Восстановление фертильности регенерантов тритикале (Triticosekale wittmack), полученных методом искусственного андрогенеза
, полученных методом искусственного андрогенеза")
Автор: Высоцкая И.Б., Кривенко А.А., Барыльник К.Г.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Культура тканей в селекционных технологиях
Статья в выпуске: 3 т.44, 2009 года.
Бесплатный доступ
В культуре пыльников in vitro из донорных сортов гексаплоидной тритикале МАД 1 (зерновой тип) и Благодарный (зернокормовой тип) получали регенеранты, выполняли их микроклональное размножение и размещали в посевах сорта-опылителя для свободного опыления. Показано, что у регенерантов со стерильными пыльниками в полевых условиях образуются жизнеспособные зиготы. Повышенная кустистость гаплоидных форм и микроклональное размножение обеспечивают достаточный выход гибридных семян. Вследствие кумуляции мутаций (сомаклональная изменчивость) и рекомбинаций в гибридной зиготе происходит накопление генотипической изменчивости. Биологический метод восстановления фертильности позволяет эффективно использовать андроклинные регенеранты тритикале в селекционном процессе.
Метод культуры пыльников, андроклинные регенеранты, восстановление фертильности, опыление, особенности селекционного процесса, тритикале
Короткий адрес: https://sciup.org/142133252
IDR: 142133252 | УДК: 633.112.9:631.527
Recovery of fertility in regenerants of Triticosekale wittmack, obtained by the method of artificial androgenesis
The possibility is discussed for biological method of fertility recovery in androclinal regenerants of hexaploid triticale. It was shown that the sterile anthers in regenerants cause the formation of viable zygotes in field conditions as result of free fertilization in body of triticale variety-pollinator. The increased tilling capacity in haploid forms and microclonal cultivation of regenerants in vitro permits to obtain the large amount of hybrid seeds resulted in increasing genotypic variability store, which is connecting with accumulation in hybrid zygote of mutations (somaclonal variability) and recombinations. The biological method permits to use efficiently triticale androclinal regenerants in breeding.
Текст научной статьи Восстановление фертильности регенерантов тритикале (Triticosekale wittmack), полученных методом искусственного андрогенеза
Метод культуры пыльников, который представляет собой один из приемов гаметной селекции и позволяет получать стабильные гомозиготные линии непосредственно из гибридов F1, существенно ускоряя селекционный процесс, пока не находит широкого применения, в частности из-за затруднений на этапе восстановления фертильности андроклинных регенерантов. С этой целью обычно используют колхицинирование, однако у тритикале как для самих колхицинированных регенерантов, так и для полученного потомства характерна значительная генетическая нестабильность (1). После обработки (несмотря на высокую кустистость) регенеранты R1 имеют только 1-2 озерненных колоса, а без обработки колхицином они полностью стерильны. Во втором поколении (R2) семенная продуктивность растений низкая, что обусловлено несбалансированностью числа хромосом. Такую генетическую разнородность связывают с миксоплоид-ностью, возникающей при культивировании клеток in vitro, а также с аномалиями генома исходных растений.
У растений после колхицинирования отмечали значительный летальный и сублетальный эффект (2): 45,0 % регенерантов погибали, 15,0 % — не развивались в полноценные плодоносящие растения, 71,3 % внешне нормальных образцов оказались стерильными при общем выходе фертильных растений 11,2 % и озерненности, которая варьировала в пределах 3,1-32,7 зерна на колос. У большинства форм имелись озерненные и полностью стерильные колосья, что указывает на химерную природу растений. Сообщалось о генетической неоднородности андроклинных дигаплоидов тритикале, низкой озерненности удвоенных гаплоидов: по мнению W. Pohler (1980), цитологическая нестабильность системы полового размножения обусловлена воздействием диплоидизирующего агента (цит. по 2). В других опытах после обработки колхицином наблюдали гибель 86,8 % полученных пшеничноржаных гибридов и невысокую (20,0 %) степень полиплоидизации (3).
Восстановление фертильности пшенично-ржаных амфигаплоидов опылением растений F1 пыльцой родительских видов или селекционных сортов и линий тритикале применяется в селекции с 1939 года (А. Muntzig).
Причем отмечено (А.З. Латыпов, Н.Г. Ходырев, 1970), что искусственное опыление дает семена крайне редко, но небольшое их число обычно формируется при цветении гибридов среди посевов родительских видов, третьего вида или тритикале, то есть в условиях значительного разнообразия пыльцы вида-опылителя, оптимальных для избирательного оплодотворения растений F1 (цит. по 4).
Биологический метод синтеза трехвидовых пшенично-ржаных ам-фидиплоидов, по мнению А.Ф. Шулындина (5), наиболее эффективен в селекции гексаплоидных тритикале. При внутривидовой гибридизации на гексаплоидном уровне изменчивость и разнообразие форм, как правило, значительны и зависят от подбора родительских пар (в этом отношении такие гибриды не отличаются от межвидовых), наблюдается хорошая скрещиваемость родительских форм и нормальная плодовитость гибридного потомства. С помощью этого метода получены высокоурожайные сорта зерновых и кормовых тритикале АД 206, АД 201, АД 209, АД 257, АД 2 и др., характеризующиеся комплексной устойчивостью к болезням, повышенным содержанием белка в зерне и зеленой массе (6).
Целью наших исследований была разработка биологически эффективного метода восстановления фертильности андроклинных регенерантов, позволяющего сохранить широкий спектр генотипической изменчивости, возникающей в культуре пыльников in vitro, для ее использования в селекции тритикале.
Методика. Объектом исследований служили регенеранты, полученные в культуре пыльников in vitro из донорных сортов гексаплоидной тритикале МАД 1 (зерновой тип) и Благодарный (зернокормовой тип). Для увеличения числа растений регенеранты R1 размножали микроклонально (7). Восстановление фертильности стерильных андроклинных регенерантов осуществляли с помощью оригинального биологического метода (экспертной оценкой установлен международный приоритет предложенного способа, принято положительное решение о выдаче патента на изобретение) (8).
Исходные сорта тритикале МАД 1 и Благодарный и микроклоны регенерантов (М-R1 и Б-R1) выращивали в поле (предшественник — черный пар). Для свободного опыления регенеранты размещали в один ряд (площадь питания 45 x 45 см) в массиве смешанного посева сортов тритикале, специально подобранных с учетом сроков наступления фазы цветения. После уборки у исходных сортов и регенерантов (анализировали соответственно по 25 и 5 растений) определяли элементы структуры урожая (9) и проводили статистический анализ данных (10).
Результаты. Ранее мы показали возможность получения широкого спектра генотипов, различающихся по числу хромосом, за счет спонтанной миксоплоидизации в культуре пыльников in vitro (11). Возникшие изменения проявились в значительном фенотипическом разнообразии растений-регенерантов. Достигшие стадии колошения образцы отличались от исходных сортов по ряду признаков — строению колоса (рис. 1, А, Б, Д) и его частей, морфологии пыльников (см. рис. 1, З). Нарушение гаметогенеза у генетически нестабильных форм сказывалось в основном на формировании мужского гаметофита и, как правило, приводило к стерильности полученных растений, о чем свидетельствует внешний вид пыльников в период цветения, а также стерильность всех колосьев, помещенных под изоляторы. Как и у пшенично-ржаных гибридов F1 (4), пыльники регенерантов имели грубую кожистую оболочку, при выходе наружу в фазу цветения не раскрывались и почти не содержали пыльцы. Наряду с лодикулярным у регенерантов был широко представлен овариумный тип цветения, при котором в полностью раскрытых цветках рыльца сохраняют тургор в течение 68
7-9 сут, что служит компенсационным приспособлением, обеспечиваю- щим завязывание семян за счет опыления пыльцой с другого растения.
В колосьях, не помещенных под изоляторы, зерновки формировались от свободного опыления пыльцой фертильных гекса-плоидных сортов и селекционных линий тритикале. По сравнению с донорными сортами (см. рис. 1, В, Е) регенеранты R1 имели более мелкое и морщинистое зерно (см. рис. 1, Г, Ж), большая часть которо-
Элементы структуры урожая у исходных сортов и регенерантов R1 тритикале, полученных в культуре пыльников in vitro
|
Сорт, регенерант |
Число, шт. |
Масса, г |
||||
|
колосков в колосе |
зерен в колосе |
зерен на колосок |
зерен с растения |
1000 зерен |
зерна с растения |
|
|
Благодарный 32,3 62,6 2,0 238 41,0 9,8 Б-R 1 : х 31,7 8,9 0,3 223 25,6 5,7 lim 29-35 1-21 0-1 187-360 – 4,2-6,4 МАД 1 26,4 58,8 2,2 247 51,3 12,7 М-R 1 : х 35,6 22,1 0,6 313 43,5 13,7 lim 19-44 10-28 0-2 267-492 – 12,8-21,2 П р и м е ч а н и е. Описание сортов и регенерантов см. в разделе |
||||||
«Методика». Прочерки означают, что разнородность полученных семян не позволяет определить пределы показателя массы 1000 зерен в соответствии с методикой.
го, несмотря на слабую выполненность, в дальнейшем обладала хорошей всхожестью. При свободном опылении у регенерантов завязывалось от 1 до
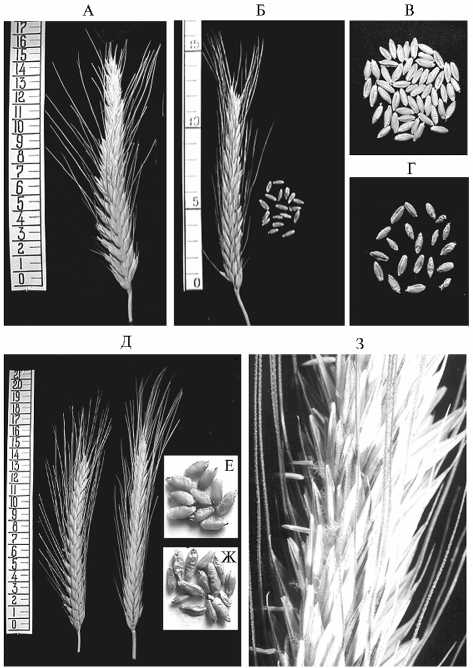
Рис. 1. Внешний вид колоса и зерновок (А-Ж) у растений исходных сортов и андроклинных регенерантов R 1 тритикале и внешний вид пыльников регенеранта М-R 1 в период цветения колоса (З) . Сорт Благодарный: А и В — исходная форма; Б и Г — растение-регенерант. Сорт МАД 1: Д (слева) и Е — исходная форма, Д (справа), Ж и З — растение-регенерант.
28 зерен в колосе, что свидетельствует о значительных нарушениях при формировании не только мужского, но и женского гаметофита (табл.). Средняя озерненность колоска не превышала 0,6 зерна, число зерен в колосе в среднем составило 8,9 и 22,1 шт. Завязываемость семян (от общего числа цветков) достигала 15,5 %, что в несколько десятков раз выше, чем в опытах по восстановлению фертильности пшенично-ржаных амфигаплоидов биологическим методом (3). Благодаря повышенной кустистости регенерантов в полевых условиях (до 3040 побегов) общее число семян с растения достигало 360 шт. (Б-R1) и 492 шт. (М-R1). За счет микро-клонального размножения одного пробирочного образца у регенеранта сорта МАД 1 в результате свободного опыления всего было получено 1567 семян. Факты успешного оплодо- творения пыльцой других растений при неспособности к самоопылению свидетельствуют о генетической неоднородности зародышевой плазмы у регенерантов и наличии яйцеклеток без значительных нарушений.
Вследствие различий по срокам наступления фазы цветения у от- цовских сортов-опылителей продолжительность периода опыления колосьев растений-регенерантов увеличивалась до 25 сут, благодаря чему достигается свободное опыление разновозрастных колосьев. Иными словами, для успешного опыления растений-регенерантов требуется подбор отцовских сортов тритикале, различающихся по срокам наступления фазы цветения и обеспечивающих наличие генетически разнокачественной пыльцы.

Рис. 2. Морфологическое разнообразие колосьев у растений тритикале R2 сорта МАД 1, полученных при свободном опылении андроклинных регенерантов R1.
Растения R2, полученные посредством опыления регенерантов R1, характеризовались широким спектром морфологической изменчивости, выходящей за пределы показателей для родительских форм. Различия проявлялись по высоте растений (от 70 до 185 см), форме куста, числу побегов, наличию воскового налета на листьях. Колосья у растений тритикале R2 имели разную форму, окраску, плотность, длину (рис. 2), различались по наличию остей, их длине и характеру расположения, озерненности (до 135 зерен), форме колосковой и цветковой чешуи. Большая часть растений имела высокую семенную продуктивностью, что косвенно свидетельствует об отсутствии значительных хромосомных нарушений в генетическом аппарате. Следовательно, при свободном опылении за счет успешного оплодотворения с участи- ем наиболее жизнеспособных пыльцевых зерен появляются жизнеспособные зиготы и наблюдается увеличение запасов генетической изменчивости за счет кумуляции в гибридной зиготе рекомбинаций и мутаций, возникающих в процессе культивирования клеток in vitro.
Таким образом, метод культуры пыльников может представлять собой важный источник генотипической изменчивости тритикале благодаря карио- типической разнокачественности популяции культивируемых клеток, которая, по мнению исследователей, служит генетическим механизмом адаптации к условиям искусственной питательной среды (12), и более высокой степени вариабельности гаплоидных тканей по сравнению с диплоидными (13).
Известно, что стабильность генотипа обеспечивается организацией адаптивно наиболее значимых генов в коадаптированные блоки (супергены, кластеры), неслучайным распределением кроссоверных обменов в хромосомах, а также постмейотической элиминацией рекомбинантных гамет и зигот. Внутренний гаметофитный отбор происходит уже в период образования пыльцы, внешний — как при прорастании пыльцы и росте пыльцевых трубок, так и при переносе микрогамет, причем наибольшему давлению элиминирующего отбора подвержены «нетрадиционные рекомбинанты», тогда как гаметы и зиготы, геномы которых соответствуют материнскому компоненту гетерозиготы, получают преимущество (14). Следовательно, метод культуры пыльников не только способствует вовлечению в процесс искусственного отбора огромного числа генотипов, но и исключает давление естественного (преимущественно стабилизирующего) отбора на стадии гаметогенеза.
Биологический метод восстановления фертильности (в отличие от применения колхицина) не только позволяет избежать гибели большей части регенерантов и их мозаичности, эффективно преодолевать стерильность регенерантов, повысить всхожесть и жизнеспособность получаемых семян — он также более экологичный и менее трудоемкий (исключаются этапы обработ- ки колхицином, необходимость выполнения цитологического анализа и т.д.).
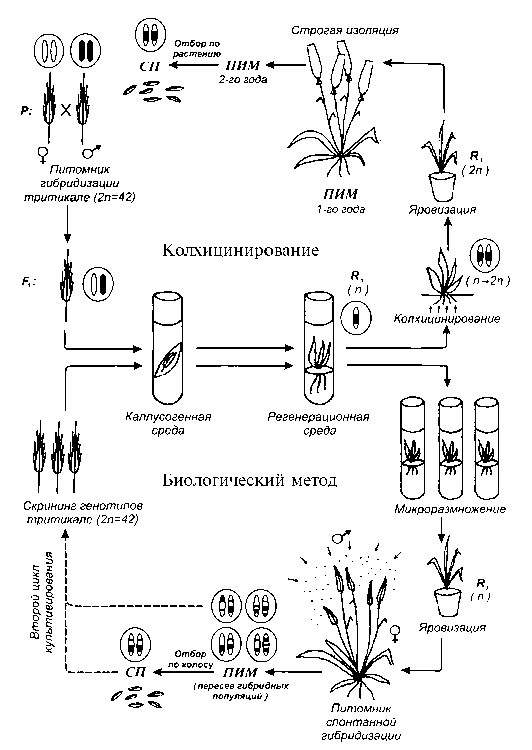
Рис. 3. Схемы использования андроклинных регенерантов тритикале R 1 в селекционном процессе с применением колхицинирова-ния и биологического метода для восстановления фертильности регенерантов. ПИМ — питомник исходного материала; СП — селекционный питомник.
С учетом возможностей применения культуры пыльников in vitro в сочетании с естественным восстановлением фертильности ан-дроклинных регенерантов следует внести ряд изменений в схему селекционного процесса (рис. 3). При колхицинирова-нии для получения константных амфи-диплоидов и исключения их переопыления с другими формами тритикале необходимо, по нашему мнению, в питомнике исходного материала (ПИМ), где в течение 2 лет проводится размножение дигаплоидов, изолировать все колосья (ПИМ 1-го года) и растения в целом (ПИМ 2-го года). Для повышения коэффициента размножения отбор родоначальников в ПИМ 2го года для посева в селекционном питомнике (СП) следует проводить индивидуально по растению. При использовании биологическо- го метода в селекционную схему вводится питомник спонтанной гибридизации, в котором для восстановления фертильности яровизированные рас- тения микроклонально размноженных андроклинных регенерантов высаживают в массиве специально подобранных отцовских форм тритикале (сорт, селекционная линия). Полученные семена F1 высевают в питомнике исходного материала (ПИМ) по растениям, полученное потомство F2 дает начало гибридным популяциям. Учитывая значительное генетическое разнообразие полученных популяций, можно проводить их пересев длительное время. Для повышения адаптивности и стабилизации процессов формирования зерновки необходимо проводить в этих популяциях сортировку семян на крупность и выполненность (12). В питомнике исходного материала отбор родоначальников из гибридных популяций для посева в селекционном питомнике 1-го года следует проводить по колосу. Для сохранения генетической чистоты изучаемых в селекционном питомнике линий тритикале, производных от андроклинных регенерантов, часть растений каждой линии необходимо изолировать. В дальнейшем следует практиковать размножение лучших селекционных линий на изолированных участках.
На основе гибридных популяций тритикале, полученных нами с помощью предложенного биологического метода восстановления фертильности андроклинных регенерантов, созданы высокопродуктивные селекционные линии озимой тритикале разного направления использования, обладающие высокой продуктивностью и повышенной адаптивностью к условиям Центрального Предкавказья (13).
Итак, предложенный нами способ восстановления фертильности ан-дроклинных регенерантов тритикале позволяет шире применять культуру пыльников in vitro в практической селекции.
Л И Т Е Р А Т У Р А
-
1. В н у ч к о в а В.А., Ч е б о т а р е в а Т.М., Г л у щ е н к о Г.И. Получение гаплоидов из пыльников тритикале и фертильного семенного потомства от них. С.-х. биол., 1983, 5: 20-24.
-
2. Е г о р о в а Т.В., Г а л у ш к о Л.Э. Получение дигаплоидных растений тритикале методом колхицинирования. В сб. науч. тр. СНИИСХ: Биотехнология и селекция кормовых культур. Ставрополь, 1990: 24-27.
-
3. С о к о л е н к о Н.И. Использование методов биологического и колхицинирования при получении новых форм тритикале на основе многолетней ржи Державина. В сб. науч. тр. СНИИСХ: Биотехнология и селекция кормовых культур. Ставрополь, 1990: 119-128.
-
4. С е ч н я к Л.К., С у л и м а Ю.Г. Тритикале. М., 1984.
-
5. Ш у л ы н д и н А.Ф. Синтез трехвидовых пшенично-ржаных амфидиплоидов. Генетика, 1970, VI(6): 23.
-
6. Ш у л ы н д и н А.Ф., Г о р б а н ь Г.С. Биологический метод получения гексаплоид-ных тритикале и их цитогенетика. Генетика, 1982, XVIII(6): 977-982.
-
7. К а т а е в а Н.В., Б у т е н к о Р.Г. Клональное микроразмножение растений. М., 1983.
-
8. В ы с о ц к а я И.Б., К р и в е н к о А.А., Н а у м ч и к Д.А. и др. А 01 Н 4/00, С 12 N 5/00 Способ восстановления фертильности андроклинных регенерантов тритикале. Изобретение, заявка ¹ 200729595/13(032227).
-
9. Г о р и н А.П. Практикум по селекции и семеноводству полевых культур. М., 1968.
-
10. А б р а м о в а В. Практикум по генетике. М., 1992.
-
11. В ы с о ц к а я И.Б. Культура пыльников in vitro в селекции тритикале. Автореф. канд. дис. Ставрополь, 2002.
-
12. Б о н д а р е н к о А.М. Клеточные технологии и сомаклональная изменчивость в селекции пшеницы. Физиол. и биохим. культ. раст. 1996, 28(3): 183-193.
-
13. Т у ч и н С.В., А р х и п о в а Л.Н., Н о с о в а Н.Н. и др. Оценка различных селективных систем для отбора на засухоустойчивость в культуре изолированных тканей пшеницы. В сб. науч. тр. НИИСХ Юго-Востока: Биологические основы селекции. Саратов, 1991: 41-48.
-
14. Ж у ч е н к о А.А. Адаптивное растениеводство (эколого-генетические основы). Кишинев, 1990.
-
15. К р и в е н к о А.А. Биология цветения и адаптивная изменчивость пшеницы и тритикале. Автореф. канд. дис. Одесса, 1986.
-
16. В ы с о ц к а я И.Б., Б а р ы л ь н и к К.Г. Перспективы использования андроклинных линий озимой тритикале для повышения урожайного потенциала культуры в условиях Центрального Предкавказья. В сб. науч. тр. Межд. науч.-практ. конф. «Инновации аграрной науки и производства: состояние, проблемы и пути решения». Ставрополь, 2008: 61-64.
ФГОУ ВПО Ставропольский государственный Поступила в редакцию аграрный университет, 1 декабря 2008 года
RECOVERY OF FERTILITY IN REGENERANTS OF Triticosekale Wittmack , OBTAINED BY THE METHOD OF ARTIFICIAL ANDROGENESIS
I.B. Vysotskaya, A.A. Krivenko, K.G. Baryl’nik
S u m m a r y
The possibility is discussed for biological method of fertility recovery in androclinal regenerants of hexaploid triticale. It was shown that the sterile anthers in regenerants cause the formation of viable zygotes in field conditions as result of free fertilization in body of triticale variety-pollinator. The increased tilling capacity in haploid forms and microclonal cultivation of regenerants in vitro permits to obtain the large amount of hybrid seeds resulted in increasing genotypic variability store, which is connecting with accumulation in hybrid zygote of mutations (somaclonal variability) and recombinations. The biological method permits to use efficiently triticale androclinal regenerants in breeding.
