Возможные механизмы изменения функциональных свойств полоски диафрагмы мыши при белковой сенсибилизации

Автор: Теплов Александр Юрьевич, Фархутдинов Альберт Мансурович, Теплов Олег Вадимович, Гришин Сергей Николаевич
Журнал: Гений ортопедии @geniy-ortopedii
Рубрика: Оригинальные статьи
Статья в выпуске: 1, 2012 года.
Бесплатный доступ
Исследованы механизмы влияния аллергической перестройки организма на сократительную функцию полоски диафраг- мы (ДМ) мыши. Показано, что при белковой сенсибилизации (БС) возрастает сила сокращения (Рос), вызванного холино- миметиком карбахолином (Кx), у ДМ in vitro и снижается неквантовая секреция (НС) ацетилхолина (Аx) в зоне концевой пластинки. Увеличение Рос на Кx является следствием повышения чувствительности постсинаптической мембраны мы- шечных волокон к холиномиметику, что обусловлено снижением уровня НС Аx в зоне концевой пластинки. Экзогенная АТФ также увеличивает Рос и снижает НС АХ полосок ДМ и интактных, и сенсибилизированных мышей. Вектор дина- мики Рос мышцы на Кx при всех вариантах экспериментальных воздействий коррелирует с изменением НС Аx. Однако степень этих изменений у мышцы сенсибилизированного животного менее выражена, чем у несенсибилизированного. Высказывается предположение, что меньшая изменчивость функциональных свойств, вызванная АТФ, у ДМ сенсибили- зированных мышей в сравнении с контролем является свидетельством развития механизмов резистентности к внешним нагрузкам у дыхательных мышц, которая появляется при бронхоспастическом синдроме и бронхиальной астме.
Диафрагмальная мышца, сократительные свойства, неквантовая секреция, белковая сенсибилизация, атф, мышь
Короткий адрес: https://sciup.org/142121508
IDR: 142121508
Possible mechanisms of changing in the functional properties of a mouse diaphragmal muscle strip for protein sensitization
The mechanisms of influencing the organism allergic reorganization on the contractile function of a mouse diaphragm (MD) muscle strip have been studied. In has been demonstrated that in case of protein sensitization (PS) the increase of the power of the contraction (PC) produced by cholinomimetic carbacholinum (ChC) in MD in vitro is observed, as well as the decrease of acetylcholine (ACh) nonquantum secretion (NQS) in the zone of end plate. PC increase under ChC is a consequence of the cholinomimetic sensitivity increase of muscle fiber postsynaptic membrane due to the decrease of ACh NQS in the zone of end plate. Exogenous ATP also increases PC and decreases ACh NQS of MD in both intact and sensitized mice. The vector of muscle PC dynamics under the cholinomimetic correlates with ACh NQS for all the variants of experimental influence. However, the extent of these changes in the muscle of a sensitized animal is less pronounced than in a non-sensitized animal. It is speculated that the lesser variability of functional properties caused by ATP in MD of sensitized mice compared with control is an evidence of developing mechanisms of the resistance for external loads in respiratory muscles, which appears for bronchospastic syndrome and bronchial asthma.
Текст научной статьи Возможные механизмы изменения функциональных свойств полоски диафрагмы мыши при белковой сенсибилизации
Аллергическая перестройка организма, изменяя функциональные свойства поперечнополосатых мышц, затрагивает холинергические механизмы их возбуждения [1]. Данное предположение было подтверждено нашими исследованиями in vitro , выполненными на «быстрых» и «медленных» мышцах голени мыши [4, 5, 7], в которых было показано изменение характеристик сокращения поперечнополосатых мышц мыши на холиномиметик карбахолин и неквантовой секреции ацетилхолина в зоне концевой пластинки при белковой сенсибилизации.
Ранее на тех же объектах и в тех же экспериментальных моделях была показана способность экзогенной АТФ обратимо изменять сократительную функцию и величину неквантовой секреции ацетилхолина [9, 11]. Одним из объяснений механизмов обнаруженного действия стало предположение, что АТФ влияет на холиноопосредованные процессы возбуждения мышечных волокон (МВ).
Диафрагма, являясь основной дыхательной мышцей, изменяет свои сократительные свойства в условиях нарушенной бронхиальной проводимости при заболеваниях, имеющих в своей основе аллергический компонент, таких как бронхоспастический синдром и бронхиальная астма [12, 13, 16].
Принимая во внимание тот факт, что АТФ, являясь эндогенным модулятором функции поперечнополосатых мышц [9], одновременно способна участвовать в механизмах генерации иммунного ответа [14], мы исследовали влияние экзогенной АТФ на функциональные свойства диафрагмы in vitro у несенсибилизированных и сенсибилизированных чужеродным белком мышей.
Цель работы: изучение влияния экзогенной АТФ на сократительные свойства и уровень неквантовой секреции Ах в зоне концевой пластинки полосок диафрагмы интактных и сенсибилизированных мышей.
МАТЕРИАЛЫ И МЕТОДЫ
Исследование сократительной функции поперечнополосатой мышцы экспериментального животного проводилось методом регистрации со -кратительных свойств in vitro , в изометрическом режиме с помощью фотоэлектрического преобразо-вателя [2].
Эксперименты проводились на препаратах изолированных мышц мыши — диафрагме. Препарат мышцы помещался в термостатируемую ванночку и одним концом крепился к датчику фотоэлектрического преобразователя. Другой конец жестко фиксировался в ванночке. Мышца при постоянной перфузии физиологическим раствором типа Кребса растягивалась в течение 20-30 минут с силой, достаточной для достижения изометрического режима сокращения. Сокращение мышцы вызывалось кар-бахолином (Кх) в конечной концентрации 2x10-4 М, раствор которого добавлялся в ванночку микродозатором при остановленной перфузии. Кривая сокращения регистрировалась самописцем Н-327–1. Для оценки сократительных свойств полоски скелетной мышцы было использовано 3 параметра, позволяющих достаточно объективно отражать функциональное состояние исследуемой полоски мышцы: 1) время развития максимального напряжения (СТ); 2) сила одиночного сокращения (Рос); 3) скорость сокраще- ния (Vос), определяемая отношением Рос / СТ. Сила сокращения определялась в милиграммах; временные характеристики — в секундах; скорость сокращения — отношением мг/сек.
Неквантовую секрецию Ах измеряли с помо-щью стеклянных микроэлектродов (сопротивлением 8–12 МΩ, заполненных 2,5 М KCl) [10]. Для определения ее величины сначала армином устранялось действие ацетилхолинэстеразы, после чего на мышцу в течение 8-12 минут апплицировал-ся блокатор Н-холинорецепторов d-тубокурарин (ТБК) (10–5 М). Разница значений мембранного потенциала (МП) до и после аппликации ТБК соответствует величине неквантовой секреции Ах (Н-эффект).
Методы сенсибилизации мышей яичным альбумином с гелем гидроокиси алюминия и контроля сенсибилизации подробно описаны в работе И. С. Гущина и соавт. [3].
Влияние на диафрагму АТФ (перфузия в концентрации 1x10-4 Мв течение 5 минут) изучалось сравнением показателей сокращения до и после перфузии мышцы раствором, содержащим изучаемое вещество в заданной молярной концентрации; время действия его на мышцу определялось длительностью перфузии.
РЕЗУЛЬТАТЫ
Изучение сократительной функции полоски диафрагмы мыши показало, что карбахолин в концентрации 2 x 10-4 М вызывает ее сократительный ответ со следующими характеристиками: СТ — 11,3 ± 0,63 сек; Рос — 335,2 ± 93,47 мг; Vос — 31,0 ± 1,7 мг/сек. Белковая сенсибилизация изменяет эти показатели: СТ–13,71 ± 0,42 сек, Рос — 469,83 ± 86,78 мг (р < 0,01), Vос — 32,08 ±0,89 мг/сек, приводя к достоверному увеличению силы сокращения.
Изучение неквантовой секреции Ах в диафрагме показало, что МП покоя, изначально составляющий (-70,7) ± 1,9 мВ (n = 150), в присутствии ТБК возрастал до (–75,9) ± 0,7 мВ (n = 150). Таким образом, Н-эффект в контроле составлял 5,2 ± 0,4 мВ (n = 150). В условиях белковой сенсибилизации МП покоя, изначально составляющий (–70,0) ± 1,5 мВ (n = 150), в присутствии ТБК возрастал до (–74,4) ± 0,6 мВ (n = 150). То есть значение Н-эффекта снижалось, составляя в описанных условиях эксперимента 4,4 ± 0,5 мВ (n = 150) (р < 0,05).
Изучение влияния АТФ на сократительную функцию диафрагмы несенсибилизированной мыши показало, что карбахолин в концентрации 2x10-4 М вызывает сократительный ответ полоски мышцы силой 335,2 ± 93,47 мг. Инкубация с АТФ увеличивает этот показатель до 425,2 ± 100,9 мг (р < 0,01).
Изучение неквантовой секреции Ах в диафрагме показало, что МП покоя, изначально составляющий (-70,7) ± 1,9 мВ (n = 150), в присутствии ТБК возрастал до (–75,9) ± 0,7 мВ (n = 150). Таким образом, Н-эффект в контроле составлял 5,2 ± 0,4 мВ (n = 150).
После инкубации с АТФ МП покоя, изначально составляющий (–70,0) ± 0,4 мВ (n = 150), в присутствии ТБК возрастал до (–71,5) ± 0,5 мВ (n = 150). То есть значение Н-эффекта снижалось, составляя в описанных условиях эксперимента 1,5 ± 0,5 мВ (n = 150) (р < 0,001).
Изучение влияния АТФ на сократительную функцию диафрагмы сенсибилизированной мыши показало, что карбахолин в концентрации 2x10-4 М вызывает сократительный ответ полоски мышцы силой 469,83 ± 86,78 мг. Инкубация с АТФ увеличивает этот показатель до 540,67 ± 80,34 мг (р < 0,01).
Изучение неквантовой секреции Ах в диафрагме сенсибилизированной мыши показало, что МП покоя, изначально составляющий (–70,0) ± 1,5 мВ (n = 150), в присутствии ТБК возрастал до (–74,4) ± 0,6 мВ (n = 150). Таким образом, Н-эффект в контроле составлял 4,4± 0,5 мВ (n = 150). После инкубации с АТФ МП покоя, изначально составляющий (-69,1) ± 0,4 мВ (n = 150), в присутствии ТБК возрастал до (–71,5) ± 0,6 мВ (n = 150). То есть значение Н-эффекта снижалось, составляя в описанных условиях эксперимента 2,4 ± 0,6 мВ (n = 150) (р < 0,001).
Предварительная перфузия сурамином (100 мкМ) устраняла влияние АТФ на Кх-вызванное сокращение диафрагмальной мышцы и неквантовую секрецию Ах как интактных, так и сенсибилизированных мышей. Аденозин в той же концентрации, что и АТФ не влиял на неквантовую секрецию Ах и не изменял сократительных свойств полоски диафрагмы как интактных, так и сенсибилизированных мышей.
ОБСУЖДЕНИЕ
Наши исследования показали, что сенсибилизация мыши к яичному альбумину приводит к увеличению силы сокращения полоски диафрагмальной мышцы мыши и снижает уровень неквантовой секреции Ах в зоне концевой пластинки. Увеличение силы сокращения на Кх до 140,16 % является следствием увеличения чувствительности постсинапса к холиномиметику (ослабление механизмов десенситизации холинорецепторов постсинаптической мембраны), отражением чего является снижение Н-эффекта до 84,61 % (рис. 1).
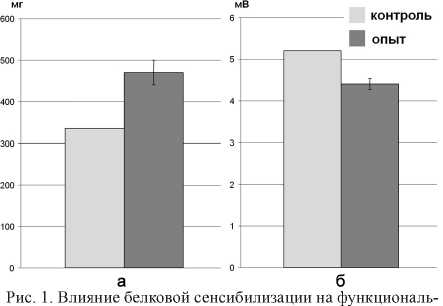
ные характеристики изолированной полоски диафрагмы мыши: а — силу сокращения, вызванного Кх (2 x1g-4 М);
б — величину Н-эффекта
Обнаруженная динамика свидетельствует, что белковая сенсибилизация затрагивает, в первую очередь, холиноопосредованные процессы возбуждения МВ. Дальнейшие наши исследования были посвящены поиску возможных причин, определяющих вариабельность функциональных свойств мышцы при белковой сенсибилизации. Изменения функциональных свойств холиновозбудимой мембраны, лежащие в их основе, могут являться следствием изменений, возникающих в механизмах выделения кофакторов синаптической передачи. Исходя из данных Tsai T. L. et al. [17], показавших участие АТФ в генерации иммунного ответа, мы предположили возможное участие в процессах изменения функции поперечнополосатых мышц при белковой сенсибилизации пуринов. Это определило изучение нами, в рамках выбранных экспериментальных моделей, динамики функциональных свойств полоски диафрагмы мыши после инкубации мышцы с АТФ.
Вопрос влияния АТФ (в том числе и экзогенной) на СМ имеет самостоятельную ценность, поскольку механизмы регуляции их функций с участием пуринов, в отличие от гладкой и сердечной мышц, до настоящего времени исследованы очень слабо.
Нами показано, что экзогенная АТФ повышает силу сокращения полоски диафрагмы мыши на КХ и снижает уровень неквантовой секреции Ах. Вектор динамики силы и величины Н-эффекта совпадал с таковыми при БС, что позволило нам предположить аналогичную последовательность изменения. Увеличение силы сокращения мышцы на Кх после перфузии с АТФ является следствием возрастания чувствительности постсинапса к холиномиметику. Угнетение неквантовой секреции Ах в зоне синапса вызывает снижение механизмов десенситизации ХР постсинаптической мембраны, что приводит к возрастанию ее чувствительности к Кх (рис. 2).
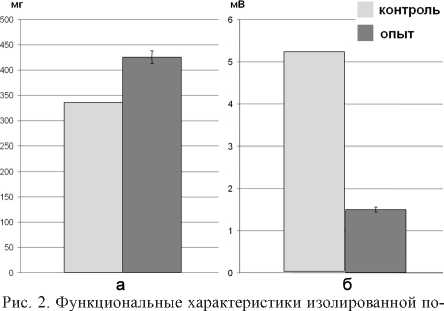
лоски диафрагмы интактной мыши до (контроль) и после (опыт) влияния АТФ: а — сила сокращения, вызванного Кх
(2x10 4 М) (п = 10); б — величина Н-эффекта (n = 150)
Изучение влияния АТФ на динамику вышеперечисленных функциональных свойств диафрагмы сенсибилизированных мышей демонстрирует ту же направленность, что указывает на отсутствие принципиальных различий в механизмах влияния пуринов на мышцы сенсибилизированных и интактных мышей (рис. 3).
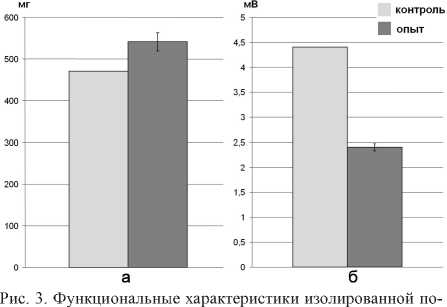
лоски диафрагмы сенсибилизированной мыши до (контроль) и после (опыт) влияния АТФ: а — сила сокращения, вызванного Кх (2x10^ М) (п = 7); б — величина Н-эффекта (n = 150)
Однако количественная оценка этих изменений выявляет определенные различия, указывающие на АТФ как на одного из участников изменения холиноопосредованных механизмов возбуждения в работе дыхательных мышц при белковой сенсибилизации. И у сенсибилизированных, и у интактных животных экзогенная АТФ увеличивала силу сокращения на Кх и уменьшала величину Н-эффекта. Однако, если сила сокращения мышцы у интактных животных возрастала на 26 %, то у сенсибилизированных — лишь на 15 %. Н-эффект диафрагмы несенсибилизированных мышей после влияния АТФ снижался до 28,8 % от исходного, у сенсибилизированных же лишь до 54,5 0%. Менее выра- женная динамика функциональных свойств диафрагмы, вызванная АТФ у сенсибилизированных мышей в сравнении с контролем, позволяет нам предполагать ее участие в механизмах изменений функциональных свойств дыхательных мышц, вызванных белковой сенсибилизацией.
Реализация постсинаптических эффектов на диафрагме мыши аналогична таковым у большинства других скелетных мышц и осуществляется через P2-рецепторы. Подтверждением этому служат как литературные данные, так и результаты собственных исследований. Во-первых, сурамин, антагонист P2-рецепторов, устранял влияние ATФ во всех экспериментальных моделях. Кроме того, замена в наших экспериментах АТФ на аденозин, реализующий свое действие не через Р2, а через аденозиновые Р1-рецепторы [8], не вызывала изменения ни параметров КХ-вызванного сокращения диафрагмы, ни величины Н-эффекта.
К сожалению, в литературе нам не встретилось работ об участии пуринов в механизмах изменения функции поперечнополосатых мышц при аллергии. Однако результаты наших экспериментов свидетельствуют, что внеклеточная АТФ может являться участником изменения свойств диафрагмальной мышцы мыши при сенсибилизации животного к чужеродному белку. Механизмы эти весьма разнообразны и включают в себя как прямое действие пуринов на контрактильную функцию дыхательных мышц, так и их влияние на не- которые этапы генерации иммунного ответа. В первом случае — это влияние АТФ на секрецию медиатора и системы внутриклеточных посредников поперечнополосатых мышц [6, 9, 10]. Также мы можем предположить ее влияние на работу АТФ-зависимых калиевых каналов [17]. Во втором случае, в литературе показана способность АТФ, участвуя в увеличении продукции интерлейкина-1, усиливать специфическое звено иммунитета [14]. Внеклеточная АТФ при генерации иммунного ответа помогает образованию активной каспазы-1, что, в свою очередь, обеспечивает секрецию биологически активных форм интерлейкина-1. Гиперэкспрессия рецептора Р2Х7 приводит к секреции зрелого интерлейкина-1β [15].
Являясь экспериментальной моделью аллергии, белковая сенсибилизация позволяет изучать механизмы как патологических, так и компенсаторных изменений, возникающих в системе внешнего дыхания. В нашей работе было показано, что сенсибилизация животных приводит к увеличению силы сокращения полоски диафрагмы мыши на Кх. Этот факт, а также меньшая изменчивость функциональных свойств диафрагмы, вызванная АТФ у сенсибилизированных мышей в сравнении с контролем, является возможным свидетельством развития механизмов резистентности, устойчивости к внешним нагрузкам у дыхательных мышц, которая должна появляться у них при бронхоспастическом синдроме и бронхиальной астме.
ВЫВОДЫ
-
1. Увеличение силы сокращения на карбахолин диафрагмальной мышцы мыши in vitro при белко-вой сенсибилизации является следствием повышения чувствительности постсинаптической мембраны мышечных волокон к холиномиметику, что обусловлено снижением уровня неквантовой секреции ацетилхолина в зоне концевой пластинки.
-
2. Увеличение силы сокращения полоски диафрагмы мыши на карбахолин, вызванное экзоген-
- ной АТФ, обусловлено снижением уровня неквантовой секреции ацетилхолина в зоне концевой пластинки.
-
3. Белковая сенсибилизация снижает динамику функциональных свойств диафрагмы мыши, вызванную экзогенной АТФ, угнетая увеличение силы сокращения мышцы на карбахолин вследствие адекватного снижения неквантовой секреции ацетилхолина в зоне концевой пластинки.
