proc, medi-alis hypotarsi более массивный. Ископаемые цевки по общей длине и промерам дистального эпифиза соответствуют современным, проксимальный эпифиз значимо меньше. Серебровский (1945, с. 66; 1948, с. 56) по поводу цевки (1 экз.) из своих материалов отмечал, что "она несколько маловата для нормальной птицы".
Замечания. В целом размеры тела бинагадинской сороки соответствовали размерам современных сорок (Р. р. pica, Р. р. bactriana), некоторые параметры костей ног были меньше, а крылья существенно короче.
Сороки неопределенной видовой принадлежности (Pica sp.) найдены в двух позднеплиоценовый местонахождениях Болгарии (Boev 1996).
Ископаемый подвид Р. р. major Janossy 1972, описанный из среднего плейстоцена Чехословакии, известен, кроме того, из верхнего плиоцена
Таблица 4. Промеры (мм) ulna, radius и carpometacarpus ископаемых (F; Бинагады) и современных (R) сорок Pica pica
|
Промер |
п |
М±т |
lim |
Р |
|
1 F |
10 |
Ulna 53.5410.467 |
51.8-55.9 |
|
|
R |
17 |
54.8310.485 |
51.5-57.8 |
>0.05 |
|
2 F |
12 |
7.3910.070 |
6.9-77 |
|
|
R |
19 |
7.7310.060 |
7.3-8.4 |
< 0.01 |
|
3 F |
11 |
7.2810.082 |
6.9-77 |
|
|
R |
18 |
7.5310.072 |
6.9-8.3 |
<0.05 |
|
4 F |
12 |
6.6010.058 |
6.2-6.Э |
|
|
R |
25 |
6.8110.050 |
6.5-7.4 |
<0.05 |
|
5 F |
12 |
5.8610.038 |
5.6-6.1 |
|
|
R |
19 |
6.0310.067 |
5.6-6.6 |
> 0.05 |
|
1 F |
1 |
Radius 46.5 |
46.5 |
|
|
R |
10 |
48.2810.508 |
46.5-51.4 |
> 0.05 |
|
2 F |
2 |
2.6510.050 |
2.6-2.7 |
|
|
R |
11 |
3.0210.055 |
2.8-3.4 |
> 0.05 |
|
3 F |
2 |
2.5510.050 |
2.5-2.6 |
|
|
R |
11 |
2.6810.040 |
2.5-3.0 |
> 0.05 |
|
4 F |
1 |
3.4 |
3.4 |
|
|
R |
13 |
3.6210.060 |
3.2-4.0 |
> 0.05 |
|
5 F |
1 |
3.3 |
3.3 |
|
|
R |
11 |
3.5510.097 |
3.1-4.1 |
> 0.05 |
|
1 F |
2 |
Carpometaca 30.0510.450 |
rpus 29.6-30.5 |
|
|
R |
14 |
32.0310.329 |
30.4-34.9 |
> 0.05 |
|
2 F |
2 |
7.4010.100 |
7.3-7.5 |
|
|
R |
12 |
7.8210.095 |
7.4 -8.6 |
> 0.05 |
|
3 F |
2 |
4.4510.050 |
4.4-4.5 |
|
|
R |
13 |
4.7510.060 |
4.5-5.3 |
>0.05 |
|
4 F |
1 |
6.9 |
6.9 |
|
|
R |
13 |
7.1310.085 |
67-7.9 |
> 0.05 |
|
5 F |
2 |
2.8510.050 |
2.8-2.Э |
|
|
R |
13 |
3.0610.033 |
2.9-3.4 |
>0.05 |
|
6 F |
2 |
27.2010.300 |
26.9-27.5 |
|
|
R |
14 |
28.9510.254 |
27.4-30.9 |
> 0.05 |
Таблица 5. Промеры (мм) femur, tibiotarsus и tarsometatarsus ископаемых (F; Бинагады) и современных (R) сорок Pica pica
|
Промер |
п |
М± т |
lim |
Р |
|
|
Femur |
|||||
|
1 |
F R |
4 12 |
42.75±0.352 42.83±0.389 |
41.9-43.6 40.9-45.4 |
> 0.05 |
|
2 |
F R |
2 12 |
8.20±0.000 7.98±0.105 |
8.2; 8.2 7.5-8.8 |
> 0.05 |
|
3 |
F R |
2 12 |
4.80±0.100 4.88±0.094 |
47-4.9 4.4-5.5 |
>0.05 |
|
4 |
F R |
5 12 |
8.26±0.112 8.25±0.120 |
7.Э-8.6 7.8-9.0 |
> 0.05 |
|
5 |
F R |
5 12 |
6.96±0.087 7.02±0.094 |
67-7.2 67-7.5 |
> 0.05 |
|
Tibiotarsus |
|||||
|
1 |
F R |
2 9 |
67.15±0.650 70.34±0.764 |
66.5-67.8 68.3-74.6 |
>0.05 |
|
2 |
F R |
3 11 |
7.33±0.285 7.45±0.114 |
7.0-7.9 6.8-8.0 |
> 0.05 |
|
4 |
F R |
5 15 |
6.54±0.136 6.69±0.062 |
6.3-7.0 6.3-7.2 |
> 0.05 |
|
Tarsometatarsus |
|||||
|
1 |
F R |
7 18 |
48.84±0775 48.97±0.380 |
47.2-52.9 45.3-51.9 |
> 0.05 |
|
2 |
F R |
7 18 |
6.76±0.081 7.12±0.072 |
6.5-7.0 6.5-77 |
<0.01 |
|
3 |
F R |
5 18 |
7.38±0.073 7.94±0.071 |
7.27.6 7.3-8.4 |
<0.01 |
|
4 |
F R |
6 17 |
4.90±0.137 4.90±0.060 |
4.6-5.S 4.5-5.4 |
> 0.05 |
|
5 |
F R |
7 18 |
2.96±0.043 3.07±0.030 |
2.8-3.1 2.9-3.3 |
>0.05 |
(Csarnota) и нижнего плейстоцена (Villany, Nagyharsany) Венгрии и среднего плейстоцена Франции, Венгрии (Vertessrollos 2) и Австрии (Mourer-Chauvire 1975; Brodkorb 1978; Janossy 1979). Кости этого подвида морфологически не отличаются от современных. Выделение в особую форму основано только на размерах. Длина и некоторые другие (не все) параметры костей Р. р. major в среднем (!) больше современных, однако по абсолютным размерам Р. р. major близка к наиболее крупным современ- ным экземплярам. Птицы из Восточной Европы и Северной Азии несколько крупнее западноевропейских (Cramp, Perrins 1994), что подтверждается и промерами элементов скелета (исключение составляет цевка; табл. 6), поэтому они ещё ближе к Р. р. major. Среди бинагадинских костей тоже встречаются экземпляры, морфометрически близкие к Р. р. major (табл. 7).
Остатки сорок, не принадлежащие к Р. р. major, в Старом Свете известны из среднеплейстоценовых отложений Франции, Израиля, Алтая (?) и Китая (Mourer-Chauvire 1975; Tchernov 1980; Hou 1993; Деревянко и др. 1998; Пантелеев, в печати) и из многих местонахождений Европы и Азии более позднего времени (табл. 8). Все они идентичны современным формам.
Самая ранняя находка сороки в Северной Америке происходит из среднего плиоцена Канзаса. Это проксимальная часть коракоида, отличающаяся от современных костей Pica pica и Р. nutallii менее расширенным и менее изогнутым, почти прямым, внутренним краем головки; птица была меньше самой маленькой современной сороки (Wetmore 1937). Вероятно, это был особый вид (и род?), возможно, предковый для Pica pica и Р. nutallii.
Полный tarsometatarsus из нижнего плейстоцена Техаса по размерам и морфологии уже не отличается от современных костей Pica pica (Miller, Bowman 1956).
Род Corvus Linnaeus, 1758
Подрод Coloeus Каир, 1829
Таблица 6. Соотношение средних размеров (М, мм) некоторых костей современных сорок Pica pica из Западной Европы (WE; Mourer-Chauvire 1975) и Восточной Европы и Северной Азии (ЕЕА)
|
Промер WE |
EEA |
||
|
п М |
n |
M |
|
|
Coracoideum |
|||
|
1 |
13 33.33 |
11 |
34.14 |
|
Humerus |
|||
|
2 |
12 12.78 |
12 |
13.80 |
|
4 |
11 10.87 |
16 |
11.19 |
|
Ulna |
|||
|
2 |
14 7.20 |
19 |
7.72 |
|
4 |
14 5.86 |
25 |
6.80 |
|
Femur |
|||
|
1 |
14 41.76 |
12 |
42.63 |
|
2 |
14 7.81 |
12 |
7.95 |
|
3 |
13 4.69 |
12 |
4.88 |
|
4 |
14 8.09 |
12 |
8.21 |
|
5 |
12 6.46 |
12 |
6.99 |
|
Tarsometatarsus |
|||
|
1 |
13 49.51 |
18 |
48.83 |
|
2 |
13 7.25 |
18 |
7.09 |
|
4 |
13 5.05 |
17 |
4.89 |
|
5 |
12 3.08 |
18 |
3.07 |
Галка Corvus monedula Linnaeus, 1758
Образует 3 подвида (monedula, spermologus, cirtensis). На Кавказе обитает номинативный подвид. На Апшеронском полуострове галка не гнездится, но встречается во время зимних кочёвок (Мустафаев 1968).
Для сравнения использованы 16 скелетов номинативного подвида с Русской равнины, Средней Азии и Казахстана.
Материал. 2 коракоида, 3 плечевые, 2 локтевые, 1 бедренная кость, 4 голени, 1 цевка. Из этих костей 1 коракоид, 1 плечевая, 1 локтевая кость и цевка — от молодой птицы.
Таблица 4. Сопоставление абсолютных размеров некоторых костей сорок Pica pica mayor (Франция, Mourer-Chauvire 1975), Р. pica ssp. (Бинагады) и Р. pica spp. (современные, Восточная Европа и Северная Азия)
Выделены кости, по размерам наиболее близкие к Р. р. major
|
Про- |
Рр. |
Р. р. ssp |
Современные |
|||
|
pica |
bactriana |
jankow |
camtsch. |
|||
|
мер |
major |
Бинагады |
N 4349 |
N N 4977 4978 |
Л/ 4451 |
N 3009 |
Coracoudeum
|
1 |
34.3 |
35,1 |
34.9 |
33.2 |
35.8 |
35.1 |
34.4 |
33161 |
||
|
5 |
2.8 |
II |
ii |
2.0 |
2.4 |
2.3 |
ii |
2.4 |
ii |
2.2 |
|
Humer |
u s |
|||||||||
|
(1) |
46.45 (2) |
Ш |
44.8 |
44,6 |
43.3 |
47.2 |
47.5 |
ii# |
47.7 |
|
|
2 |
13.15 (2) |
13.5 |
13,5 |
13.2 |
12.7 |
14.9 |
13.9 |
Bi |
13.8 |
13.9 |
|
4 |
10.95 (2) |
10,9 |
11.2 |
10,6 |
10.6 |
11.6 |
11.4 |
ITO |
10.9 |
11.5 |
|
Ulna |
||||||||||
|
2 |
7.6 |
ii |
ш |
ii |
ii |
8.0 |
8.0 |
■ |
7.9 |
|
|
4 |
5.8 |
II |
M |
li |
ii |
6.5 |
6.0 |
II |
■1 |
6.0 |
|
Femur |
||||||||||
|
1 |
46.4 |
43.9 |
■ |
42.9 |
41.9 |
45,4 |
44,1 |
42.2 |
40.9 |
43.9 |
|
2 |
8.3 |
8,2 |
M |
8.6 |
8.4 |
7.5 |
7.7 |
8.0 |
||
|
3 |
4.8 |
4.9 |
ii |
5,1 |
5.5 |
4.9 |
4.6 |
5.2 |
||
|
4 |
8.7 |
8,3 |
li |
8.6 |
8.2 |
9.0 |
8.8 |
7.8 |
8.0 |
8.7 |
|
5 |
7.1 |
7.1 |
II |
6.9 |
6.7 |
ill |
li |
6.7 |
6.7 |
6.8 |
|
Tarsometatarsus |
||||||||||
|
1 |
51.6; 53.1 |
48,9 |
52.9 |
48,6 |
49.8 |
51.7 |
49.9 |
50,0 |
47.4 |
49,5 |
|
2 |
7.2; 7.20(2) |
7.0 |
6,9 |
6.9 |
6,9 |
7,4 |
7.4 |
ii |
7.3 |
7.4 |
|
4 |
5.5; 5.07(3) |
5.0 |
ii |
4.9 |
5.1 |
4.8 |
4,7 |
4.8 |
6.1 |
|
|
5 |
3.3; 3.10(3) |
3.1 |
It |
3.0 |
ii |
3.3 |
3,2 |
О |
2.9 |
3.3 |
Таблица 5. Возраст известных находок сороки Pica pica в различных регионах (в скобках - Pica pica major)
|
Регион |
Плиоцен |
Плейстоцен |
Голоцен |
|
2 3 |
1 2 3 |
||
|
Северная Америка |
+? |
+ + |
+ |
|
Европа |
(+) |
(+) (+)+ + |
+ |
|
Западная часть Азии |
+ + |
+ |
|
|
Северная Азия |
+? + |
+ |
|
|
Центральная и Восточная Азия |
+ + |
+ |
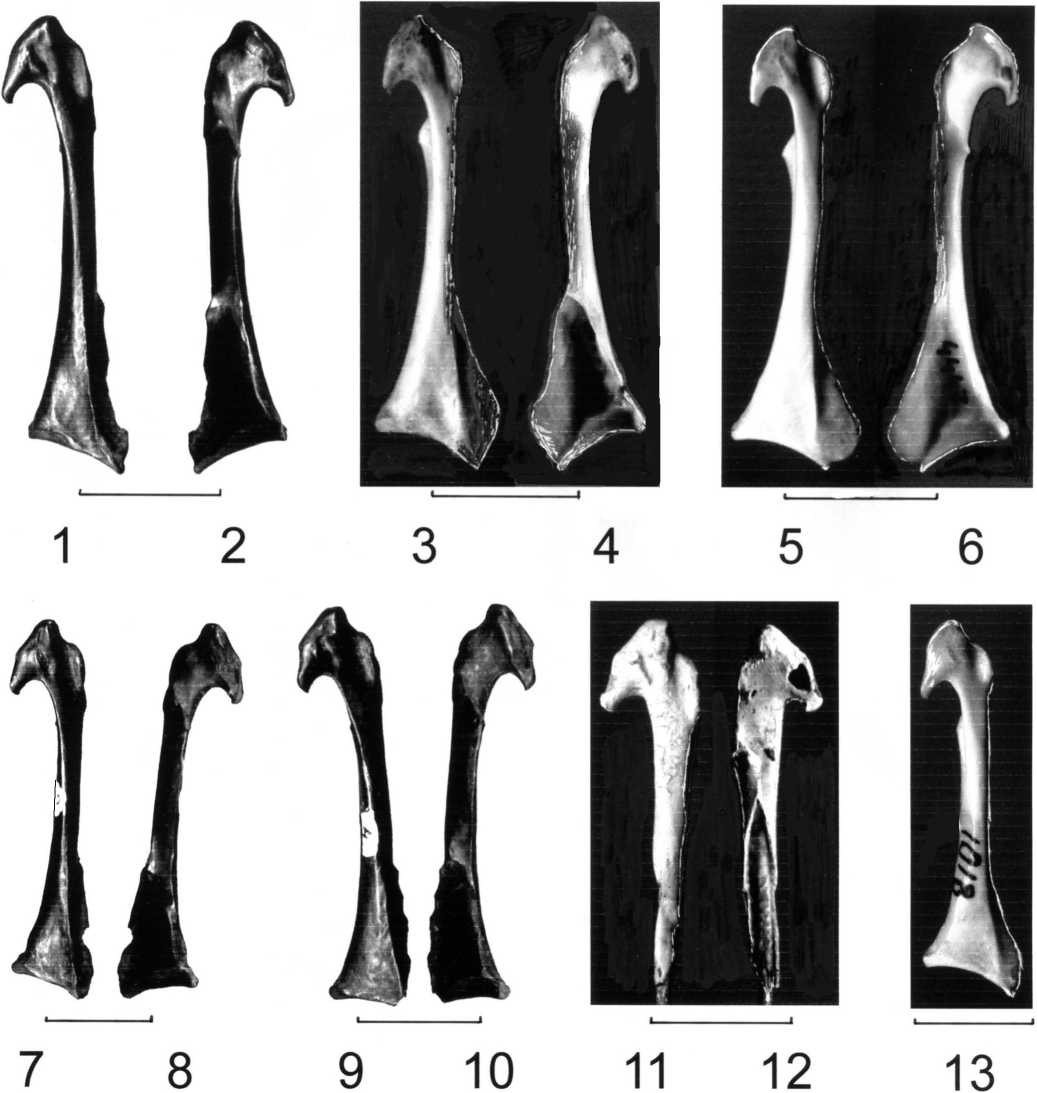
Коракоиды соек Garrulus glandarius (1-6) и сорок Pica pica (7-13).
1-2 - Garrulus glandarius assiduus ssp. nov., голотип,
ЗИН N PO 4599, Бинагады (1 - с краниальной, 2 - с каудальной стороны);
3-4 - Garrulus glandarius glandarius, ЗИН N 3145 (3 - с краниальной,
4 - с каудальной стороны); 5-6 - Garrulus glandarius brandtii, ЗИН N 4447
(5 - с краниальной, 6 - с каудальной стороны); 7-8 - Pica pica ssp., N 2, Бинагады (7 - с краниальной, 8 - с каудальной стороны); 9-10 - Pica pica ssp., N 4, Бинагады (9 - с краниальной, 10 - с каудальной стороны); 11-12 - Pica pica ssp., N 5, Бинагады (11 - с краниальной, 12 - с каудальной стороны);
13 - Pica pica pica, ЗИН N PO 1018, с краниальной стороны.
Таблица 9. Промеры (мм) coracoideum, humerus и ulna ископаемых (F; Бинагады) и современных (R) галок Corvus monedula
|
Промер |
п |
М+т |
Нт |
|
1 F |
1 |
Coracoideum 32.5 |
32.5 |
|
R |
12 |
34.27±0.304 |
32.6-36.3 |
|
2 F |
2 |
31.00±1.100 |
29.9-32.1 |
|
R |
12 |
31.70±0.296 |
30.2-33.4 |
|
3 F |
2 |
6.55±0.050 |
6.5-6.6 |
|
R |
13 |
6.92±0.058 |
6.6-7.2 |
|
4 F |
1 |
8.8 |
8.8 |
|
R |
12 |
9.22±0.072 |
8.9-Э.6 |
|
5 F |
2 |
2.35±0.150 |
2.2-2.5 |
|
R |
12 |
2.60±0.064 |
2.3-3.1 |
|
6 F |
2 |
5.20±0.200 |
5.0-5.5 |
|
R |
13 |
5.48±0.057 |
5.1-5.8 |
|
1 F |
3 |
Humerus 46.57±0.233 |
46.2-47.0 |
|
R |
15 |
46.07±0.303 |
44.2-48.2 |
|
2 F |
3 |
13.80±0.551 |
13.3-14.9 |
|
R |
14 |
14.39±0.157 |
13.5-15.3 |
|
3 F |
3 |
5.27±0.088 |
5.1-5.4 |
|
R |
17 |
5.44±0.055 |
4.Э-5.8 |
|
4 F |
3 |
10.67±0.145 |
10.4-10.9 |
|
R |
17 |
10.96±0.067 |
10.5-11.5 |
|
1 F |
2 |
Ulna 58.30±0.200 |
58.1-58.5 |
|
R |
16 |
58.47±0.412 |
55.8-61.5 |
|
2 F |
3 |
7.65±0.050 |
7.6-7.7 |
|
R |
16 |
7.73±0.046 |
7.4-8.0 |
|
3 F |
3 |
7.10±0.100 |
7.0-7.2 |
|
R |
16 |
7.44±0.056 |
7.1-7.8 |
|
4 F |
3 |
6.90±0.100 |
6.8-7.0 |
|
R |
17 |
7.12±0.040 |
6.Э-7.6 |
Таблица 10. Промеры (мм) femur, tibiotarsus и tarsometatarsus ископаемых (F; Бинагады) и современных (R) галок Corvus monedula
|
Промер |
п |
М±т |
lim |
|
1 F |
1 |
Femur 39.0 |
39.0 |
|
R |
12 |
37.3610.420 |
33.8-39.5 |
|
2 F |
1 |
7.9 |
7.9 |
|
R |
12 |
7.6410.063 |
7.3-8.1 |
|
3 F |
1 |
4.9 |
4.9 |
|
R |
12 |
4.7710.056 |
4.4-5.1 |
|
5 F |
1 |
6.2 |
6.2 |
|
R |
12 |
6.4610.101 |
6.1-7.1 |
|
1 F |
1 |
Tibiotarsus 65.5 |
65.5 |
|
R |
12 |
66.6710.634 |
62.5-70.0 |
|
2 F |
2 |
6.7510.250 |
6.5-7.0 |
|
R |
113 |
6.7110.050 |
6.5-7.1 |
|
4 F |
3 |
6.0010.000 |
6.0-6.0 |
|
R |
14 |
6.1610.041 |
5.8-6.4 |
|
1 F |
1 |
Tarsometatarsus 42.1 |
42.1 |
|
R |
13 |
43.7910.331 |
41.9-46.2 |
|
2 F |
1 |
6.2 |
6.2 |
|
R |
13 |
6.8010.051 |
6.5-7.1 |
|
3 F |
1 |
6.8 |
6.8 |
|
R |
13 |
7.4510.092 |
7.0-8.2 |
|
4 F |
1 |
5.1 |
5.1 |
|
R |
15 |
5.1610.051 |
4.9-5.5 |
|
5 F |
1 |
2.7 |
2.7 |
|
R |
15 |
3.0510.027 |
2.9-3.2 |
Все ископаемые кости морфологически неотличимы от современных. По размерам бинагадинские птицы были несколько мельче современных из Северо-Западного и Центрального районов России (табл. 9 и 10). Но современные галки из этих районов, как показали промеры шкурок из коллекции Зоологического института РАН, в среднем немного меньше кавказских (табл. 11). Следовательно, реальные различия в размерах ископаемых и современных костей будут ещё больше.
Ископаемые галки известны начиная с нижнего плейстоцена. Птица из Крыма ("Corvus cf. monedula"), представленная локтевой и лучевой костями, крупнее современных и, по мнению М.А.Воинственского
(1967), может принадлежать другому (особому) виду. Остатки галки (коракоид, плечевая кость и карпометакарпус) из Венгрии (Janossy 1979) тоже отличаются от современных костей ("Corvus aff. monedula").
Среднеплейстоценовые (Mourer-Chauvire 1975; Tchernov 1962, 1980) и более поздние остатки уже идентичны современным.
Таблица 7. Соотношение размеров галок Corvus monedula из Северо-Западного и Центрального районов России (С) и Кавказа (К).
Промеры: В1 - длина клюва от переднего края ноздри;
В2 - высота клюва на уровне переднего края ноздри;
W1 - длина крыла; W2 - ширина крыла (от сгиба до конца самого длинного второстепенного); TL - длина хвоста; Тт - длина цевки
|
Промер |
Регион |
п |
М+т |
Пт |
Р |
|
В1 |
С |
16 |
21.1410.242 |
19.3-23.0 |
|
|
К |
8 |
22.3010.291 |
21.5-23.8 |
<0.05 |
|
|
В2 |
С |
14 |
12.4710.092 |
11.7-13.0 |
|
|
К |
7 |
12.8010.215 |
12.0-13.5 |
>0.05 |
|
|
W1 |
С |
18 |
231.612.10 |
213-245 |
|
|
К |
8 |
232.412.34 |
225-244 |
> 0.05 |
|
|
W2 |
С |
18 |
151.211.73 |
136-162 |
|
|
К |
8 |
159.9+4.48 |
145-180 |
<0.05 |
|
|
TL |
С |
18 |
122.611.70 |
112-133 |
|
|
К |
8 |
123.111.99 |
116-132 |
> 0.05 |
|
|
Тт |
С |
18 |
42.5010.682 |
36.5-47.5 |
|
|
К |
8 |
43.6010.368 |
42.2-44.8 |
> 0.05 |
