Возрастной аспект в исследовании роста ели под пологом березняков подзоны южной тайги европейской части России

Автор: Дерюгин А.А.
Журнал: Лесохозяйственная информация @forestry-information
Рубрика: Лесоведение и лесоводство
Статья в выпуске: 4, 2020 года.
Бесплатный доступ
Рассматривается рост подпологовой еловой популяции в березняках южной тайги в связи с возрастом деревьев ели. Для возрастных генераций ели установлены некоторые закономерности динамики ряда таксационных показателей и среднепериодических приростов, приведено их сравнение с данными существующих таблиц хода роста насаждений ели
Южная тайга, березовые леса, подпологовые популяции ели, возраст насаждений, рост деревьев ели
Короткий адрес: https://sciup.org/143173000
IDR: 143173000 | УДК: 630.221.2:630.561 | DOI: 10.24419/LHI.2304-3083.2020.4.02
Age aspect in the study of spruce growth under the canopy of birch forests in the southern taiga of the European part of Russia
The study of growth or of different ages under the canopy of 70-year-old birch by North LOS Institute of Forest Science RAS in the two test areas. 296 model spruce trees were cut down here. Their age varied from 21 to 66 years, height - from 1 to 17 m. Birch forests belong to stands with a late renewal of spruce (the period of renewal of the main part of the population - 35-40 years, age 31-40 years). The age-related generations of 21-30, 41-50, and 51-60 years are less represented. The growth of spruce was analyzed based on measurements of model trees with their division into four 10-year generations. The analysis of average periodical (over a 5-year period) increases showed the following. In the studied phytocenoses, with the late renewal of spruce, a small part of the spruce population (age generation 51-60 years) does not significantly affect the growth of the next two generations (31-40 and 41-50 years). This is explained by the small density of crowns (0.14) of the trees of the first generation. Crowns of trees of the second generation increase the closeness to 0,38. However, this did not significantly affect the growth of trees for 31-40 years. The regeneration and subsequent growth of generation for 21-30 years occurred with a crown density of 0,87. This has a negative impact on growth. Studies have not established a significant difference in the growth of the first three generations of spruce. During the first 50 years of growth and development, all age-related generations in the spruce population differ in the value of taxational indicators from the values in the existing growth tables. To adequately reflect the growth of the spruce population in the birch forests of the southern taiga, it is necessary to develop a model for the formation of such stands.
Текст научной статьи Возрастной аспект в исследовании роста ели под пологом березняков подзоны южной тайги европейской части России
Березняки – одна из наиболее распространенных растительных формаций южной тайги. Их доля в землях лесного фонда постоянно увеличивается, что является следствием сокращения рубок ухода в молодых мелколиственных насаждениях, формирующихся после рубки древостоев. В подзоне южной тайги европейской территории страны площадь древостоев с преобладанием мелколиственных пород составляет около 20 млн га. Березняки представлены на 80 % этой территории. На значительной площади (75 %) под пологом таких древостоев имеются подрост или второй ярус ели [1]. Эта предварительная генерация может стать основой для восстановления коренных ельников. Рост популяции ели рассматривается, главным образом, в связи с влиянием рубки мелколиственных древостоев. В 1980-х гг. для северной и средней таежных подзон европейской части России были разработаны таблицы хода роста березово-еловых древостоев с елью последующего возобновления для разных типов леса [2]. В них приводятся средние значения основных
таксационных характеристик ели в березняках разного возраста. Подобные таблицы для подзоны южной тайги отсутствуют.
В ранних публикациях рассматривались закономерности роста подпологовой популяции ели в связи с периодом возобновления, высотными и объемными характеристиками деревьев [3–6]. Тем не менее, несмотря на имеющуюся информацию, рост подпологовой ели в зависимости от возраста при естественном формировании березняков остается недостаточно изученным.
Объекты и методы
Изучение роста подпологовой ели разного возраста проводилось на Северной ЛОС Института лесоведения РАН в Рыбинском районе Ярославской обл. С этой целью были заложены 2 пробные площади (ПП) в березняках 70-летнего возраста с подпологовой елью. Насаждения относятся к кислично-черничной группе типов леса. Для изучения хода роста на ПП было вырублено 296 деревьев ели. Диапазон таксационных характеристик взятых моделей приведен в табл. 1.
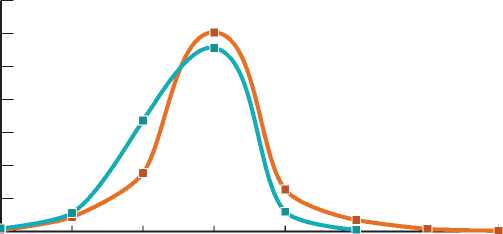
1-10 11-20 21-30 31-40 41-50 51-60 61-70 71-75
Возраст ели, лет
ПП 1 ПП 2
Рис. 1. Распределение деревьев ели на ПП по 10-летним возрастным группам
Анализ возрастной структуры ели показал, что на ПП она различается несущественно (рис. 1). Учитывая это обстоятельство и сопоставимый возраст березы (около 70 лет), при анализе роста подпологовой ели разного возраста все взятые на ПП модели были объединены в один массив. Подпологовая ель на ПП относится к популяции с поздним возобновлением – основная часть популяции появляется в березняках старше 30-летнего возраста. При раннем возобновлении это происходит в березняках возрастом до 20 лет [7].
Ель, произрастающая под пологом мелколиственных древостоев, в первые годы жизни характеризуется очень замедленным ростом [5, 8]. Кроме того, часть отобранных моделей имеет небольшие размеры. В связи с этим принятые в таксации схемы раскряжевки стволов для анализа роста в данном случае неприемлемы. Для анализа спилов была принята следующая схема: до высоты 1,3 м их брали через 0,25 м, далее через 0,5 м или 1 м в зависимости от текущего прироста ели в высоту. Спилы обрабатывали по принятым в таксации методам [9]. Показатели роста каждого дерева (высота, диаметр стволов, объем стволов, средние и текущие приросты) определяли по 5-летним возрастным периодам с применением специально разработанной программы.
Результаты и обсуждение
Анализ роста ели проводили по наиболее представительным возрастным поколениям и модельным деревьям, имеющим более высокий возраст. Это дает возможность получить статистически достоверные данные за относительно длительный временной интервал. На предварительном этапе все модельные деревья были распределены по 10-летним возрастным группам, в которых определены основные
морфометрические характеристики в год взятия моделей (табл. 2).
Эти характеристики моделей в возрастных генерациях достоверно различались при уровне значимости 0,05 (Т0,05 = 2,00 < Tфакт. = 2,30–8,30), что объясняется не только возрастом елей, но и спецификой условий роста разных возрастных поколений, связанных с внутривидовыми отношениями. Данное утверждение можно проиллюстрировать на примере динамики высоты елей разных возрастных поколений в зависимости от возраста березы. По мере увеличения возраста березняка и нарастания внутривидовой конкуренции разница в высоте между поколениями постепенно возрастает (рис. 2).
Из графика следует, что ни одна из рассматриваемых возрастных групп деревьев в березняках до 60-летнего возраста, т.е. до возраста количественной спелости, не может выйти в первый ярус древостоя. Высота березы существенно больше высоты ели самой старшей генерации (51–60 лет).
С целью установления различий в росте ели подпологовой популяции разного возраста была проанализирована динамика высоты и диаметра ствола на высоте 0,1 м по четырем наиболее представительным возрастным группам модельных деревьев. Установлено, что возрастные изменения рассматриваемых характеристик
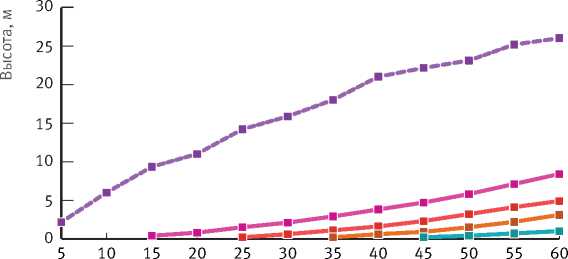
Возраст березняка, лет
Ель 21-30 Ель 31-40
Ель 51-60 Береза
Ель 41-50
Рис. 2. Изменение высоты березы и ели в зависимости от возраста березняка
5 10 15 20 25 30 35 40 45 50 55
Возраст ели, лет
21-30 31-40 41-50 51-60
Рис. 3. Динамика высоты деревьев ели разных возрастных групп
у возрастных групп 31–40 и 41–50 лет протекают идентично (рис. 3 и 4).
Для этих двух групп различие значений основных таксационных характеристик в возрасте 25 лет недостоверно при уровне значимости 0,05 (табл. 3). По отношению же к двум другим группам различие достоверно – фактические значения Т-критерия (Tфакт.) равны или больше табличных значений при уровне значимости 0,05.
Соотношение роста деревьев ели в рассматриваемых возрастных генерациях подтверждается анализом среднепериодических (за 5 лет) приростов. В возрастной группе 21–30 лет приросты по высоте, диаметру и площади поперечного сечения стволов на высоте 0,1 м с 10 лет, а по объему ствола – с 15 лет начинают существенно уступать приросту деревьев более старших возрастных генераций (табл. 4). В двух следующих возрастных группах (31–40 и 41–50 лет) в пределах сопоставимых возрастов значения приростов по всем показателям практически одинаковы. Небольшие различия приростов по объему ствола в этих группах не имеют принципиального значения и статистически недостоверны (в возрасте 35 лет T0,05 = 1,96>Tфакт. = 0,80).
Наибольшими значениями приростов в конкретном возрасте ели отличаются деревья возрастной группы 51–60 лет (см. табл. 4).
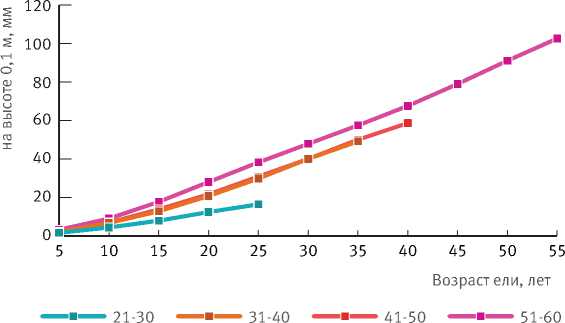
Рис. 4. Динамика диаметра стволов деревьев ели на высоте 0,1 м в разных возрастных группах
Таблица 3. Достоверность различия характеристик деревьев разных возрастных групп в возрасте 25 лет
|
Сравниваемые возрастные генерации |
Фактическое значение Т-критерия для характеристик |
|||
|
высота, м |
диаметр ствола, мм, на высоте |
объем ствола, дм3 |
||
|
0,1 м |
1,3 м |
|||
|
21–30 и 31–40 |
5,20 |
6,74 |
- |
2,40 |
|
31–40 и 41–50 |
0,75 |
0,63 |
0,21 |
0,87 |
|
41–50 и 51–60 |
2,18 |
2,02 |
2,02 |
2,22 |
|
21–30 и 31–50 |
4,93 |
6,71 |
- |
2,49 |
|
31–50 и 51–60 |
2,02 |
2,85 |
2,02 |
2,21 |
Примечание. Табличное значение Т-критерия на уровне значимости 0,05 при имеющихся степенях свободы более 40 равно 2,02.
Таблица 4. Среднепериодический прирост деревьев ели в высоту, по диаметру, площади поперечного сечения и объему ствола в разных возрастных группах
|
Возрастная группа, лет |
Возраст ели, лет |
||||||||||
|
5 |
10 |
15 |
20 |
25 |
30 |
35 |
40 |
45 |
50 |
55 |
|
|
Прирост ели в высоту (ZtH), см/год |
|||||||||||
|
21–30 |
5 |
5 |
6 |
6 |
5 |
- |
- |
- |
- |
- |
- |
|
31–40 |
5 |
8 |
9 |
11 |
15 |
17 |
16 |
- |
- |
- |
- |
|
41–50 |
4 |
8 |
10 |
11 |
14 |
17 |
18 |
16 |
- |
- |
- |
|
51–60 |
7 |
9 |
12 |
14 |
16 |
18 |
20 |
23 |
26 |
27 |
22 |
|
Прирост ели по диаметру ствола на высоте 0,1 м (ZtD), мм/год |
|||||||||||
|
21–30 |
0,3 |
0,6 |
0,7 |
0,9 |
0,8 |
- |
- |
- |
- |
- |
- |
|
31–40 |
0,4 |
1,0 |
1,2 |
1,6 |
1,8 |
2,0 |
1,8 |
- |
- |
- |
- |
|
41–50 |
0,4 |
1,0 |
1,4 |
1,6 |
1,8 |
1,8 |
2,0 |
1,8 |
- |
- |
- |
|
51–60 |
0,6 |
1,2 |
1,7 |
2,0 |
2,1 |
2,2 |
2,3 |
2,3 |
2,3 |
2,4 |
2,3 |
|
Прирост ели по площади поперечного сечения ствола на высоте 0,1 м (ZtG), мм2/год |
|||||||||||
|
21–30 |
1 |
3 |
7 |
14 |
18 |
- |
- |
- |
- |
- |
- |
|
31–40 |
1 |
7 |
19 |
43 |
72 |
110 |
126 |
- |
- |
- |
- |
|
41–50 |
1 |
7 |
23 |
45 |
75 |
113 |
129 |
154 |
- |
- |
- |
|
51–60 |
2 |
11 |
36 |
74 |
106 |
130 |
159 |
197 |
262 |
324 |
350 |
|
Прирост ели по объему ствола (ZtV), см3/год |
|||||||||||
|
21–30 |
1 |
1 |
4 |
15 |
33 |
- |
- |
- |
- |
- |
- |
|
31–40 |
1 |
2 |
12 |
40 |
110 |
240 |
380 |
- |
- |
- |
- |
|
41–50 |
1 |
3 |
15 |
47 |
127 |
271 |
466 |
622 |
- |
- |
- |
|
51–60 |
1 |
10 |
45 |
119 |
239 |
399 |
643 |
971 |
1564 |
2333 |
2681 |
Однако значения рассматриваемых показателей характеризуются значительной изменчивостью: коэффициент вариации составляет 47–90 %. Это обусловливает существенные ошибки средних значений и, соответственно, недостоверные различия в границах одинаковых возрастов (T0,05 = 2,00 > Tфакт. = 0,68–1,99). Из вышеизложенного следует, что в изучаемых фитоценозах при позднем возобновлении ели под пологом березняков немногочисленная часть популяции ели, появившаяся первой (в березняке возрастом менее 30 лет), не оказывает существенного влияния на прирост деревьев двух последующих генераций ели. Прежде всего это объясняется небольшой сомкнутостью крон деревьев первой возрастной генерации (0,14) при среднем расстоянии между деревьями 7,5 м. Сомкнутость крон деревьев второй генерации увеличивается до 0,38, а расстояние между деревьями сокращается до 3,5 м. Тем не менее это не так сильно повлияло на приросты деревьев ели генерации 31–40 лет. Возобновление и последующий рост генерации 21–30 лет происходили уже при сомкнутости крон 0,87, что существенным образом сказалось на росте деревьев.
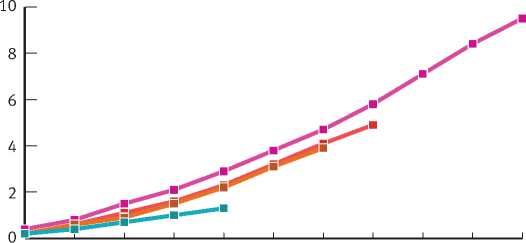
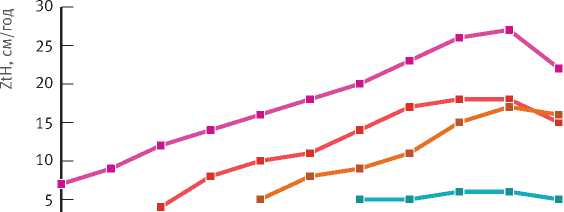
Несущественность различия приростов деревьев ели трех старших возрастных генераций можно проиллюстрировать на примере динамики среднепериодического прироста в высоту в зависимости от возраста березняка. У деревьев этих возрастных групп прирост увеличивается практически синхронно до возраста березы 60–65 лет (рис. 5). По достижении этого возраста (т. е. в березняке, прошедшем возраст количественной спелости) отмечается снижение прироста в высоту у деревьев всех возрастных генераций.
Снижение прироста в высоту и по диаметру на высоте 0,1 м в выделенных группах наступает в разном возрасте – от 25 лет в младшей до 55 лет в старшей группе. Однако это не приводит к снижению прироста по объему ствола, что объясняется увеличением прироста площади поперечного сечения стволов (см. табл. 4).
К настоящему времени разработан ряд таблиц хода роста еловых и березово-еловых насаждений, в частности «Ход роста нормальных еловых насаждений» [10], «Ход роста полных
20 25 30 35 40 45 50 55 60 65 70
Возраст березняка, лет
21-30 31-40 41-50 51-60
Рис. 5. Изменение среднепериодического прироста в высоту деревьев ели разного возраста в зависимости от возраста березняка
двухъярусных березово-еловых древостоев с последующим возобновлением ели подзоны средней тайги» [2]. Нами были сопоставлены некоторые таксационные показатели исследуемых насаждений со значениями в вышеприведенных таблицах хода роста. Учитывая производительность ельников и березняков, наиболее распространенных в регионе исследований, для сравнения в таблицах хода роста взяты следующие значения: нормальные насаждения ели II класса бонитета, полные двухъярусные березово-еловые древостои I класса бонитета. Соотношения полученных значений с показателями в таблицах хода роста приведены в табл. 5.
Значения показателей изучаемых фитоценозов значительно уступают таковым в нормальных насаждениях ели. У старшей возрастной генерации разница менее значима, чем у молодых генераций. В старшей генерации (возрастная группа 51–60 лет) по мере увеличения возраста наблюдается некоторое возрастание величин отношений, но значения показателей остаются существенно меньше таковых в нормальных насаждениях (см. табл. 5).
Таблица 5. Сравнение таксационных характеристик деревьев подпологовой ели и деревьев ели, приведенных в таблицах хода роста
|
Показатель |
Возрастная |
Отношение, %, к значениям в таблицах хода роста* в возрасте ели, лет |
||||
|
группа, лет |
10 |
20 |
30 |
40 |
50 |
|
|
Средняя высота (H) |
31–40 |
33/55 |
37/77 |
44/103 |
- |
- |
|
41–50 |
33/55 |
39/82 |
45/107 |
47/113 |
- |
|
|
51–60 |
47/76 |
51/107 |
53/126 |
56/133 |
62/142 |
|
|
Площадь поперечного сечения ствола на высоте 1,3 м (G) |
31–40 |
- |
3/- |
26/101 |
- |
- |
|
41–50 |
- |
4/- |
25/100 |
28/111 |
- |
|
|
51–60 |
- |
17/- |
40/152 |
47/188 |
51/213 |
|
|
Средний объем ствола (V) |
31–40 |
4/- |
8/27 |
13/68 |
- |
- |
|
41–50 |
5/- |
10/32 |
15/78 |
17/108 |
- |
|
|
51–60 |
16/- |
27/87 |
27/137 |
26/169 |
31/224 |
|
|
Коэффициент формы ствола (q2) |
41–50 |
- |
- |
115/85 |
105/89 |
- |
|
51–60 |
- |
- |
100/74 |
95/80 |
96/84 |
|
* Числитель – по общим таблицам хода роста нормальных еловых насаждений II класса бонитета [10], знаменатель – по таблицам хода роста полных двухъярусных березово-еловых древостоев I класса бонитета с последующим возобновлением ели для подзоны средней тайги [2].
Полные двухъярусные березово-еловые насаждения уже в 30-летнем возрасте по большинству показателей, кроме среднего объема ствола в двух поздних генерациях, уступают рассматриваемым насаждениям. К 50-летнему возрасту значения показателей деревьев ели ранней генерации (51–60 лет) в 1,4–2,2 раза выше, чем деревьев ели в таблице хода роста двухъярусных березово-еловых насаждений (см. табл. 5).
Анализ формы стволов по значению коэффициента формы q2 показал, что в рассматриваемом древостое сбежистость стволов незначительно отличается от стволов в нормальных еловых древостоях, но существенно больше, чем в среднетаёжных березово-еловых древостоях с последующим возобновлением ели.
Выводы
В ходе проведенных исследований установлено следующее.
При позднем возобновлении ели, когда основная часть популяции появляется в березняках возрастом более 30 лет, немногочисленные возрастные генерации ели с периодом возобновления менее 30 лет не оказывают значительного влияния на рост деревьев последующей генерации.
В 70-летних березняках генерация ели в возрасте 41–60 лет с сомкнутостью крон 0,38 не влияет существенно на рост деревьев наиболее представительной генерации в возрасте 31–40 лет. Среднепериодический прирост деревьев в высоту, по диаметру на высоте 0,1 м и объему ствола у этих возрастных генераций статистически недостоверны. Деревья более поздних возрастных генераций, испытывая сильную внутривидовую конкуренцию, значительно отстают в росте от деревьев ранних генераций.
В течение первых 50 лет роста и развития все возрастные генерации в подпологовой популяции ели по значениям основных таксационных характеристик существенно уступают нормальным насаждениям ели, но превосходят полные двухъярусные березово-еловые с последующим возобновлением ели.
Для адекватного отображения роста и развития подпологовой популяции ели в березняках южной тайги необходимо разработать модель формирования таких насаждений.
Список литературы Возрастной аспект в исследовании роста ели под пологом березняков подзоны южной тайги европейской части России
- Писаренко, А.И. Лесовосстановление / А.И. Писаренко. - М.: Лесная промышленность, 1977. - 256 с.
- Лесотаксационный справочник для северо-востока европейской части СССР. - Архангельск: Архангельский институт леса и лесохимии, 1986. - 356 с.
- Рубцов, М.В. Закономерности роста ели под пологом березняков в онтоценогенезе древостоев / М.В. Рубцов, А.А. Дерюгин // Лесоведение. - 2002. - № 5. - С. 18-25.
- Рубцов, М.В. Рост ели под пологом южно-таежных березняков и после рубки их с сохранением подроста / М.В. Рубцов, А.А. Дерюгин // Лесной журнал. - 2007. - № 2. - С. 19-27.
- Дерюгин, А.А. Особенности роста популяции ели под пологом березняков в южной тайге Русской равнины [Электронный ресурс] / А.А. Дерюгин // Лесохозяйственная информация. - 2018. - № 1. - С. 21-30. - Режим доступа: URL:http:// lhi.vniilm.ru
- Дерюгин, А.А. Строение и рост предварительной генерации ели различной густоты после рубки березы в южной тайге европейской части России / А.А. Дерюгин, Н.А. Рыбакова // Вестник Поволжского государственного технологического университета. Сер.: Лес. Экология, Природопользование. - 2019. - № 3 (43). - С. 46-54.
- Рубцов, М.В. Возрастная динамика морфоструктуры и состояния популяции ели под пологом южно-таежных березняков / М.В. Рубцов, А.А. Дерюгин // Комплексные стационарные исследования в лесах южной тайги (Памяти М.В. Рубцова). - М.: Тов-во научных изданий КМК, 2017. - С. 125-153.
- Алексеев, В.И. Возобновление ели на вырубках / В.И. Алексеев. - М.: Наука, 1978. - 130 с.
- Анучин, Н.П. Лесная таксация / Н.П. Анучин. - М.: Лесная промышленность, 1977. - 511 с.
- Общесоюзные нормативы для таксации лесов: справочник. - М.: Колос, 1992. - 495 с.
