Возобновительные процессы в популяциях широколиственных деревьев, обитающих в лесистых пойменных ландшафтах европейской России

Автор: Браславская Татьяна Юрьевна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Лесные ресурсы
Статья в выпуске: 3-3 т.15, 2013 года.
Бесплатный доступ
В облесенных поймах двух незарегулированных рек, в условиях заповедного режима исследована демографическая структура и плотность подроста широколиственных лесообразующих видов в лесных и нелесных сообществах. Выявлено, что в лесах (краткопоемных и среднепоемных) интенсивно возобновляются ясень (в условиях зоны широколиственных лесов), вяз и липа (в условиях зоны хвойно-широколиственных лесов), но затруднено возобновление дуба. На краткопоемных лугах в обеих поймах через 10-12 лет после прекращения сенокошения (ранее проводившегося ежегодно) интенсивно возобновляется дуб, и практически не возобновляются другие перечисленные широколиственные виды.
Пойменные леса, возобновление леса, широколиственные виды, дуб черешчатый (quercus robur l.), первичные и вторичные сукцессии
Короткий адрес: https://sciup.org/148201899
IDR: 148201899 | УДК: 635.054:556.5
Reproductive processes in populations of broad-leaved woody species, inhabiting in afforested floodplain landscapes of European Russia
In two afforested floodplains of similar plain rivers, within state natural reserves in the both, demographic structure of broad-leaved woody populations was observed in forest, shrub and meadow communities. It was revealed, by comparing of regeneration density, that in forests (inundated for 1-4 weeks annually) recruitment of ash (in nemoral zone), elm and linden (in hemiboreal zone) are high numerous, but reproduction of oak populations is suppressed. Contrary, in both floodplains oak regeneration is numerous in meadows (of the same inundation longevity) about 10-12 years after mowing was ceased, whereas other broad-leaved woody species do not invade in these communities.
Текст научной статьи Возобновительные процессы в популяциях широколиственных деревьев, обитающих в лесистых пойменных ландшафтах европейской России
Изучение закономерностей возобновления лесообразующих древесных видов – одна из важнейших задач лесной экологии. Уточнение этих закономерностей применительно к условиям пойменных территорий имеет большое значение в настоящее время, особенно для Европейской России, где лесной покров в поймах сильно фрагментирован и преобразован в результате сельскохозяйственного и лесопромышленного освоения [9] и нуждается в охране. Поэтому, наряду с изучением возобновления древесных видов в пойменных лесах, требуется выявлять закономерности этого процесса в других типах растительных сообществ, как первичных, так и вторичных по происхождению, поскольку для сохранения популяции вида в пойменном ландшафте может оказаться важным каждый популяционный локус, если перспективы его существования благополучны.
Цель работы: в условиях пойменного ландшафта сравнить интенсивность возобновительных процессов у массовых лесообразующих широколиственных видов в растительных сообществах разных типов – лесах и лугах центральной поймы, ивняках прирусловой поймы.
Результаты сравнительного анализа помогут выявить экологические и ценотические условия, которые необходимы каждому из видов для
устойчивого самоподдержания его ландшафтных популяций в поймах лесного пояса. Для проведения исследований были выбраны поймы, сохранившие высокую облесенность (не менее 80%), на незарегулированных реках сходного географического масштаба (табл. 1), текущих в сходных ландшафтных условиях – по флювиогляциальным зандровым равнинам (полесьям).
Исследуемые поймы относятся к типу сегментно-гривистых, у каждой из них общая (по обоим берегам) ширина варьирует от 2 до 5 км. На низких среднепоемных уровнях прирусловой поймы (продолжительность заливания 15-30 дней [10]) произрастают кустарниковые ивняки асс. Salicetum triandro-viminalis (Tx.1931) Tx. et Lohmeyer 1950 на аллювиальных слоистых дерновых кислых маломощных супесчаносуглинистых почвах; это – интразональные сообщества, представленные в обеих поймах. Состав лесной растительности в исследуемых поймах имеет зональную и региональную специфику [1, 2, 6, 7]. В Неруссо-Деснянском полесье на высоких краткопоемных уровнях центральной поймы (продолжительность заливания не более 10-14 дней [10]) произрастают широколиственные леса асс. Aceri campestris-Quercetum roboris Bulokh. et Solom. 2003 на аллювиальных дерновых лесных кислых среднемощных супесчаных почвах, а на более низких среднепоемных уровнях центральной поймы произрастают широколиственные леса асс. Fraxino excelsioris-Quercetum roboris Bulokh. et Solom. 2003 на аллювиальных луговых лесных кислых маломощных суглинистых и супесчаных почвах. В Марийском полесье на краткопоемных уровнях центральной поймы произрастают темнохвойношироколиственные леса асс. Rhodobryo rosei-Piceetum abietis Korotkov 1991 на аллювиальных дерновых луговых легко- и среднесуглинистых почвах, на среднепоемных уровнях – широколиственные леса асс. Filipendulo ulmariae-Quercetum roboris Polozov et Solomestch 1999 на аллювиальных луговых поверхностнооглеенных тяжелоглинистых почвах. Наиболее массовые лесообразующие широколиственные виды в пойме р. Неруссы (в порядке убывания) – ясень обыкновенный (Fraxinus excelsior L.), дуб че-решчатый (Quercus robur L.) и вяз гладкий (Ulmus laevis Pall.); липа мелколистная (Tilia cordata Mill.) произрастает в этой пойме, но ее распространение и численность незначительны. В пойме р. Большой Кокшаги наиболее массовые лесообразующие широколиственные виды (в порядке убывания) – липа мелколистная, вяз гладкий, дуб черешчатый; ареал ясеня обыкновенного не распространяется на этот регион. Более подробная информация о лесных сообществах в исследуемых поймах, включая параметры древостоев, содержится в ранее публиковавшихся работах [1-3, 6, 7]. Краткопоемные злаковотаволговые луга центральной поймы, произрастающие на аллювиальных дерновых луговых почвах, сходны в обоих регионах по видовому составу растительности и относятся к асс. Lysimachio vulgaris-Filipenduletum ulmariae Bal.-Tul. 1978.
Таблица 1. Физико-географические условия в исследуемых поймах
|
Название реки |
Нерусса |
Большая Кокшага |
|
природно климатическая зона |
широколиственных лесов |
хвойно широколиственных лесов |
|
физико-географический район |
Нерусо-Деснянское полесье |
Марийское полесье |
|
протяженность реки, км |
161 |
178 |
|
площадь бассейна, км2 |
5630 |
6345 |
|
сроки максимума весеннего половодья |
1-3 дня в период с 8 марта по 10 апреля |
1-3 дня в период с 15 апреля по 10 мая |
|
сроки вегетации растительности |
14 апреля – 17 октября |
3 мая – 16 сентября |
|
четвертичные породы, слагающие пойму |
мелкозернистые пески, супеси поверх песков |
тяжелые и средние суглинки поверх мелкозернистых песков |
В обеих поймах сохранились не подвергавшиеся сплошным рубкам обширные лесные выделы с доминированием зональных широколиственных и темнохвойных лесообразующих видов и преобладающим возрастом древостоя 120-150 лет. Наряду с этим, внутри лесных массивов были в прошлом проведены локальные расчистки под луга площадью по несколько гектар, на которых велось сенокошение. В обеих поймах оно было прекращено на рубеже 19801990-х годов, при организации заповедников «Брянский лес» (Неруссо-Деснянское полесье) и «Большая Кокшага» (Марийское полесье). В результате установления заповедного режима краткопоемные луга начали постепенно зарастать древесной растительностью. Информация о дальнейшей спонтанной динамике популяций лесообразующих видов в таких лесных массивах представляет большой интерес, как для выработки методов устойчивого лесопользования, так и для уточнения теоретических представлений о ходе первичных и вторичных сукцессий в лесном покрове.
Исследования проводились: в пойме р Не-руссы – в 1999 г., в пойме р. Большой Кокшаги – в 2003 г. В ходе исследований были заложены временные пробные площади (ПП), размер которых варьировал от 0,04 га до 0,1 га – в зависимости от размеров тех мезоформ ложбинногривистого флювиального рельефа поймы, на которых располагались выбранные сообщества. ПП закладывали в центральной и прирусловой пойме на краткопоемных и среднепоемных уровнях, которые первоначально диагностировали при помощи промера амплитуды высотных отметок гидроуровнем [3], а после выявления соответствий между условиями заливания и растительными сообществами [1-3, 6, 7] – по составу растительности. На ПП выполнены геоботанические описания и проведен сплошной перечет всех древесных видов, в который были включены все экземпляры диаметром 2 см и более (на высоте 1,3 м). Для определения популяционной плотности мелких экземпляров древесных видов (диаметром менее 2 см или высотой менее 1,3 м) на каждой ПП закладывали 5-15 площадок размером 2х2 м. В ходе перечетов на
ПП и на малых площадках для каждого экземпляра определяли онтогенетическое состояние и жизненность, используя морфологические критерии [2, 5]. При обработке полевых данных для каждой ПП рассчитывалась популяционная плотность особей древесных видов в разных онтогенетических состояниях в пересчете на 0,1 га (такая площадь выбрана как соразмерная с наиболее типичной для исследуемых пойм площадью участков, однородных по условиям и составу растительности).
Для сопоставления наиболее массовых широколиственных видов (дуба, липы, вяза, ясеня) по интенсивности возобновления сравнивались по непараметрическому критерию Колмогорова-Смирнова (p<0,01) данные о популяционной плотности их особей подроста (онтогенетические состояния: j – ювенильное, im – имма-турное, v – виргильное, g1 – молодое генеративное). В каждой пойме сравнение проводилось по одинаковым онтогенетическим состояниям как между разными видами внутри одного и того же типа сообществ, так и для одного и того же вида между разными типами сообществ.
Популяционная плотность подроста наиболее массовых широколиственных видов в сообществах различных типов представлена на рис. 1 (пойма р. Неруссы) и рис. 2 (пойма р. Большой Кокшаги. В ходе анализа этих данных количество особей в ювенильном состоянии можно рассматривать в качестве кумулятивного за последние 2-3 года, нивелирующего разногодичные колебания, показателя интенсивности плодоношения родительских деревьев или заноса семян с окружающих территорий. Количество особей более старших онтогенетических состояний характеризует наличие/отсутствие в сообществе условий для продолжения онтогенетического развития новых поколений. По количеству имматурных особей, учитывая данные об их возрасте [5], можно качественно оценить, были ли условия в сообществе благоприятны для вида в течение последних 5-10 лет. По количеству виргильных и молодых генеративных особей можно оценить это для последних 10-30 лет или же примерно определить время начала инвазии вида в сообщество, если оно представляет собой начальную стадию в ходе сукцессии древесной растительности.
В пойме р. Неруссы (рис. 1) оказались статистически значимы следующие особенности возобновления: в краткопоемных и в среднепоемных лесах у ясеня (г, з) плотность ювенильных особей, а у вяза (в, ж) – плотность имматур-ных особей больше, чем у остальных видов. В прирусловых кустарниковых ивняках плотность ювенильных и имматурных особей липы (о)
гораздо ниже, чем у всех остальных широколиственных видов, а у ясеня (р) и дуба (н) выше, чем у вяза (п). На краткопоемных лугах плотность ювенильных особей дуба (и) и ясеня (м) выше, чем у вяза (л) и липы (к), плотность имма-турных особей дуба выше, чем у всех остальных видов, а среди последних самая низкая плотность имматурных особей – у липы. При сопоставлении разных типов сообществ значимы только отличия лугов от всех типов древесной растительности – по плотности имматурных особей дуба.
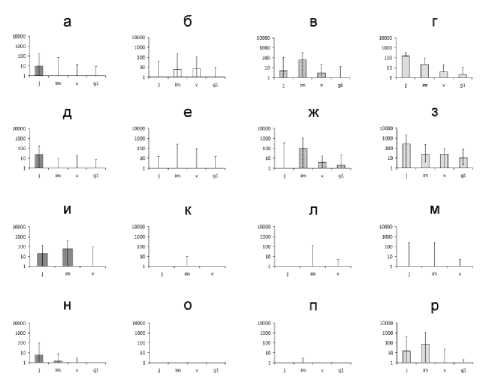
Рис. 1. Онтогенетический состав и популяционная плотность подроста у наиболее массовых широколиственных видов в пойме р. Неруссы: по вертикальной оси – плотность особей (шт./0,1 га), по горизонтальной оси – индексы онтогенетических состояний древесных растений (см. пояснения в тексте). На гистограммах высота столбиков соответствует выборочной медиане популяционной плотности (столбик не изображен, если значение медианы – 0) , размах варьирования – от минимума до максимума. Типы растительных сообществ: а-г – краткопоемные леса центральной поймы (15 ПП), д-з – среднепоемные леса центральной поймы (13 ПП), и-м – краткопоемные луга центральной поймы (23 ПП), н-р – среднепоемные кустарниковые ивняки прирусловой поймы (8 ПП). Лесообразующие виды: а, д, и, н – дуб; б, е, к, о – липа; в, ж, л, п – вяз; г, з, м, р – ясень
Таким образом, в лесных сообществах этой поймы дуб уступает ясеню и вязу в интенсивности возобновления уже на самом начальном его этапе – появлении всходов, хотя выживаемость всходов дуба еще не настолько зависит от режима освещенности в сообществе, как у его более старших онтогенетических состояний [6]. И даже если низкая плотность ювенильного дуба в лесных сообществах – результат кратковременной флюктуации, то сильное отставание от ясеня и вяза по плотности имматурных и вирги-нильных особей все равно свидетельствует, что возобновление дуба под пологом пойменных лесов менее эффективно, чем у этих видов. В прирусловые кустарниковые ивняки (то есть в сообщества начальных стадий классической пойменной первичной сукцессии) вяз, видимо, внедряется не так массово, как ясень и дуб, но и в этих сообществах наиболее интенсивно и эффективно возобновляется не дуб, а ясень. Однако для дуба наличие негативных тенденций в самоподдержании в составе сформировавшейся древесной растительности компенсируется наличием лугов, где прекращено сенокошение. Дуб не только может благополучно развиваться в этих условиях до имматурного состояния (кое-где к моменту проведения исследований – уже и до виргинильного), но и значительно опережает остальные широколиственные виды по интенсивности своей инвазии на луга. Можно предположить, что два фактора обеспечивают дубу такую возможность – запасание его плодов мышевидными грызунами, прячущими желуди в густой траве на лугах [8], и роющая деятельность кабанов на лугах, в результате которой уменьшается проективное покрытие травяного покрова и к почве поступает больше света [4].
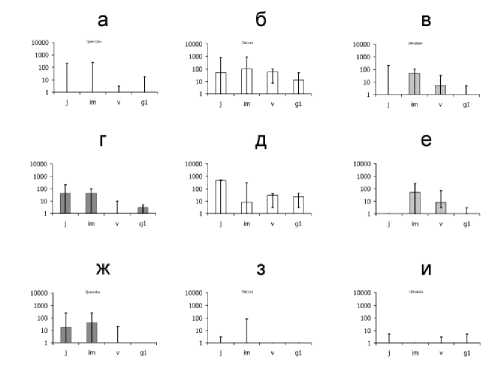
Рис. 2. Онтогенетический состав и популяционная плотность подроста у наиболее массовых широколиственных видов в пойме р. Большой
Кокшаги:
обозначения на гистограммах – такие же, как на рис. 1. Типы растительных сообществ: а-в – краткопоемные леса центральной поймы (22 ПП), г-е – среднепоемные леса центральной поймы (5 ПП), ж-и – краткопоемные луга центральной поймы (10 ПП). Лесообразующие виды: а, г, ж – дуб; б, д, з – липа; в, е, и – вяз
В пойме р. Большой Кокшаги (рис. 2) выявлены следующие значимые отличия: в краткопоемных лесах – плотности имматурных, вирги-нильных и молодых генеративных особей липы (в) от аналогичных особей дуба (а) и вяза (б), а на лугах – плотности имматурных особей дуба
(ж) от имматурных особей вяза (з) и липы (и). В сопоставлении сообществ разных типов значима только разница между лугами и лесами всех типов по плотности липы (б, д, з) и вяза (в, е, и) во всех онтогентических состояниях.
Полученные данные (рис. 2, а, г) согласуются с ранее публиковавшейся информацией о том, что в пойме Большой Кокшаги возобновление дуба в среднепоемных лесах идет успешнее, чем в краткопоемных [6]. Это может быть обусловлено тем, что в среднепоемных условиях снижается интенсивность возобновления липы (рис. 2, д) или более частым выпадением деревьев из древостоя из-за достаточно длительного переувлажнения и образованием просветов в пологе древесных крон. Кроме того, в этой пойме тоже выявляется значительное преимущество дуба перед остальными широколиственными видами в ходе зарастания лугов.
Наблюдаемое активное участие ясеня (в пойме р. Неруссы) в первичной сукцессии древесной пойменной растительности и дуба (в обеих поймах) – в после-луговой вторичной сукцессии позволяет, возможно, понять причины широкого распространения в европейской широколиственной зоне именно дубовых и дубовоясеневых лесов.
Список литературы Возобновительные процессы в популяциях широколиственных деревьев, обитающих в лесистых пойменных ландшафтах европейской России
- Браславская, Т.Ю. Влияние режима речного стока на формирование и динамику пойменных лесных сообществ на реках с незарегулированным стоком//Оценка влияния изменения режима вод суши на наземные экосистемы. -М.: Наука, 2005. С. 109-134.
- Браславская, Т.Ю. Материалы к характеристике онтогенеза и популяционной динамики лесообразующих видов в пойменных лесах заповедника «Большая Кокшага»/Т.Ю. Браславская, М.В. Харлампиева, Т.В. Скоморохова и др.//Научные труды зап. «Большая Кокшага». -Йошкар-Ола: МарГТУ, 2011. Вып. 5. С. 109-126.
- Браславская, Т.Ю. Градиентный анализ флористического состава и экологической структуры травяного яруса в пойменных лесах//Известия Самарского НЦ РАН. 2012. Т. 14. № 1(5). С. 1202-1205.
- Горнов, А.В. Природные и антропогенные механизмы поддержания биологического разнообразия влажных внутрилесных лугов Неруссо-Деснянского полесья. Автореф. дисс. … канд. биол. наук. -М.: Московский педагогич. гос. ун-т, 2010. 22 с.
- Чистякова, А.А. Диагнозы и ключи возрастных состояний лесных растений. Деревья и кустарники: методические разработки для студентов биологических специальностей/А.А. Чистякова, Л.Б. Заугольнова и др. -М.: Изд-во Прометей МГПИ им. В.И. Ленина, 1989. 102 с.
- Евстигнеев, О.И. Популяционная организация и антропогенные преобразования пойменной дубравы реки Большая Кокшага/О.И. Евстигнеев, М.И. Почитаева, С.Е. Желонкин//Бюлл. МОИП. Отд. биол. 1993. Т. 98. Вып. 5. С. 80-87.
- Евстигнеев, О.И. Эрозионно-аккумулятивная деятельность реки и циклические сукцессии пойменной растительности/О.И. Евстигнеев, Т.Ю. Браславская//Биогеоценотический покров Неруссо-Деснянского полесья: механизмы поддержания биологического разнообразия. -Брянск: Типография «Бумполиграфинформ», 1999. С. 71-96.
- Евстигнеев, О.И. Методы оценки дальности разноса семян животными в лесном покрове//Методические подходы к экологической оценке лесного покрова в бассейне малой реки. -М.: Т-во научных изданий КМК, 2010. С. 97-99.
- Липатова, В.В. Растительность пойм//Растительность Европейской части СССР. -Л.: Наука, 1980. С.346-372.
- Раменский, Л.Г. Введение в комплексное почвенно-геоботаническое изучение земель. -М.: Огиз, Сельхозгиз, 1938. 620 с.
