Возникновение диплоидных растений в диплоидно-тетраплоидных скрещиваниях у кукурузы (Zea mays L.): анализ плоидности эндосперма выполненных зерновок
: анализ плоидности эндосперма выполненных зерновок")
Автор: Мавлютова Л.И., Эльконин Л.А., Колесова А.Ю.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Генетика, цитогенетика
Статья в выпуске: 5 т.60, 2025 года.
Бесплатный доступ
Важным компонентом апомиктичного размножения у растений служит способность к формированию нередуцированных зародышевых мешков (ЗМ). Гетероплоидные скрещивания - эффективный инструмент для выявления таких ЗМ, поскольку при оплодотворении диплоидных ЗМ пыльцой тетраплоидов в центральной клетке ЗМ происходит слияние двух диплоидных полярных ядер с диплоидным спермием, ведущее к восстановлению геномного баланса в эндосперме (2♀:1♂) и развитию полноценных зерновок. Ранее мы обнаружили, что на початках диплоидной линии кукурузы АТ, способной к гаплоидному партеногенезу, и ее гибридов при опылении пыльцой тетраплоидов формировались выполненные зерновки, из которых развивались диплоидные растения матроклинного типа и тетраплоидные гибриды. Нашей целью стала дальнейшая проверка гипотезы о возникновении выполненных зерновок в гетероплоидных скрещиваниях 2 n × 4 n на основе нередуцированных зародышевых мешков, образующихся у материнских линий, с помощью анализа плоидности эндосперма таких зерновок методом проточной цитометрии. В работе использовали выполненные зерновки, развившиеся на початках диплоидных линий кукурузы: АТ ГПЛ (2 n), обладающей способностью к гаплоидному партеногенезу, 18-10-4 (2 n), В47 (2 n), Коричневый Маркер (КМ) (2 n), Краснодарская 2 (2 n), Краснодарская 9 (2 n), Краснодарская 22 (2 n), а также гибридов F1 и F3(В47/АT ГПЛ) (2 n), F1(КМ1/Сахарная1) (2 n) и F1(КМ/АТ ГПЛ) (2 n), которые были опылены пыльцой тетраплоидных линий Тетра-Парий (ТП) (4 n), Краснодарский тетраплоид (КрТ) (4 n) и Черная Тетра (ЧТ) (4 n). Скрещивания были проведены в 2022 году. Растения выращивали на опытном участке Селекционного комплекса ФАНЦ Юго-Востока (г. Саратов). Пыльцу с метелки каждого тетраплоидного растения стряхивали в индивидуальный чистый изолятор и наносили на рыльца початка диплоидного материнского растения. На 18-е сут после опыления с початков диплоидных линий собирали крупные выполненные зерновки, стерилизовали, отделяли эндосперм и фиксировали его в смеси спирта (95 %) и пропионовой кислоты (3:1) в течение 3 ч, отмывали в 70 % спирте и оставляли в нем на хранение при температуре -20 °C для последующего исследования плоидности с помощью проточной цитометрии. Зародыш помещали в пробирки на питательную агаризованную среду MS и выращивали в растильной комнате (фотопериод 16 ч день/8 ч ночь; 26±1 °C). Проростки с развитыми корнями переносили на несколько суток в пробирки с водой для адаптации к нестерильным условиям, после чего высаживали в теплицу в пластиковые сосуды и затем - на делянку экспериментального поля Селекционного центра ФАНЦ Юго-Востока. Для определения плоидности использовали кончики корешков растений, выращенных в пластиковых сосудах, на 12-е-14-е сут после высаживания в землю. Для каждого растения плоидность определяли на основании анализа 8-10 метафазных пластинок. При исследовании плоидности с помощью проточной цитометрии в качестве стандартов использовали ядра из листа гаплоидного растения линии АТ ГПЛ и из эндоспермов зерновок АТ ГПЛ и тетраплоидной линии Черная Тетра. У разных линий и гибридов частота початков с выполненными зерновками варьировала от 7 до 75 %; частота выполненных зерновок от общего числа семян варьировала от 0,7 до 85,4 % и зависела от генотипа опылителя: при использовании ТП и ЧТ частота была значимо выше, чем для КрТ; частота початков с выполненными зерновками была наибольшей у ТП. Из выполненных зерновок, как показал цитологический анализ корневых меристем проростков, развивались диплоидные, триплоидные и тетраплоидные растения. В эндосперме выполненных зерновок было обнаружено несколько пиков флюоресценции, при этом 1-й пик наблюдался при значениях, в 2 раза превышающих таковые в эндосперме диплоидной линии (3С) и соответствовал 6С, аналогично показателю 1-го пика флюоресценции эндосперма тетраплоидной линии. Такой результат наблюдался у эндосперма зерновок, из которых были получены как тетраплоидные гибриды, так и диплоидные «матроклинные» растения. Кроме того, как в триплоидном, так и в гексаплоидном эндосперме наблюдались пики, соответствовавшие более высоким значениям плоидности (12С, 24С), которые были обусловлены процессами эндополиплоидизации. Полученные данные свидетельствуют в пользу гипотезы о развитии выполненных зерновок в скрещиваниях 2 n × 4 n у кукурузы на основе нередуцированных ЗМ, важного компонента апомиксиса, и демонстрируют эффективность использования тетраплоидных опылителей для выявления таких зародышевых мешков у линий и гибридов кукурузы, в том числе в работах по созданию апомиктичных генотипов с помощью геномного редактирования. Кроме того, полученные результаты впервые подкрепляют балансовую теорию развития эндосперма прямыми экспериментальными доказательствами, поскольку плоидность эндосперма зерновок, формирующихся в гетероплоидных скрещиваниях, с помощью проточной цитометрии ранее не исследовалась.
Апомиксис, гетероплоидные скрещивания, геномный баланс, проточная цитометрия
Короткий адрес: https://sciup.org/142247701
IDR: 142247701 | УДК: 633.15:581.163 | DOI: 10.15389/agrobiology.2025.5.827rus
Origin of diploid plants in diploid–tetraploid crosses in maize (Zea mays L.): analysis of endosperm ploidy of plump kernels
An important component of apomictic reproduction in plants is the ability to form unreduced embryo sacs (ES). Heteroploid crosses are an effective tool for identifying such ESs, since when diploid ES is fertilized with tetraploid pollen, two diploid polar nuclei fuse with a diploid sperm, leading to the restoration of the genomic balance in the endosperm (2♀:1♂) and formation of normally developed seeds. Previously, we found that on the ears of the diploid AT maize line, capable of haploid parthenogenesis, as well as on the ears of its hybrids, when pollinated with pollen from tetraploids, plump kernels were formed, from which diploid maternal-type plants and tetraploid hybrids developed. Our aim was to further test the hypothesis on the formation of plump kernels in 2 n × 4 n heteroploid crosses from unreduced embryo sacs developed in maternal lines by analyzing the endosperm ploidy of such kernels using flow cytometry. In this study, we used the plump kernels developed on the ears of diploid maize lines: AT GPL (2 n), which has the ability for haploid parthenogenesis, 18-10-4 (2 n), V47 (2 n), Korichnevyi Marker (KM) (2 n), Krasnodarskaya 2 (2 n), Krasnodarskaya 9 (2 n), Krasnodarskaya 22 (2 n), as well as the F1 and F3 hybrids V47/AT GPL (2 n), F1 KM1/Sakharnaya1 (2 n), and F1 KM/AT GPL (2 n) which were pollinated with pollen of the tetraploid lines Tetra-Pariy (TP) (4 n), Krasnodarskiy tetraploid (KrT) (4 n), and Chernaya Tetra (ChT) (4 n). The crosses were carried out in 2022. The plants were grown in the experimental field of the Federal Agricultural Research Centre of the South-East Region (Saratov, Russia). Pollen from the panicle of each tetraploid plant was shaken off into an individual clean parchment bag and applied to the stigmas of the ears of the diploid maternal plant. On the 18th day after pollination, large, mature kernels were collected from the ears of the diploid lines, sterilized, the endosperm was separated and fixed in a mixture of alcohol (95 %) and propionic acid (3:1) for 3 h, washed in 70 % alcohol and stored in 70 % alcohol at a temperature of -20 °C for subsequent ploidy analysis using flow cytometry. Embryos were placed in test tubes on MS agar nutrient medium, and grown in a growth room (photoperiod 16 h day/8 h night; 26±1 °C). Seedlings with developed roots were transferred to test tubes with water for several days to acclimate to non-sterile conditions, after which they were planted in plastic containers in a greenhouse and then in an experimental field of Federal Research Centre of Agriculture of South-East Region. Root tips from plants grown in plastic containers were used to determine ploidy on the 12th to 14th days after planting in the ground. For each plant, ploidy was determined based on the analysis of 8-10 metaphase plates. When studying ploidy using flow cytometry, nuclei from the leaf of the haploid plant of AT GPL line, and from the endosperms of AT GPL and tetraploid ChT kernels were used as standards. The frequency of ears with normally developed plump kernels varied from 7 % to 75 % among different lines and hybrids; the frequency of plump kernels from the total number of seeds varied from 0.7 to 85.4 % and depended on the pollinator genotype: with TP and ChT, the frequency was significantly higher than for KrT; the frequency of ears with plump kernels was the highest for TP. Cytological analysis of the root meristems of seedlings showed that diploid, triploid, and tetraploid plants developed from the plump kernels. Several fluorescence peaks were detected in the endosperm of the plump kernels, with the first peak observed at values twice higher than those in the endosperm of the diploid line (3C) and corresponding to 6C, similar to the value of the first fluorescence peak of the endosperm of the tetraploid line. This result was observed in the endosperm of kernels from which both tetraploid hybrids and diploid "maternal-type" plants were obtained. Furthermore, peaks corresponding to higher ploidy values (12C, 24C) were also observed in both triploid and hexaploid endosperm, which were attributed to endopolyploidization processes. These data support the hypothesis that normally developed plump kernels develop in 2 n × 4 n maize crosses on the bases on unreduced ES, an important component of apomixis, and demonstrate the effectiveness of tetraploid pollinators to identify such embryo sacs in maize lines and hybrids, including in studies on the creation of apomictic genotypes using genome editing. Furthermore, the obtained results provide the first direct experimental evidence to support the balance theory of endosperm development, since the ploidy of endosperm of kernels formed in heteroploid crosses has not previously been studied using flow cytometry.
Текст научной статьи Возникновение диплоидных растений в диплоидно-тетраплоидных скрещиваниях у кукурузы (Zea mays L.): анализ плоидности эндосперма выполненных зерновок
Апомиксис — способ семенного размножения покрытосеменных растений, при котором зародыш развивается из неоплодотворенной яйцеклетки, приковывает внимание генетиков и селекционеров растений уже более 100 лет. Интенсивные исследования апомиктичного способа размножения связаны с надеждами на возможность его использования для закрепления гетерозиса гибридов F 1 и их клонального размножения семенами (13). Апомиксис достаточно широко распространен в дикой флоре, преимущественно у полиплоидных видов, и редко встречается у культурных растений. В этой связи высказывалось мнение о принципиальной невозможности создания линий и гибридов диплоидных культурных растений с наследуемой способностью к апомиктичному размножению (4). Однако недавние публикации результатов экспериментов исследовательских групп из Франции и Китая, в которых сообщалось о создании гибридов F 1 риса, способных к апомиктичному размножению, на основе использования технологии геномного редактирования, свидетельствуют о достижимости решения этой задачи (5-8).
Один из ключевых факторов формирования апомиктичных семян — способность к образованию нередуцированных зародышевых мешков (ЗМ), диплоспорических, возникающих в результате нарушений мейоза, либо апоспорических, развивающихся из соматических клеток, расположенных вблизи мейотического ЗМ (1, 2). Эффективным подходом для выявления способности к формированию нередуцированных ЗМ у диплоидных растений, в частности у кукурузы, может служить использование тетраплоидных опылителей. Известно, что у кукурузы скрещивания $ 2 n х ^ 4 n чаще всего приводят к формированию щуплых зерновок вследствие нарушения баланса материнского (м) и отцовского (о) геномов 2м:1о в эндосперме, который в таких скрещиваниях имеет геномный баланс 2м:2о из-за слияния двух гаплоидных полярных ядер с диплоидным спермием (9-12). Однако в экспериментах, проведенных нами ранее, при опылении диплоидной линии кукурузы АТ, способной к гаплоидному партеногенезу (13), а также ее гибридов, пыльцой разных тетраплоидных линий, в некоторых случаях на початках формировались крупные выполненные зерновки, из которых развивались диплоидные матроклинные растения и тетраплоидные гибриды (14). На основании этих фактов было высказано предположение, что выполненные зерновки могли возникать вследствие оплодотворения нередуцированных ЗМ диплоидными спермиями, поскольку в таких случаях эндосперм развивается в результате слияния двух диплоидных полярных ядер с диплоидным спермием, в результате чего образуется гексаплоидный эндосперм и восстанавливается баланс материнского и отцовского геномов 2м:1о (14, 15).
Для проверки этой гипотезы мы провели генотипирование диплоидных «матроклинных» растений, развивающихся из выполненных зерновок, 828
с помощью SSR- и InDel-маркеров. Полученные результаты свидетельствовали в пользу гипотезы о возникновении таких растений на основе оплодотворения нередуцированных яйцеклеток диплоидными спермиями и последующей элиминации хромосом одного из родительских компонентов (преимущественно тетраплоидного опылителя) (16). Некоторые растения по совокупности фенотипических признаков, практически, полностью соответствовали материнской линии (АТ ГПЛ), однако на одной-двух хромосомах присутствовали маркеры отцовской линии.
Нами впервые с помощью проточной цитометрии получены экспериментальные доказательства, свидетельствующие, что развитие выполненных зерновок у кукурузы в скрещиваниях 2 n х 4 n происходит на основе оплодотворения нередуцированных зародышевых мешков, формирующихся у материнских линий. Такая гексаплоидная природа эндосперма у выполненных зерновок, развивающихся в скрещиваниях 2 n х 4 n , подкрепляет балансовую теорию развития эндосперма прямыми экспериментальными доказательствами.
Нашей целью стала дальнейшая проверка гипотезы о возникновении выполненных зерновок в гетероплоидных скрещиваниях 2 n * 4 n на основе нередуцированных зародышевых мешков, образующихся у материнских линий, посредством анализа плоидности эндосперма таких зерновок с помощью проточной цитометрии.
Методика. В работе использовали выполненные зерновки, развившиеся на початках диплоидных линий кукурузы: АТ ГПЛ (2 n ), обладающей способностью к гаплоидному партеногенезу, 18-10-4 (2 n ), В47 (2 n ), КМ (2 n ), Краснодарская 2 (2 n ), Краснодарская 9 (2 n ), Краснодарская 22 (2 n ), а также гибридов F 1 и F 3 (B47/AT ГПЛ) (2 n ), F 1 (КМ1/Сахарная1) (2 n ) и F 1 (КМ/АТ ГПЛ) (2 n ), которые были опылены пыльцой тетраплоидных линий Тетра-Парий (ТП) (4 n ), Краснодарский тетраплоид (КрТ) (4 n ) и Черная Тетра (ЧТ) (4 n ). Скрещивания были проведены в 2022 году. Растения выращивали на опытном участке Селекционного комплекса ФАНЦ Юго-Востока (г. Саратов), пространственно изолированном от других посевов кукурузы с целью исключения случайного опыления. У всех растений для предотвращения неконтролируемого опыления до начала цветения удаляли метелки и изолировали початки пергаментными изоляторами. При проведении скрещиваний пыльцу с метелки каждого тетраплоидного растения стряхивали в индивидуальный чистый изолятор и наносили на рыльца початка диплоидного материнского растения.
На 18-е сут после опыления с початков диплоидных линий собирали крупные выполненные зерновки, которые примерно в 2 раза и более превосходили по объему другие зерновки, завязавшиеся на том же початке. Зерновки стерилизовали 70 % этиловым спиртом (30 с) и 0,1 % раствором сулемы (15 мин). После 3-кратной промывки автоклавированной водой в стерильных условиях отделяли эндосперм и фиксировали его в смеси спирта (95 %) и пропионовой кислоты (3:1) в течение 3 ч, отмывали 3кратно в 70 % спирте и оставляли в нем на хранение при температуре - 20 ° C для последующего исследования плоидности с помощью проточной цитометрии, а зародыш помещали в пробирки на питательную агаризованную среду MS.
Зародыши выращивали в растильной комнате, оборудованной лампами Fluora L 36w/77 («Osram Licht AG», Германия) (фотопериод 16 ч день/8 ч ночь; 26±1 °C). Проростки с развитыми корнями переносили на несколько суток в пробирки с водой для адаптации к нестерильным условиям, после чего высаживали в теплицу в пластиковые сосуды и затем — на делянку экспериментального поля Селекционного центра ФАНЦ Юго-Востока. Агроклиматические условия соответствовали росту и развитию растений кукурузы в полевых условиях и способствовали получению семян.
Для определения плоидности кончики корешков растений, выращенных в пластиковых сосудах, фиксировали в ацетоалкоголе (1:3) (14). Фиксацию проводили на 12-е-14-е сут после высаживания в землю в 10-11 ч утра при 26-28 ° С. Перед фиксацией кончики корешков помещали на 3 ч в насыщенный раствор а -бром-нафталина для укорачивания хромосом. Кончики корешков проводили через серию растворов в следующей последовательности: 3,6 % (1N) HCl (10 мин), 50 % HCl (20 мин), дистиллированная вода (1-2 мин), 45 % уксусная кислота (20 мин). Затем окрашивали в 4 % ацетогематоксилине (30-60 мин). Препараты анализировали с помощью светового микроскопа AxioScope A1 («Zeiss», Германия) и фотографировали с помощью цифровой камеры MrC («Zeiss», Германия) при увеличении 1000½. Для каждого растения плоидность определяли на основании анализа 8-10 метафазных пластинок.
Для проведения проточной цитометрии эндосперм измельчали лезвием в течение 1,5-2 мин в пластиковой чашке Петри на льду в 2 мл охлажденного буфера Голбрайта (45 мМ MgCl 2 , 30 мM цитрата натрия, 20 мM МОРS, 1 мг/мл Triton X-100, pH 7,0) (17), фильтровали через фильтр с диаметром пор 50 мкм и центрифугировали (5418 R, «Eppendorf», Германия) в течение 2 мин при 100 g и 4 ° С. Супернатант сливали, а осадок с ядрами ресуспендировали в 1 мл буфера с добавлением DAPI (4 мкг/мл) и РНКазы (50 мкг/мл). Окрашивали 10 мин на льду в темноте, встряхивали и центрифугировали на вортексе (2 с) (Microspin FV-2400, «Biosan», Латвия) для осаждения крахмала, после чего надосадочную жидкость переносили в другую пробирку и анализировали на флуороцитометре BD FAC SCANTO II («BD Bioscience», Германия) в Институте биохимии и генетики УФИЦ РАН (г. Уфа) при FSC-300, SSC-280, Pacific Blue-A ( X = 250 нм). В качестве стандартов использовали ядра из листа гаплоидного растения линии АТ ГПЛ и из эндоспермов зерновок АТ ГПЛ и тетраплоидной линии Черная Тетра. В этих экспериментах исследовали выполненные зерновки из следующих комбинаций скрещиваний: F 3 (B47/AT ГПЛ)(2 n ) х ТП(4 n ), F 1 (КМ/Сахар-ная1)(2 n ) х КрТ(4 n ), АТ ГПЛ(2 n ) х КрТ(4 n ), В47(2 n ) х ТП(4 n ). В каждом образце учитывали по 2,5-3 тыс. ядер. Анализировали только экспериментальные данные, в которых величина ошибки каждого пика не превышала 5 %.
Статистическую обработку данных проводили посредством сравнения долей по методу Фишера, применяющемуся для малых, а также неравновеликих выборок (18). Обработку результатов проточной цитофотометрии осуществляли при помощи программного обеспечения FSC-Express, v.7.
Результаты. Для получения выполненных зерновок с целью последующего анализа плоидности их эндосперма были проведены опыления ряда диплоидных линий и гибридов пыльцой тетраплоидных линий Тетра-Парий, КрТ и Черная Тетра. В этих скрещиваниях в большинстве случаев формировались щуплые зерновки с неразвитым эндоспермом, что следовало ожидать, учитывая тетраплоидную природу эндосперма таких зерновок, геномный баланс которых соответствовал соотношению 2м:2о. Однако, наряду с зерновками со щуплым эндоспермом, на некоторых початках изредка развивались зерновки с полностью или частично выполненным эндоспермом (рис. 1).

Рис. 1. Щуплые (белые стрелки) и выполненные (черные стрелки) зерновки кукурузы ( Zea mays L.), завязавшиеся на початках диплоидных (2 n ) растений F 1 (В47/АТ ГПЛ) (a) и F 3 (В47/АТ ГПЛ) (б, в) , опыленных пыльцой тетраплоидных (4 n ) линий Краснодарский Тетраплоид (а) и Черная Тетра (б, в) (г. Саратов, 2022 год) .
1. Частота выполненных зерновок, завязавшихся на початке диплоидных линий и гибридов кукурузы (Zea mays L.) при их опылении пыльцой от тетраплоид-ных линий (г. Саратов, 2022 год)
|
Линия, гибрид (2 n ) |
Опылитель (4 n ) |
Число опыленных початков, шт. |
Число зерновок на початках с выполненными зерновками, шт. |
Частота выполненных зерновок, % |
||||
|
всего |
с выполнен ными зерновками, шт. (%) |
всего |
уплых |
частично выполненных |
выпол ненных |
|||
|
АТ ГПЛ |
ЧТ |
9 |
3 (33,3) |
238 |
231 |
4 |
3 |
1,2 (0,7-3,0)1 |
|
КрТ |
21 |
2 (9,5) |
120 |
11 |
5 |
4 |
3,3 (2,7-9,0) |
|
|
КМ |
КрТ |
17 |
2 (11,7) |
91 |
37 |
52 |
2 |
2,1 (1,6-1,9) |
|
В47 |
ТП |
4 |
1 (25,0) |
34 |
33 |
2 |
2 |
5,8 (5,8) |
|
Краснодарская 2 |
КрТ |
18 |
3 (16,6) |
312 |
307 |
0 |
5 |
1,6 (0,5-100) |
|
Краснодарская 9 |
КрТ |
16 |
4 (25,0) |
306 |
275 |
20 |
11 |
4,1 (1,5-7,5) |
|
Краснодарская 22 |
ЧТ |
9 |
2 (22,2) |
441 |
414 |
19 |
8 |
1,8 (1,7-1,8) |
|
КрТ |
10 |
2 (20,0) |
10 |
4 |
4 |
2 |
20,0 (20-100) |
|
|
18-10-4 |
ТП |
4 |
3 (75,0) |
235 |
150 |
73 |
12 |
5,1 (1,6-8,3) |
|
ЧТ |
6 |
1 (16,6) |
43 |
40 |
2 |
1 |
2,3 (2,3) |
|
|
F 1 (КМ/АТ ГПЛ) |
ТП |
6 |
2 (33,3) |
167 |
131 |
22 |
14 |
8,3 (5,8-11,1) |
|
ЧТ |
7 |
1 (14,2) |
37 |
33 |
3 |
1 |
2,7 (2,7) |
|
|
КрТ |
26 |
1 (3,8) |
308 |
300 |
5 |
3 |
0,9 (0,9) |
|
|
F 1 (В47/АТ ГПЛ) |
ЧТ |
4 |
2 (50,0) |
126 |
117 |
4 |
5 |
4,0 (1,8-5,5) |
|
КрТ |
3 |
2 (66,6) |
68 |
61 |
3 |
4 |
5,9 (1,8-2,3) |
|
|
F 3 (В47/АТ ГПЛ) |
КрТ |
41 |
3 (7,3) |
980 |
929 |
44 |
7 |
0,7 (0,2-2,2) |
|
ЧТ |
20 |
3 (15,0) |
377 |
363 |
8 |
6 |
1,5 (0,5-12,9) |
|
|
ТП |
7 |
3 (42,8) |
378 |
264 |
102 |
12 |
3,2 (0,8-8,5) |
|
|
F 1 (КМ1/Сахарная 1) |
ЧТ |
7 |
3 (42,8) |
48 |
7 |
0 |
41 |
85,4 (33,3-100,0) |
|
КрТ |
14 |
8 (57,1) |
436 |
329 |
90 |
17 |
3,9 (1, 9-37,5) |
|
|
Всего |
КрТ |
166 |
27 (16,3) |
2631 |
2353 |
223 |
55 |
2,1 |
|
ЧТ |
62 |
15 (24,2) |
1309 |
1204 |
40 |
65 |
5,0*** |
|
|
ТП |
17 |
8 (47,1)*, ** |
780 |
545 |
197 |
38 |
4,9*** |
|
|
Примечание. |
1 — в скобках указано варьирование на отдельных початках. КМ — |
Коричневый Маркер, |
||||||
ЧТ — Черная Тетра, КрТ — Краснодарский Тетраплоид, ТП — Тетра Парий. Линия 18-10-4 была создана на основе диплоидного растения, полученного из выполненной зерновки, завязавшейся в скрещивании КМ ½ ЧТ.
* и ** p < 0,05 и p < 0,01 соответственно по сравнению с ЧТ и КрТ.
*** p < 0,001 по сравнению с КрТ в соответствии с F -критерием.
У разных линий и гибридов частота початков с выполненными зерновками варьировала от 7 до 75 % (табл. 1); частота выполненных зерновок от общего числа семян — от 0,7 до 85,4 % и зависела от генотипа опылителя. При использовании линий ТП и ЧТ частота была значительно выше, чем у КрТ; частота початков с выполненными зерновками была наибольшей у ТП.
Из выполненных зерновок развивались диплоидные, триплоидные и тетрап-лоидные растения (табл. 2, рис. 2).
2. Плоидность растений, полученных из выполненных зерновок кукурузы ( Zea mays L.), завязавшихся в гетероплоидных скрещиваниях (2 n ½ 4 n ) (г. Саратов, 2022 год)
|
Линия, гибрид (2 n ) |
Опылитель (4 n ) |
Число растений с уровнем плоидности |
||
|
2 n |
3 n |
4 n |
||
|
АТ ГПЛ |
КрТ |
1 |
1 |
1 |
|
ЧТ |
1 |
– |
1 |
|
|
В47 |
ТП |
– |
– |
2 |
|
КМ |
КрТ |
– |
– |
1 |
|
F 1 (КМ/АТ ГПЛ) |
ТП |
1 |
– |
– |
|
ЧТ |
1 |
4 |
– |
|
|
F 1 (В47/АТ ГПЛ) |
КрТ |
1 |
– |
– |
|
F 3 (В47/АТ ГПЛ) |
КрТ |
1 |
2 |
– |
|
ТП |
4 |
– |
– |
|
|
F 1 (КМ1/Сахарная 1) |
ЧТ |
2 |
– |
– |
|
КрТ |
– |
2 |
– |
|
Примечание. КМ — Коричневый Маркер, КрТ — Краснодарский Тетраплоид, ЧТ — Черная Тетра, ТП — Тетра Парий. Прочерки означают отсутствие данных
Для анализа плоидности эндосперма были отобраны выполненные зерновки из комбинаций F 3 (В47/АТ ГПЛ) ½ ТП и F 1 (КМ/Сахарное1) ½ КрТ, из которых развивались диплоиды, и АТ ГПЛ ½ КрТ и В47 ½ ТП, из которых развивались тетраплоиды (см. табл. 2).
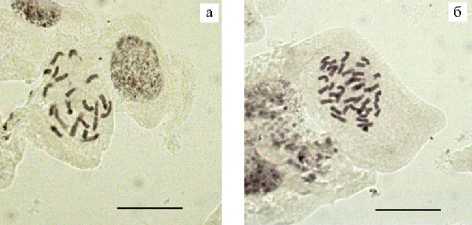
Рис. 2. Метафазные пластинки диплоидного (a) и тетраплоидного (б) растений кукурузы ( Zea mays L.), выросших из выполненных зерновок, которые завязались на початках при скрещиваниях 18-10-4 (2 n ) ½ Тетра-Парий (4 n ) и В47 (2 n ) ½ Тетра-Парий (4 n ) (окрашивание ацетогематоксилином, шкала 20 мкм, 1000½, микроскоп AxioScope A1, «Zeiss», Германия) .
Согласно результатам исследования плоидности эндоспермов с помощью проточной цитофотометрии (табл. 3, рис. 3), во всех образцах наблюдалось несколько пиков флюоресценции, что следовало ожидать, учитывая известное явление эндополиплоидизации ядер эндосперма у кукурузы (9, 19, 20). У эндосперма диплоидного растения (3 n -стандарт), плоидность ядер которого должна быть не менее 3 n , среднее значение интенсивности флюоресценции 1-го пика соответствовало 95 ед. шкалы на оси Х, тогда как среднее значение для 2-го пика было 191 ед. (см. рис. 3, Б). Очевидно, что эти пики соответствовали содержанию ДНК 3С и 6С, поскольку пики 1С и 2С у ядер, выделенных из листа гаплоида ( n -стандарт), имели значения 35 и 70 ед. (см. табл. 3; рис. 3, А). Кроме того, в триплоидном эндосперме наблюдались пики, соответствовавшие более высоким значениям плоидно-сти (12С, 24С), которые, как и ожидалось, присутствовали вследствие эн-дополиплоидизации. У эндосперма тетраплоидной линии (6 n -стандарт) среднее значение 1-го пика флюоресценции было почти в 2 раза выше, чем у триплоидного эндосперма (182 ед.) и, по-видимому, соответствовало величине 6С (см. табл. 3). Кроме того, в гексаплоидном эндосперме наблюдались пики, соответствовавшие более высоким уровням плоидности (12C, 24C, 48C).
-
3. Результаты анализа плоидности эндосперма зерновок кукурузы (Zea mays L.), завязавшихся в гетероплоидных скрещиваниях (2n ½ 4n), с помощью проточной цитометрии (г. Саратов, 2022 год)
|
Плоидность полученного Гибридная комбинация растения |
Число ядер Средние значения пиков флюоресценции, ед. |
|
АТ ГПЛ ( n ), лист ( n -стандарт) АТ ГПЛ (2 n ), эндосперм (3 n -стандарт) АТ ГПЛ (2 n ) ½ КРТ (4n), зерновка ¹ 1 4 n F 3 (В47/АТ ГПЛ) (2 n ) ½ ТП (4 n ), зерновка ¹ 2 2 n F 3 (В47/АТ ГПЛ) (2 n ) ½ ТП (4 n ), зерновка ¹ 3 2 n F 3 (В47/АТ ГПЛ) (2 n ) ½ ТП (4 n ), зерновка ¹ 4 2 n F 1 (КМ/Сахарная1) (2 n ) ½ КрТ (4 n ), зерновка ¹ 5 2 n F 1 (КМ/Сахарная1) (2 n ) ½ КрТ (4 n ), зерновка ¹ 7 2 n F 1 (КМ/Сахарная1) (2 n ) ½ КрТ (4 n ), зерновка ¹ 8 2 n В47 (2 n ) ½ ТП (4 n ), зерновка ¹ 9 4 n В47 (2 n ) ½ ТП (4 n ), зерновка ¹ 10 4 n ЧТ (4 n ), эндосперм (6 n -стандарт) |
7781 35 (1C); 70 (2C) 3306 96 (3С); 191 (6C); 388 (12C); 710 (24C) 2607 173 (6C); 340 (12C); 649 (24С); 1230 (48C) 2696 174 (6C); 340(12C); 661 (24С); 1263 (48С) 2604 164 (6C); 321 (12C); 620 (24С); 1134(48С) 2719 173 (6C); 343 (12C); 673 (24С); 1298 (48C) 2171 175 (6C); 349 (12C); 667 (24С); 1286 (48C) 2726 189 (6C); 362 (12C); 704 (24С); 1333 (48С) 2327 181 (6C); 368 (12C); 717 (24С); 1334 (48C) 2765 171 (6C); 349 (12C); 679 (24C); 1298 (48C) 2654 164 (6C); 333 (12C); 643 (24С); 1444 (48C) 1634 182 (6C); 385 (12C); 756 (24C); 1427 (48C) |
Примечание. КМ — Коричневый Маркер, ТП — Тетра-Парий, КРТ — Краснодарский тетраплоид, ЧТ — Черная Тетра.
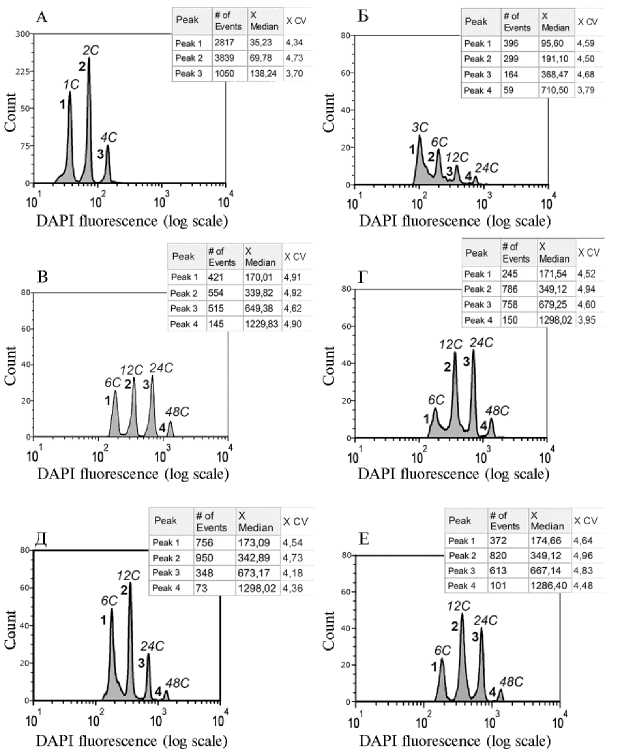
Рис. 3. Плоидность ядер эндосперма выполненных зерновок кукурузы ( Zea mays L.), завязавшихся в гетероплоидных скрещиваниях ( 2 n % 4 n ): А — лист гаплоидного растения линии АТ ГПЛ ( n -стандарт); Б — эндосперм диплоидного растения линии АТ ГПЛ (3 n -стандарт); В — эндосперм выполненной зерновки, завязавшейся на початке АТ ГПЛ (2 n ), опыленном пыльцой КрТ (4 n ), из которой было получено тетраплоидное растение; Г — эндосперм выполненной зерновки, завязавшейся на початке В47 (2 n ), опыленном пыльцой ТП (4 n ), из которой было получено тетраплоидное растение; Д — эндосперм выполненной зерновки, завязавшейся на початке гибрида F 3 (В47/АТ ГПЛ) (2 n ), опыленного пыльцой ТП (4 n ), из которой было получено диплоидное растение; Е — эндосперм выполненной зерновки, завязавшейся на початке гибрида F 1 (КМ/Сахарная1) (2 n ), опыленного пыльцой КрТ (4 n ), из которой было получено диплоидное растение; # of events — число ядер в каждом пике, X Median — средняя по оси X для каждого пика флюоресценции, X CV — ошибка средней для каждого пика по оси X.
В отличие от эндосперма диплоидного растения линии АТ ГПЛ, в эндоспермах большинства выполненных зерновок от скрещиваний 2n ½ 4n средние значения 1-го пика флюоресценции были значительно выше (164189 ед.) и указывали на более высокое содержание ДНК, предположительно соответствовавшее 6С и выше. Такой результат наблюдался у эндосперма зерновок, из которых были получены как тетраплоидные гибриды (см. табл. 3, зерновки ¹¹ 1, 9, 10; рис. 3, В, Г), так и диплоидные F1(КМ1/Сахар-ная1) растения (см. табл. 3, зерновки ¹¹ 2-5, 7, 8; рис. 3, Д, Е). Эти данные указывают, что плоидность эндоспермов таких зерновок значительно превышала 3n. Возможно, эндоспермы развивались на основе слияния нередуцированных полярных ядер (2n + 2n), оплодотворенных диплоидным спермием (2n), в результате чего имели плоидность 6n. В таком случае соотношение родительских геномов в развивающемся эндосперме — 4м:2о, что соответствует соотношению 2м:1о, необходимому для развития полноценного эндосперма (10, 11).
Эти данные подтверждают сформулированную ранее гипотезу о развитии выполненных зерновок в скрещиваниях 2 n ½ 4 n , сформировавшихся из нередуцированных зародышевых мешков (ЗМ) (14, 15). Как известно, формирование нередуцированных ЗМ — это один из компонентов гамето-фитного апомиксиса, в связи с чем полученные данные указывают на наличие этого компонента у ряда изученных линий и гибридов кукурузы. Несколько сниженные значения флюоресценции ядер эндосперма у таких гибридных зерновок по сравнению с флюоресценцией ядер эндосперма тет-раплоида могут быть объяснены частичной элиминацией хромосом, которая могла иметь место в сложном гибридном геноме.
Примечательно, что в случае оплодотворения нередуцированнных ЗМ диплоидной пыльцой гибридным может быть не только эндосперм (6 n ), но и зародыш (4 n ). Действительно, некоторые выполненные зерновки давали такие тетраплоидные растения (см. табл. 2). Тем не менее в ряде случаев из выполненных зерновок развиваются диплоидные растения, причем фенотипы этих растений весьма сильно схожи с фенотипом материнских линий (14, 15). Однако проведенное нами ранее генотипирование «матро-клинных» диплоидов с помощью полиморфных кодоминантных SSR-маркеров, дифференцирующих материнские линии и использованную в скрещиваниях тетраплоидную линию ЧТ, выявило, что такие диплоиды, будучи гомозиготными по материнским маркерам для большинства хромосом, оказываются гетерозиготным по одной-двум хромосомам (а в некоторых случаях и по большему числу хромосом) (16).
Исходя из этих фактов, мы высказали предположение о том, что диплоидная природа растений, развивающихся из выполненных зерновок, формирующихся у некоторых линий кукурузы в 2 n ½ 4 n скрещиваниях, оказывается следствием элиминации хромосом, при этом элиминируют преимущественно хромосомы тетраплоидной отцовской линии (16). Такая элиминация хромосом отцовских родителей — широко распространенное явление при отдаленной гибридизации (21-23), а также при скрещиваниях с линиями-гаплоиндукторами (24-26). В наших экспериментах, однако, в отличие от работ по получению гаплоидов, были получены диплоиды, появление которых, по-видимому, обусловлено наличием нередуцированных ЗМ у материнских линий кукурузы.
Следует отметить, что частота формирования выполненных зерновок в скрещиваниях 2n ½ 4n в наших экспериментах значительно различалась у разных линий и гибридов (см. табл. 2). По-видимому, это следствие влияния генотипа на частоту формирования нередуцированных ЗМ. Однако сильнее интригуют данные о влиянии генотипа опылителя — тетраплоид-ной линии, которая служит «индуктором» этого процесса. Использованные в нашем опыте тетраплоиды различались по своей «индуцирующей» способности (см. табл. 1). Как известно, у линий-гаплоиндукторов индуцирующая способность связана с наличием мутации в гене прицентромерного гистона CENH3, участвующего в прикреплении хромосом к веретену деления и обусловливающего элиминацию хромосом опылителя в гибридном геноме (27, 28). Однако в наших экспериментах наблюдалась не только элиминация хромосом опылителя (16), но и разная частота формирования самих выполненных зерновок при опылении разными опылителями. Очевидно, что это связано с процессами, происходящими на более ранних стадиях развития и определяется, возможно, разной оплодотворяющей способ- ностью спермиев разных тетраплоидов.
Таким образом, при определении плоидности эндосперма выполненных зерновок кукурузы, развившихся в гетероплоидных скрещиваниях (2 n x 4 n ), с помощью проточной цитометрии были получены данные, подтверждающие гипотезу о развитии таких зерновок на основе нередуцированных зародышевых мешков — одного из ключевых элементов апомиктич-ного размножения растений. Эти зерновки содержат гексаплоидный эндосперм и тетраплоидный или диплоидный зародыш гибридного происхождения, у которого преобладают хромосомы материнской линии, но могут присутствовать отдельные хромосомы отцовского родителя. На первый взгляд, полученные результаты указывают на отсутствие диплоидного апомиксиса у изученных линий и гибридов. Однако они показывают наличие у них одного из его основных компонентов — нередуцированных зародышевых мешков. Но наличие только этого компонента оказывается недостаточным для развития апомиктичных растений. Полученные данные свидетельствуют об эффективности применения тетраплоидных опылителей для выявления нередуцированных зародышевых мешков у линий и гибридов кукурузы. Подобный подход может в дальнейшем использоваться в работах по конструированию апомиксиса у этой важной сельскохозяйственной культуры посредством геномного редактирования. Следует также отметить, что полученные нами результаты подкрепляют балансовую теорию развития эндосперма прямыми экспериментальными доказательствами, поскольку исследование плоидности эндосперма у зерновок, развившихся в гетеро-плоидных скрещиваниях, которые использовались для обоснования балансовой теории эндосперма, посредством проточной цитофотометрии нами проведено впервые.
ФГБНУ Федеральный аграрный научный центр Юго-Востока,
