Возможности естественного лесовозобновления в очагах корневой губки

Автор: Чураков Борис Петрович, Битяев Сергей Геннадьевич, Чураков Роман Андреевич
Журнал: Ульяновский медико-биологический журнал @medbio-ulsu
Рубрика: Общая биология
Статья в выпуске: 1, 2017 года.
Бесплатный доступ
Цель. Изучение в течение 5 лет процессов дифференциации деревьев сосны и естественного возобновления леса в очагах усыхания деревьев от корневой губки. Материалы и методы. Исследования проводились в период с 2010 по 2014 г. в 6 действующих очагах усыхания. Очаги имели округлую форму со средним диаметром 41 м. Для сравнения на межочаговом пространстве были заложены контрольные площади размером 35×40 м, сопоставимым по площади очагам усыхания. На каждой контрольной площади и в каждом из очагов усыхания проводился сплошной перечет деревьев с подразделением их на 6 категорий состояния. Средняя площадь очага и контрольного участка составляла 0,14 га, среднее число деревьев на каждом из них - 100 шт. Результаты. В течение 5 лет количество условно здоровых деревьев на контрольных площадях сократилось с 45 штук в 2010 г. до 40 штук в 2014 г. В то же время в очагах усыхания количество условно здоровых деревьев сосны снизилось с 41 дерева в 2010 г. до 14 в 2014 г. В очагах усыхания и особенно в окнах полога может образоваться вполне достаточное количество разновозрастного самосева и подроста различных древесных пород. В обследованных лесных культурах отмечено заметное увеличение количества самосева сосны в очагах усыхания по мере их развития от возникающих (2010 г.) к прогрессирующе действующим (2014 г.). Заключение. В процессе дифференциации деревьев в очагах усыхания сохраняется определенное количество деревьев без признаков поражения болезнью, что указывает на их устойчивость к инфекции. Условно здоровые деревья сосны в очагах корневой губки могут служить хорошим генофондом для естественного возобновления леса. В очагах усыхания по мере их развития формируется определенное количество самосева и подроста древесных пород.
Сосна, корневая губка, дифференциация деревьев, тип леса, очаг усыхания, категория состояния деревьев, естественное возобновление
Короткий адрес: https://sciup.org/14113249
IDR: 14113249 | УДК: 630*443.3 | DOI: 10.23648/UMBJ.2017.25.5257
Possibility of natural reforestation hearth root fungus
Objective: the study within 5 years of the processes of differentiation of pine trees and natural regeneration of forests in the dieback of trees from root sponges. Material and methods research. The research was conducted in the period between 2010 and 2014 in 6 existing dieback. The lesions had a rounded shape with an average diameter of 41 m. For comparison, Melchakova space was laid a control area with similar homes characteristics the size 35×40 m, comparable in size to the dieback. On each control area and each of the foci drying was carried out solid are very rare trees with their division 6 status categories. The average area of the lesion and control area amounted to 0.14 ha, the average number of trees at each of them is 100 PC. Results. Within 5 years the number of healthy trees in the control areas decreased from 45 units in 2010 to 40 in 2014 At the same time in the dieback the number of healthy trees decreased with tree 41 in 2010 to 14 in 2014In dieback and, especially in the Windows of the canopy may produce an ample amount of mixed-age self-sowing and undergrowth of different wood species. In the surveyed forest cultures a marked increase in the number of self-seeding pines in dieback as they evolve from emerging (2010) to progressively existing (2014). Conclusion. In the process of differentiation of trees in the dieback remains a certain number of trees without signs of disease, indicating their resistance to infection. Healthy pine trees in the centers of the root sponges can serve as a good gene pool for natural regeneration of the forest. In the dieback as they progress, formed a certain amount of self-sowing and undergrowth of tree species.
Текст научной статьи Возможности естественного лесовозобновления в очагах корневой губки
Введение. Корневая губка Heterobasidion annosum (Fr.) Bref. является одним из самых вредоносных и наиболее распространенных в мире дереворазрушающих грибов, она поражает многие хвойные и лиственные древесные породы, но лиственные породы страдают от корневой губки меньше, чем хвойные. Поражение древостоев корневой губкой носит очаговый характер.
По С.Ф. Негруцкому, очаг корневой губки – это групповое поражение деревьев с их явно выраженным патологическим ослаблением и усыханием, сопровождающимся ветровалом [1]. В соответствии с «Рекомендациями по защите хвойных пород от корневой губки в лесах европейской части России» очагом корневой губки считается весь выдел, в котором обнаружены пораженные деревья
[2]. Площадь, занятая группой (куртиной) ослабленных, усыхающих и усохших деревьев, называется очагом усыхания. Корневой губкой чаще всего поражаются лесные культуры, созданные на старопахотных землях. В.Г. Стороженко и И.Г. Вишневская выделяют в них очаги инфекции – участки культур, в которых возбудитель болезни обладает патогенностью, позволяющей ему поражать живые деревья [3].
Естественные сосновые древостои поражаются корневой губкой меньше, поскольку они чаще всего формируются как разновозрастные насаждения. По данным многолетних исследований В.Г. Стороженко, в разновозрастных лесах естественного происхождения сосна обыкновенная в возрасте до 40 лет практически не поражается гнилевыми бо- лезнями, а в возрасте от 40 до 80 лет отмечаются лишь единичные поражения [4].
Проведенные некоторыми авторами исследования показывают, что в очагах корневой губки имеется определенное количество деревьев сосны без признаков ослабления, эти деревья могут находиться в любом месте очага [5–7]. Так, на приведенной в монографии С.Ф. Негруцкого схеме многолетнего очага корневой губки внешне здоровые деревья сосны располагаются по периферии очага [1]. О.Н. Гусева также отмечает, что по периметру хронически действующих очагов усыхания сохраняются хорошо развитые деревья [7]. В то же время В.Б. Звягинцев и Г.А. Волченкова указывают на наличие устойчивых к болезни деревьев в центре очага [6].
Такие деревья являются тем генетическим потенциалом, который может дать возможность естественного возобновления леса в очагах корневой губки . Это особенно актуально в связи с тем, что искусственные посадки сосны чаще, чем самосев, подвергаются поражению корневой губкой. Устойчивость лесных культур к поражению корневой губкой может повышаться в связи с постепенно происходящим процессом развития флористического разнообразия на лесокультурной площади [7–12].
Но появление такого разнообразия в чистых культурах в виде живого напочвенного покрова, самосева древесно-кустарниковых пород во многом определяется лесорастительными условиями. Известно, что в сосняках лишайниковых этот процесс формирования флористического разнообразия идет очень медленно. Кроме того, в этих типах леса и сама возможность появления очагов корневой губки минимизирована [1, 6, 13]. Так, С.Ф. Негруцкий отмечает, что в насаждениях сосны низких бонитетов, произрастающих на бедных сухих почвах, распространение гриба практически не бывает значительным [1]. В «Рекомендациях по защите хвойных пород от корневой губки в лесах европейской части России» также указывается на то, что сильнее всего подвержены поражению корневой губкой хвойные породы, произрастающие в наиболее продуктивных типах леса [2].
В более благоприятных местообитаниях процесс формирования флористического разнообразия идет гораздо активнее. Но в этих условиях происходит жесткая конкурентная борьба между видами растений, и культуры сосны быстро заглушаются самосевом и порослью лиственных пород, что требует вмешательства человека в виде проведения рубок ухода.
При наличии в очагах корневой губки деревьев без признаков поражения патогеном можно предположить, что при благоприятных почвенно-климатических условиях и достаточном количестве здоровых семян вполне возможно естественное возобновление сосны. Его успешность будет зависеть от множества факторов биотического и абиотического характера. При этом можно рассматривать два варианта развития ситуации.
Первый вариант характерен для сосняков, растущих на богатых почвах с большим разнообразием подлеска, сопутствующих древесных пород и травянистого покрова: типы леса сложные, разнотравные с преобладанием злаковых растений и др. При развитии очагов корневой губки в этих условиях, следствием чего является распад древостоя, в образующихся окнах полога может появиться самосев таких лиственных пород, как клен остролистный, липа мелколистная, дуб че-решчатый, лещина обыкновенная. При наличии в составе древостоя березы и осины, помимо самосева этих пород, возможно появление также их поросли после рубки таких деревьев. В этих условиях самосев сосны, даже при наличии достаточного количества семян, не всегда сможет сформировать благонадежный подрост из-за высокой конкуренции с дерновыми злаками и лиственным самосевом. В конце концов, в таких лесорастительных условиях на площади очага корневой губки может произойти смена сосны на лиственные породы с единичным участием коренной породы.
В варианте сосняков на более бедных почвах (сосняки-брусничники, черничники, зеленомошники и др.) в окнах полога при распаде древостоя сосны в очагах корневой губки наряду с лиственным может сформироваться сосновый, а иногда и дубовый под- рост. Сохранению такого подроста может способствовать регулярное проведение рубок ухода с целью предотвращения заглушения сосны и дуба другими лиственными породами. Постепенно на таких участках сможет восстановиться сосновый древостой с частичным участием дуба или других лиственных пород.
Поскольку зарастание распадающихся очагов корневой губки лиственными породами является часто происходящим естественным процессом смены пород, то с точки зрения возобновления леса коренными породами этот процесс не представляет значительного практического интереса. Гораздо более важным представляется вопрос возможности появления и сохранения в очагах корневой губки самосева сосны и дуба.
Цель исследования. Изучение в течение 5 лет процессов дифференциации деревьев сосны и естественного возобновления леса в очагах усыхания деревьев от корневой губки.
Задачи исследования:
-
1. Изучить распределение деревьев сосны по категориям состояния в очагах усыхания в течение 5 лет.
-
2. Выявить в очагах усыхания деревья без признаков ослабления как потенциальных производителей самосева сосны.
-
3. Провести учет самосева и подроста в очагах усыхания в течение пятилетнего периода.
Материалы и методы. На протяжении 5 лет (2010–2014 гг.) проводилось изучение процесса дифференциации деревьев по категориям состояния и хода естественного возобновления леса в очагах усыхания в сосняках Кузоватовского лесничества Ульяновской области.
Исследования начались в августе 2010 г. в 21-м выделе (площадью 10,5 га) 11-го квартала Кузоватовского семенного лесничества в 6 действующих очагах усыхания. Очаги имели округлую форму со средним диаметром 41 м. Для сравнения на межочаговом пространстве были заложены контрольные площади размером 35×40 м, сопоставимым по площади очагам усыхания. На каждой контрольной площади и в каждом из очагов усыхания проводился сплошной перечет деревь- ев с подразделением их на 6 категорий состояния (рис. 1). Средняя площадь очага и контрольного участка составляла 0,14 га, среднее число деревьев на каждом из них – 100 шт.
Таксационная характеристика выдела: состав 10С с единичной примесью осины, липы мелколистной, клена остролистного, дуба че-решчатого и березы повислой, культуры сосны 1956 г., класс возраста III, средняя высота 19 м, средний диаметр 18 см, класс бонитета II, тип леса – сосняк бруснично-зеленомошни-ковый, тип условий местопроизрастания – су-борь свежая – А 2 , полнота 0,8.
Ярус подлеска разрежен и представлен рябиной обыкновенной, бересклетом бородавчатым и ракитником русским; травянокустарничковый ярус – брусникой обыкновенной – Vaccinium vitis-idaea L. , земляникой обыкновенной – Fragaria vesca L. , ландышем майским – Convallaria majalis L. , грушанкой круглолистной – Pyrola rotundifolia L ., чиной весенней – Lathyrus vernus Bernh. , ортилией (рамишией) однобокой – Orthilia secunda (L.) House , геранью лесной – Geranium sylvaticum L. , фиалкой собачьей – Viola canina L. , костяникой – Rubus saxatilis L. Общее проективное покрытие травяно-кустарничкового яруса составляло в среднем 45 %.
Ярус зеленых мхов хорошо выражен и представлен Pleurocium schreberi и Dicranum undulatum . Общая замоховелость составляла в среднем 50 %.
В исследуемых очагах и на контрольных площадях были заложены ленточные пробные площади размером 25×2 м по одной на каждом участке. На пробных площадях был проведен учет самосева и подроста сосны и лиственных пород. При этом пневая поросль лиственных пород после выборочной санитарной рубки 2007 г. учитывалась по числу пней с порослью, а каждый корневой отпрыск – как самостоятельное растение. Подлесок из кустарников не учитывался. Количество самосева и подроста во всех вариантах приводится на площадь 50 м2.
Повторное исследование на этих же площадях было осуществлено в августе 2012 г. Таксационная характеристика выдела осталась прежней, за исключением полноты: 0,7.
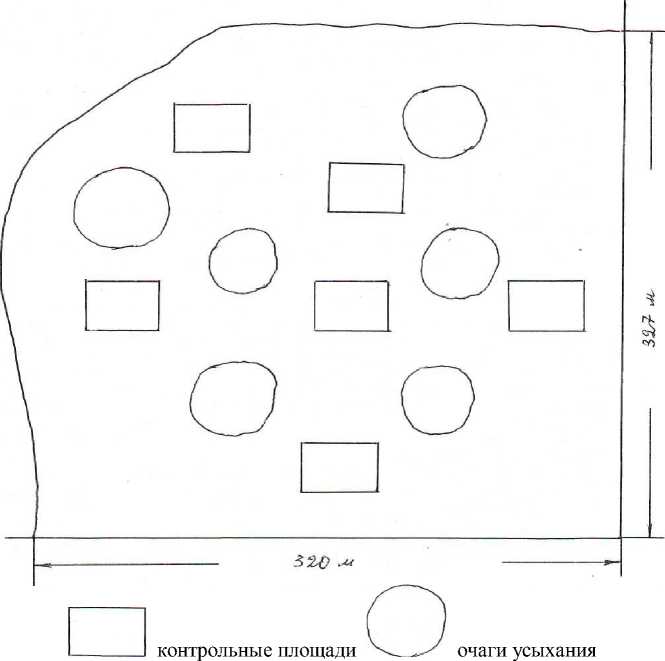
Рис. 1. Схема расположения контрольных площадей и очагов усыхания в 21-м выделе 11-го квартала
Травяно-кустарничковый ярус несколько изменился. Помимо видов, зарегистрированных в 2010 г., появились новые травянистые растения: мятлик узколистной – Poa angusti-folia L ., марьянник луговой – Melampyrum pretense L. , прострел раскрытый – Pulsatilla patens L. Общее покрытие травяно-кустар-ничково яруса составляло в среднем 55 %. Присутствовали отдельные пятна зеленых мхов, замоховелость составляла в среднем 45 %.
В августе 2014 г. проведено последнее изучение дифференциации деревьев и естественного возобновления в очагах усыхания.
Таксационная характеристика выдела изменилась незначительно: средняя высота деревьев сосны стала 20 м, средний диаметр – 21 см, полнота – 0,6.
Травяно-кустарничковый ярус также несколько изменился. Помимо видов, зарегистрированных в 2012 г., появились новые травянистые растения: подмаренник северный – Galium boreale L ., кошачья лапка двудомная –
Antennaria dioica (L.) Gaertn . , но исчезли теневыносливые ортилия однобокая и грушан-ка круглолистная. Общее покрытие травяно-кустарничково яруса составило в среднем 65 %. Присутствовали отдельные пятна зеленых мхов, замоховелость составила в среднем 35 %.
Результаты исследований обрабатывались статистическим методом с определением среднего арифметического X и среднеквадратического отклонения S.
Результаты и обсуждение. Дифференциация деревьев в одновозрастных чистых насаждениях – это естественный закономерный процесс, постоянно происходящий без вмешательства человека во всех лесных насаждениях. На ход этого процесса могут оказывать заметное влияние факторы окружающей среды, а также патогенный фактор. С практической точки зрения определенный интерес представляет вопрос о характере распределения деревьев по категориям состояния в древостоях сосны, пораженных корневой губкой, по сравнению с древостоями вне очагов усыхания и о влиянии этого процесса на ход естественного возобновления леса.
В связи с этим было изучено распределение деревьев сосны на контрольных площадях и в действующих в течение 5 лет очагах усыхания в сосняке бруснично-зеленомошни- ковом. Категории состояния деревьев сосны приведены по А.И. Воронцову и др.: I – деревья без признаков ослабления, II – ослабленные деревья, III – сильно ослабленные деревья, IV – усыхающие деревья, V – сухостой текущего года, VI – сухостой прошлых лет [14]. Результаты представлены в табл. 1.
Таблица 1
|
Вид пробы |
Всего учтено деревьев, шт. |
Учтено деревьев по категориям состояния, шт. |
|||||
|
I |
II |
III |
IV |
V |
VI |
||
|
Результаты исследований 2010 г. |
|||||||
|
Контроль |
103,0±1,8 |
45,0±1,3 |
34,0±1,2 |
21,0±1,2 |
2,0±1,0 |
1,0±0,6 |
- |
|
Очаг |
99,0±1,6 |
41,0±1,4 |
28,0±1,3 |
12,0±1,4 |
10,0±0,9 |
6,0±1,1 |
2,0±0,8 |
|
Результаты исследований 2012 г. |
|||||||
|
Контроль |
95,0±1,5 |
43,0±1,1 |
31,0±1,2 |
11,0±1,2 |
4,0±1,0 |
4,0±0,6 |
2,0±0,5 |
|
Очаг |
90,0±1,7 |
36,0±1,2 |
22,0±1,3 |
10,0±1,4 |
12,0±0,9 |
7,0±1,0 |
3,0±0,6 |
|
Результаты исследований 2014 г. |
|||||||
|
Контроль |
91,0±1,7 |
40,0±1,2 |
30,0±1,4 |
8,0±1,2 |
5,0±0,8 |
5,0±0,6 |
3,0±0,4 |
|
Очаг |
84,0±2,1 |
14,0±1,1 |
16,0±1,4 |
17,0±1,5 |
12,0±1,0 |
14,0±1,1 |
11,0±0,8 |
Распределение деревьев сосны по категориям состояния в действующих очагах корневой губки
Результаты дифференциации деревьев по категориям состояния в 2010 г. показывают, что в сосновых культурах 1956 г., в сосняке бруснично-зеленомошниковом, на контрольных площадях в процессе в среднем было 45 деревьев (44 %) без внешних признаков ослабления, т.е. условно здоровых деревьев. На аналогичной площади в очагах усыхания сохранилось 41 такое дерево (41 %). Следовательно, получены почти сравнимые результаты по условно здоровым деревьям.
В 2012 г. на контрольных площадях условно здоровых деревьев насчитывалось в среднем 43 штуки (45 %), в очагах усыхания – 36 штук (40 %); в 2014 г. – в среднем 40 штук (44 %) и 14 штук (17 %) соответственно.
Таким образом, в течение 5 лет количество условно здоровых деревьев на контрольных площадях сократилось с 45 в 2010 г. до 40 в 2014 г. В то же время в очагах усыхания количество условно здоровых деревьев сосны снизилось с 41 дерева в 2010 г. до 14 в 2014 г. В связи с тем что количество условно здоровых деревьев в очагах усыхания с 2010 по 2012 г. снизилось незначительно (с 41 до 36 штук), действующие в этот временной интервал очаги усыхания можно отнести по классификации И.А. Алексеева [14] к категории формирующихся, в которых в пределах четко наметившихся границ начинают выпадать деревья основного яруса только от болезни. При этом в пределах контура очага сохраняется еще достаточно высокая полнота (0,6–0,8) и наблюдается четко выраженный фон травяно-мохового покрова. Очаги усыхания, действующие в изучаемом выделе до 2010 г. включительно, можно отнести к категории возникающих. С 2012 по 2014 г. в связи со значительным уменьшением количества условно здоровых деревьев эти очаги начинали переходить в категорию прогрессирующе действующих, которые характеризуются наличием полностью сформировавшихся окон и небольшого количества условно здоровых деревьев, расположенных между окнами, или сплошь усохших деревьев в пределах контура очага.
Одновременно с обследованиями древостоев на контрольных площадях и в очагах усыхания проводился учет самосева и под- роста. Для этого были использованы пробные площадки размером 25×2 м. Результаты учета представлены в табл. 2.
Таблица 2
|
Вид пробы |
Количество самосева подроста, шт. |
||
|
хвойные |
лиственные |
всего |
|
|
Результаты учета 2010 г. |
|||
|
Контроль |
5,0±0,8 |
19,0±2,1 |
24,0±2,2 |
|
Очаг |
7,0±1,0 |
17,0±1,9 |
24,0±1,2 |
|
Результаты учета 2012 г. |
|||
|
Контроль |
8,0±1,1 |
12,0±1,6 |
20,0±2,1 |
|
Очаг |
11,0±1,4 |
12,0±1,1 |
23,0±0,5 |
|
Результаты учета 2014 г. |
|||
|
Контроль |
11,0±1,2 |
12,0±1,4 |
23,0±1,8 |
|
Очаг |
16,0±1,0 |
14,0±1,5 |
30,0±0,5 |
Учет самосева и подроста в очагах усыхания
Анализ данных табл. 2 показывает, что в 2010 г. на контрольных площадях сосновый самосев составлял в среднем 21,0 % от общего учтенного количества подроста и был представлен возрастной группой 1–3 лет. Лиственное молодое поколение в среднем составляло 79,0 %, было представлено возрастной группой 1–4 лет и состояло из самосева клена остролистного, дуба черешчатого, березы повислой, пневой поросли липы мелколистной и корневых отпрысков осины.
В очагах усыхания сосновый самосев в среднем составлял 29 %, лиственный подрост – 71 % от общего учтенного количества самосева и подроста. Самосев и подрост были представлены теми же возрастными группами и породным составом, что и на контрольных площадях. Усредненный состав подроста – 3С4Кл2Д1Б ед. Ос и Л.
В 2012 г. сосновый самосев был представлен возрастной группой от одного года до пяти лет. На контрольных площадях он составлял в среднем 40 % от общего числа учтенного подроста. Лиственный подрост был представлен возрастной группой от одного года до шести лет и составлял в среднем 60,0 %.
В очагах усыхания сосновый подрост составлял в среднем 48 %, а лиственный – 52 %. Следовательно, в очагах усыхания относительное количество соснового самосева и подроста увеличилось на 19 %, а лиственного соответственно уменьшилось также на 19 % по сравнению с 2010 г. Состав самосева и подроста в 2012 г. несколько изменился: 4С3Кл2Д1Б ед. Л и Ос.
В 2014 г. на контрольных площадях самосев сосны в среднем составлял 48 %, а лиственный – 52 % от общего количества самосева и подроста. В очагах усыхания по сравнению с контрольными площадями среднее количество самосева и подроста сосны несколько увеличилось и составило 53 % от общего количества самосева и подроста. Количество самосева и подроста лиственных пород в очагах составило 47 %. Сосновый подрост был представлен возрастными группами 1–7 лет, лиственный – 1–8 лет.
Увеличение количества самосева сосны в очагах усыхания в 2012 и 2014 гг. по сравнению с 2010 г. связано, по нашему мнению, с несколькими причинами. В 2010 г. Среднее Поволжье охватила засуха, что сильно сказа- лось на урожае и прорастании семян древесных пород. Кроме того, при развитии очагов усыхания в них со временем появляются окна полога, в которых создаются лучшие условия для возобновления сосны.
Полученные данные дают основание говорить о том, что в очагах усыхания и особенно в окнах полога может образоваться вполне достаточное количество разновозрастного самосева и подроста различных древесных пород. В пересчете на 1 га это будет выглядеть так: в 2010 г. – 4800 шт., в т.ч. сосны – 1400 шт.; в 2012 г. соответственно – 4600 и 2200 шт. и в 2014 г. – 6000 и 3200 шт. Таким образом, можно предположить, что по мере развития очагов усыхания происходит увеличение количества соснового самосева. Однако на самом деле все гораздо сложнее. Дело в том, что судьба этого самосева непредсказуема. Самосев и подрост сосны в очагах хотя и разновозрастный, но в основном представлен растениями в возрасте до 3 лет, что указывает на то, что часть молодого поколения сосны в очагах усыхания может погибать в связи с поражением корневой губкой и по другим причинам. К аналогичным выводам пришли в своих работах И.А. Алексеев [14] и О.Н. Гусева [7].
Заключение. Процессы дифференциации деревьев по категориям состояния идут с большей интенсивностью в очагах усыхания по сравнению с сосновыми древостоями в межочаговом пространстве. По мере активизации болезни интенсивность дифференциации деревьев также усиливается.
В процессе дифференциации деревьев в очагах усыхания сохраняется определенное количество деревьев без признаков поражения болезнью, что указывает на их устойчивость к инфекции. Условно здоровые деревья сосны в очагах корневой губки могут служить хорошим генофондом для естественного возобновления леса.
В очагах усыхания по мере их развития формируется определенное количество самосева и подроста древесных пород. В обследованных лесных культурах отмечено заметное увеличение количества самосева сосны в очагах усыхания по мере их развития от возникающих (2010 г.) к прогрессирующе действующим (2014 г.). По нашему мнению, это связано как с погодными условиями (засухой 2010 г.), так и с биоэкологическими особенностями сосны, которая лучше возобновляется в окнах полога, появляющихся в процессе развития очагов усыхания.
Достоверно невозможно утверждать, что весь подрост сосны окажется в будущем благонадежным, поскольку часть его погибает по различным причинам.
Список литературы Возможности естественного лесовозобновления в очагах корневой губки
- Негруцкий С.Ф. Корневая губка. М.: Агропромиздат; 1986. 196.
- Рекомендации по защите хвойных пород от корневой губки в лесах европейской части России. Пушкино; 2001. 12.
- Стороженко В.Г., Вишневская И.Г. Диагностика пораженных сосновых культур корневой губкой. В кн.: Защита леса от вредителей и болезней. М.; 1980: 192-201.
- Стороженко В.Г. Эволюционные принципы поведения дереворазрушающих грибов в лесных биогеоценозах. Тула: Гриф и К; 2014. 184.
- Чураков Б.П., Маслов В.Д., Чураков Р.А. Влияние корневой губки на древесную продукцию сосны в очагах болезни. Вестник УГСХА. 2013; 4: 56-66.
- Звягинцев В.Б., Волченкова Г.А. Трансформация патогенеза корневой губки при интенсификации лесного хозяйства. В кн.: Грибные сообщества лесных экосистем. Т. 4. М.; Петрозаводск; 2014: 15-25.
- Гусева О.Н. Поражение корневой губкой чистых и смешанных культур сосны в условиях экологического стресса: дис. … канд. с.-х. наук. Йошкар-Ола; 2011. 230.
- Алексеев И.А. Научные основы лесохозяйственных мер борьбы с корневой губкой в лесах Полесья и лесостепи УССР: автореф. дис. … д-ра с.-х. наук. Л.; 1974. 35.
- Алексеев И.А., Кусакин А.В., Коток О.Н. Определение показателей биоразнообразия в насаждениях как объективных факторов устойчивого ведения лесного хозяйства. Международная конференция «Рациональное использование лесных ресурсов». Йошкар-Ола: МарГТУ; 2001: 36-38.
- Ахметов В.М. Корневые гнили сосны обыкновенной (Pinus sylvestris L.) в Нижнем Прикамье и меры по снижению их вредоносности: автореф. дис. … канд. с.-х. наук. Йошкар-Ола; 2007. 22.
- Василяускас А.П. Корневая губка и устойчивость экосистем хвойных лесов. Вильнюс; 1989. 176.
- Woodword S., Steinlid J., Karjalainen R. Heterobasidion annosum: Biology, Ecology, Impact and Control. CAB International. New York; 1998. 589.
- Федоров Н.И. Корневые гнили хвойных пород. М.: Лесная промышленность; 1980. 160.
- Чураков Б.П., Алексеев И.А., Чураков Д.Б. Лесная фитопатология. Ульяновск: УлГУ; 2013. 476.
