Возобновленные кладки как один из механизмов поддержания численности популяции птиц в период размножения

Автор: Дьяконова Ирина Владимировна, Дьяконов Юрий Владимирович
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Наземные экосистемы
Статья в выпуске: 5-1 т.16, 2014 года.
Бесплатный доступ
Предпринята попытка экспериментальным путем изучить один из механизмов сохранения численности популяции птиц через возобновляемые яйцекладки в неблагоприятный период размножения, когда наблюдается массовая гибель основных яйцекладок птиц.
Птицы, основные яйцекладки, возобновленные яйцекладки, размер кладки, репродуктивная система, успешность вылупления
Короткий адрес: https://sciup.org/148203363
IDR: 148203363 | УДК: 598.2:591.525
Renewed laying as a mechanism for supplying of the population of birds in the period of breeding
The paper refers to the study of one of the mechanisms for preserving of birds population through renewable laying during unfavourable breeding period experimentally when there is a mass death of the main egg-laying birds.
Текст научной статьи Возобновленные кладки как один из механизмов поддержания численности популяции птиц в период размножения
В отличие от повторных кладок, которые относятся ко второй потенции размножения одного репродуктивного сезона, основные и возобновленные яйцекладки - это яйцекладки одной потенции размножения птиц. Возобновленные кладки возникают в результате гибели основных яйцекладок под воздействием внешних факторов [1, 2, 3,4]. Поэтому, возобновленные кладки следует рассматривать, на наш взгляд, как компенсацию элиминации основных кладок. То есть они являются механизмом поддержания численности популяции при воздействии неблагоприятных факторов на основные яйцекладки.
В экспериментальных работах по изучению основных и возобновленных кладок особое значение имеют методики получения возобновленных яйцекладок. Эти методики гарантируют экспериментальное получение именно возобновленных кладок и позволяют исследовать параметры возобновленных яйцекладок на фоне основных.
МАТЕРИАЛ И МЕТОДЫ
В качестве модельного вида нами был выбран грач ( Corvus frugilegus L.). С нашей точки зрения этот вид является наиболее подходящим для отработки методики получения возобновленных кладок.
Исследования проводились в двух поселениях грачей, находящихся близ станции Курашимский Свердловской железной дороги. Одно из них состояло из 200-250 гнезд, размещенных преимущественно на елях на высоте 10-15 метров. Второе находилось в 3 км от первого, их разделял лесной массив. Это поселение в разные годы насчитывало только 30-50 гнезд, сооруженных, как и в первом случае, на елях примерно той же высоты. В связи с тем, что индивидуальное мечение грачей,
которое бы позволило опознать особь на значительном расстоянии очень затруднено из-за особенностей их перьевого покрова, основные работы по искусственному стимулированию птиц к возобновлению кладок проводилось на малочисленной колонии, где большинство гнезд было доступно для исследования. С этой целью одновременно изымались все полные кладки, которые считались завершенными, если в течение трех дней число яиц в ней оставалось неизменным.
Гнезда грачей посещали ежедневно в первой половине дня, картировали, помечали биркой. Каждое очередное снесенное яйцо метили быстросохнущей краской.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Полученные нами результаты показали, что у грача к возобновлению кладки самки приступают примерно через 3-5 дней после утраты основной яйцекладки. В нашем эксперименте в этом приняли участие более 80% самок. Для этой цели грачи использовали, как правило, старые гнезда и лишь некоторые из них были разобраны самими птицами при строительстве новых.
Размер возобновленных кладок – 2,7 я/гн., был на единицу меньше, чем основных – 3,7 я/гн. У самок, приступивших к возобновлению первыми, начальная плодовитость составила 2,5 я/гн. Данный показатель возрастал по мере повышения активности яйцекладки и достиг самого высокого значения – 3,4 я/гн. в период ее пика. Размер яйцекладок, начало которых совпало с периодом снижения активности и температур, оказался самым низким – 2,3 я/гн.
Таким образом, в период возобновления сохранилась та же тенденция, что и в основном – увеличение среднего размера кладок в период пика активности, совпадающего с повышением температур и снижение его у последних кладок.
При возобновлении изменилось соотношение ритмичных и аритмичных кладок в сторону увеличения первых – 82,6% (табл. 1). При этом их размер – 2,9 я/гн. превышал таковой показатель аритмичных – 2,5 я/гн., которые преобладали лишь в начале возобновления. В период пика все кладки (100%) были ритмичными. В результате увеличения ритмичных кладок темп возобновления яйцекладок составил 10 яиц в день, значительно превысив таковой в основной.
Таблица. Соотношение ритмичных и аритмичных кладок и их размеров у грача
|
Тип кладки |
о 5 о м о ы |
^5 S 2 S |
^5 у S S 2 S |
Размер кладок |
|
|
о 5 2 S |
< щ |
||||
|
Основные кладки |
47/100 |
30/63,9 |
17/36,2 |
2,9 |
3,9 |
|
Возобновл. кладки |
28/100 |
19/67,2 |
9/32,1 |
3,5 |
3,6 |
Нам удалось проследить динамику изменения наседного пятна и репродуктивной системы самок с учетом основной и возобновленной кладок. Так, перед началом яйцекладки (рис. 1) размер наседного пятна у одной из самок составил 4,0х6,7 см, кожа в этом месте была тонкой, с легким шелушением, перьевой покров выпал полностью. В этот период масса яичника достигла 4,0 г, яйцевода – 12,7 г при длине в 32,0 см, что указывало на ее готовность к основному циклу размножения.
При откладке первого яйца размер наседного пятна остается примерно в тех же пределах, в данном случае он был равен 3,7х5,9 см, но заметные изменения произошли в его структуре: кожа стала тонкой, сухой, отчетливо были видны кровеносные сосуды, без шелушения. Увеличилась масса яичника – 6,5 г и яйцевода 17,9 г при длине 36,0 см.
После откладки второго яйца (третье находилось в средней части яйцевода и имело частичную белковую оболочку) площадь наседного пятна увеличилась до 5,4х6,7 см (рис. 2). Кожа при этом осталась сухой, без шелушения, но морщинистой, что является верным признаком начала накопления лимфы. Масса яичника составила 4,3 г, яйцевода 17,2 г при увеличении его размера – 38,0 см. Во время завершения основной кладки (4 яйца) хорошо просматривается наполнение кожных структур лимфой (рис. 3), верхний слой кожи начинает шелушиться, размер наседного пятна – 5,2х9,0 см. Масса яйцевода – 25,8 г и длина 44,0 см, значительно увеличены, а масса яичника составила всего 1,2 г. Через двое суток после завершения кладки (рис. 4) при оптимальных размерах наседного пятна – 5,8х10,2 см кожа становится глянцевой, влажной на ощупь, бугорчатая в результате интенсивного наполнения лимфой. Резко сократилась масса яичника – 0,46 г, яйцевода – 5,6 г и его размер – 22,8 см.

Рис. 1. Наседное пятно и репродуктивная система самки грача перед основной яйцекладкой
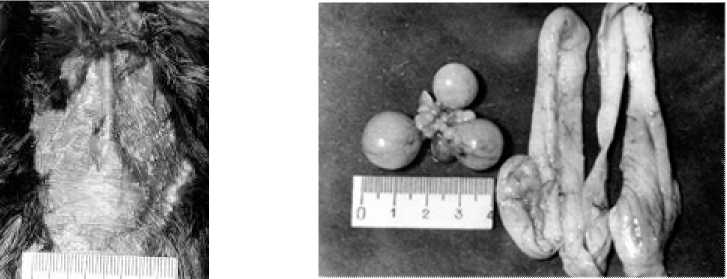
Рис. 2. Наседное пятно и репродуктивная система самки грача при откладке второго яйца
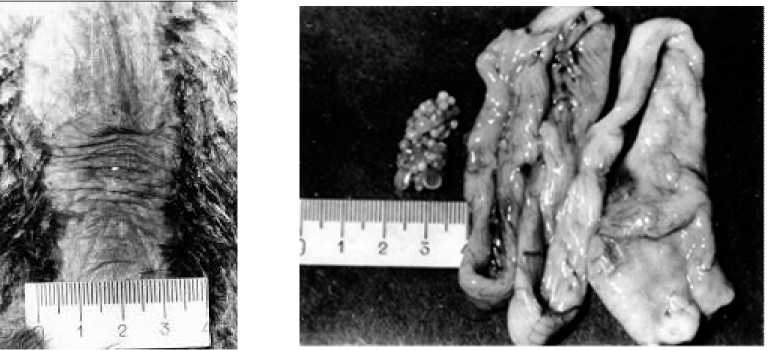
Рис. 3. Наседное пятно и репродуктивная система самки грача при завершении основной яйцекладки
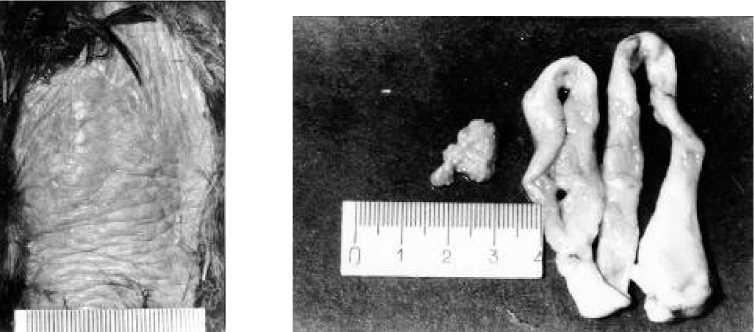
Рис. 4. Наседное пятно и репродуктивная система самки грача через двое суток после завершения основной яйцекладки
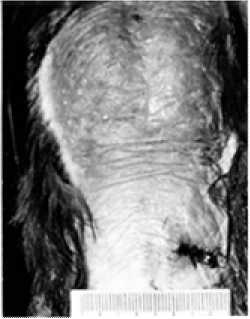
Рис. 5. Наседное пятно самки грача перед возобновлением яйцекладки
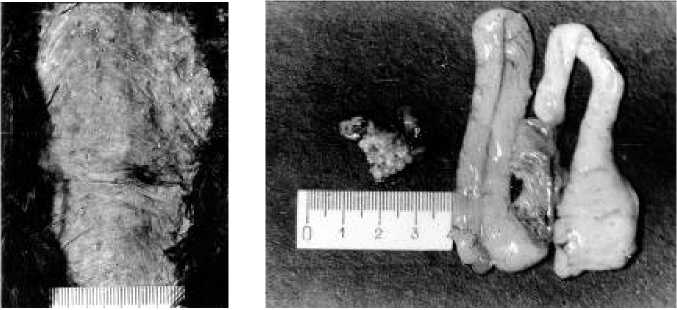
Рис. 6. Наседное пятно и репродуктивная система самки грача через двое суток после завершения возобновленной яйцекладки
Исследование самки перед возобновлением кладки выявило некоторое увеличение площади наседного пятна – 6,2×10,7 см в большей степени за счет брюшного участка (рис. 5). На коже, заполненной лимфой, вновь появилось шелушение, характерное для второй половины периода основной яйцекладки. Масса яичника в это время увеличилась до 1,8 г, яйцевода 15,9 г при длине 31,0 см.
К моменту завершения возобновления кладки (четвертое яйцо находилось в нижней части яйцевода) размер наседного пятна составил 5,8×11,9 см, кожа продолжала сильно шелушиться (рис. 6). Масса яичника равнялась 1,3 г, яйцевода 15,1 г, а его длина – 38,5 см. По-видимому, по характеру шелушения кожи в области наседного пятна можно с определенной достоверностью судить о месте яйцекладки в цикле размножения.
Данные показали, что в период основной яйцекладки используется лишь некоторая часть фолликул, большая же из них остается резервной, и, как известно, недоиспользуется в обычных условиях одного сезона размножения. Причем, судя по «зернистости яичника», фолликулы довольно долго остаются в состоянии готовности к их дальнейшему развитию и превращению в яйцекладку. Этот резерв является одной из составных частей той основы, которая позволяет птицам при гибели кладок приступать к их двух, трех и более раз возобновлению. По-видимому, это и есть та биологическая основа, которая позволила человеку вывести продуктивные породы кур.
Успех вылупления в основных и возобновленных кладках грача нами изучался в течение двух лет. В первый год под наблюдением находилось 13 основных и 14 возобновленных кладок, средний размер которых был одинаков – 3,5 я/гн. В основном цикле размножения во всех находящих-
ся под наблюдением гнездах успешно вылупилось 26 птенцов или по 2,0 птенца на гнездо. В возобновленном цикле в среднем на гнездо приходилось по 2,6 птенца. Следовательно, успех вылупления в возобновленных кладках, – 75,0%, был выше, чем в основных – 68,4%.
На второй год кладки в сравниваемых группах также были идентичны по среднему уровню начальной плодовитости – 3,6 я/гн. В основных кладках в среднем приходилось по 2,5 вылупившихся птенца, в возобновленных значительно выше – 2,9 птенца на гнездо. Успех выупления составил соответственно 67,9% и 79,3%.
Сопоставляя исследуемые показатели за два года, следует отметить, что в последний год количество птенцов, успешно вылупившихся в среднем на одно гнездо, было больше, как из основных, так и из возобновленных кладок. Однако, даже и в этот год успешность вылупления в этих кладках не превышала соответствующий показатель в возобновленных предыдущего года. Таким образом, возобновленные кладки могут играть значительную роль в пополнении численности популяции. Наибольшее значение они, несомненно, приобретают в годы массовой гибели основных кладок в экстремальных условиях.
Список литературы Возобновленные кладки как один из механизмов поддержания численности популяции птиц в период размножения
- Мальчевский А.С. Гнездовая жизнь певчих птиц. Изд-во ЛГУ, 1959. 281 с.
- Нумеров А.Д. К биологии зимородка в Окском заповеднике//VII Всесоюзная орнитологическая конференция. Киев, 1977. Ч. 1. С. 292-293.
- Степанян Л.С. Случаи атипического размножения у некоторых воробьиных птиц//Бюллетень МОНП. 1962. Том XVII. Вып. 1. С. 119-120.
- Шварц С.С. Экологические закономерности эволюции. М.: Наука, 1980. 279 с.
