Встречаемость и разнообразие морфологических аномалий популяций озерной лягушки (Anura, Amphibia) Среднего Поволжья
 Среднего Поволжья")
Автор: Файзулин Александ Ильдусович
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Наземные экосистемы
Статья в выпуске: 5-1 т.14, 2012 года.
Бесплатный доступ
На территории Среднего Поволжья в 4 популяциях озерной лягушки отмечены 10 типов морфологических аномалий внешнего строения. Нами проведена оценка разнообразия аномалий по параметрам фенотипического разнообразия μ и h. Установлено, возрастание в 1,65 раза, показателя разнообразия спектра аномалий (µ±Sµ) в условиях высокого антропогенного воздействия (2,50±0,021), по сравнению с контролем (1,49±0,004).
Аномалии, среднее поволжье, биоиндикация
Короткий адрес: https://sciup.org/148201436
IDR: 148201436 | УДК: 591:
Occurrence and morphological anomalies varietyof populations of marsh frog (Anura, Amphibia) of the Middle Volga
In the Middle Volga region in 4 lake frog populations are marked morphological abnormalities of 10 types of external structure: polydactyly, polimeliya, ectromelia, ectrodactyly, absence of eyelids, eyes, the aberrations of pigmentation of the iris. We assessed abnormalities diversity in phenotypic diversity parameters μ and h. 1.65 times-increase in diversity index of anomalies spectrum (μ ± Sμ) in high antropopress (2,50 ± 0,021) compared to the controls (1,49 ± 0,004) was found.
Текст научной статьи Встречаемость и разнообразие морфологических аномалий популяций озерной лягушки (Anura, Amphibia) Среднего Поволжья
В озникновение аномалий у земноводных связано с многими независимыми и взаимодействующими факторами. Отклонения в строении вызывают мутации и взаимодействия генов, химические тератогены, а также повреждения хищников и ме-тацеркарий трематод, вызывающих аномальные регенерации конечностей [19, 21] и нарушения морфогенеза позвоночника [4]. В условиях высокой антропогенной нагрузки отмечается повышение разнообразия и общей частоты аберраций у амфибий [2, 3, 8, 16, 20, 22]. В настоящее время предлагается использовать встречаемость и разнообразие аномалий для оценки состояния популяций земноводных [2, 10, 15].
В Европейской части России морфологические отклонения в строении (кладок, личинок, неполовозрелых и половозрелых особей) отмечены у всех видов земноводных [2, 3, 7, 8, 11, 13, 14, 18]. Наибольшим разнообразием аномалий характеризуется озерная лягушка Rana ridibunda [7, 13, 14, 15, 16].
Цель нашего исследования – проанализировать состав, встречаемость и разнообразие морфологический аномалий в популяциях озерной лягушки из различных по степени антропогенного воздействия местообитаний Среднего Поволжья.
МАТЕРИАЛ И МЕТОДИКА
Для характеристики влияние антропогенной нагрузки на изменение параметров половозрастной состава в 1998 г. нами исследованы 4 популяции озерной лягушки в районе устья р. Чапаевки и Мордовенской поймы (Безенчукский и Ставропольский районы Самарской области).
Уровень антропогенной нагрузки учитывали по уровню нарушения морфогенетического гомеостаза [15, 17]. Таким образом, нами выделены для исследования следующие выборки, которые распределены с учетом возрастания антропогенной нагрузки: I. «Брусяны» – водоемы западной притеррасная часть Мордовенской поймы, изолированная от Саратовского водохранилища, 200–500 м южнее с. Брусяны (N juv. =98; N ad. =70); II. – «Мордово» – восточная центральнопойменная, занимает большую
часть поймы: Кольцовскую воложку, межгривные озера прирусловой части поймы в 200–400 м южнее пос. Мордово (N juv. =79N ad. = 21); III. – «Кольцово» – пруд оросительной системы, у восточной окраины с. Кольцово (N juv. =70N ad. = 14); IV. – «Васильевские острова» – прибрежного мельководья Васильевских о-вово, Саратовского водохранилища (N juv. =38N ad. = 27).
Для акватории у Васильевских островов в Саратовском водохранилище (2–2,5 км ниже устья р. Чапаевки) отмечены превышение ПДК (здесь и далее ПДК указаны для водоемов рыбохозяйственного назначения): в 1995–1996 гг. для легкоокисляе-мых органических веществ в 2–3 раза, фенолов в 3– 5 раз, фосфора 3–9 раз; в 1997 г. для марганца в 11 раз [12].
Кроме, собственных данных, учитывались результаты проведения исследования цитогенетического гомеостаза озерной лягушки в районе устья р. Чапаевки (Чубинишвили, 1998) и данные по интегральной оценке состояния Саратовского водохранилища по морфологическим аномалиям личинок рыб [5]. Участок акватории водохранилища в районе Васильевских островов отнесен в 1997 г. к зоне «экологического бедствия» с максимальным значением Индекса Состояния Популяции (3,24) применяемый для оценки состояния пресноводных экосистем, в то время как, у пос. Мордово, ИПС составлял в 1997 – 2,22 [5].
Статистическая оценка встречаемости по параметрам фенотипического разнообразия μ и h Л. А. Животовского [6]. Установлена, статистически достоверные различия (по критерию идентицино-сти I) между популяциями обитающими в различных условиях антропогенной нагрузки. Классификация типов аномалий проведена с учетом симметрии их проявления.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В районе исследования нами обнаружено 10 типов морфологических отклонений: полимелия, эк-тромелия, симметричная полидактилия, несимметричная полидактилия, эктродактилия, брахидакти-лия, клинодактилия, отсутствие (недоразвитие) пяточного бугра, циклопия, отсутствие зрачка. (табл. 1).
Таблица 1. Распределение морфологических аномалий в районе устья р. Чапаевки
|
Типы аномалии |
№ св Щ ев н В и В ^ & S |
Брусяны |
Мордово |
Кольцово |
Васильевские о–ва |
|
|
n/P±Sp |
n/P±Sp |
n/P±Sp |
n/P±Sp |
|||
|
Полимелия |
A/З |
juv. |
– |
– |
2/2,86±1,99 |
– |
|
Эктромелия |
A/З |
juv. |
– |
– |
– |
1/2,63±2,60 |
|
Полидактилия |
S/З |
juv. |
– |
1/1,27±1,26 |
– |
1/2,63±2,60 |
|
Полидактилия |
A/З |
juv. |
1/1,02±1,01 |
– |
– |
– |
|
Эктродактилия |
A/П |
juv. |
1/1,02±1,01 |
– |
– |
– |
|
Эктродактилия |
A/З |
ad. |
– |
– |
– |
1/3,7±3,63 |
|
Брахидактилия |
A/П |
juv. |
1/1,02±1,01 |
1/1,27±1,26 |
– |
– |
|
Брахидактилия |
A/З |
ad |
– |
– |
– |
1/3,7±3,63 |
|
Клинодактилия |
A/З |
ad |
– |
1/4,76±4,65 |
– |
– |
|
Отсутствие пяточного бугра |
A |
ad |
– |
– |
1/7,14±6,88 |
– |
|
Отсутствие глаз |
A |
juv. |
– |
– |
2/2,86±1,99 |
– |
|
Отсутствие зрачка |
A |
juv. |
– |
– |
1/1,43±1,42 |
1/2,63±2,60 |
|
Особи без аномалий |
juv. |
97/96,94±1,74 |
77/97,46±1,77 |
65/92,85±3,08 |
35/92,11±4,37 |
|
|
Особи без аномалий |
ad |
70/100±0,00 |
20/95,24±4,65 |
13/92,86±6,88 |
25/92,6±5,04 |
|
|
Всего аномалий |
juv. |
3/3,06±1,74 |
2/2,54±1,77 |
5/7,89±3,08 |
3/7,15±4,37 |
|
|
ad |
– |
1/4,76±4,65 |
1/7,4±6,88 |
2/7,14±5,04 |
||

мы Саратовского водохранилища (окр. пос. Мор-дово, Самарской области). В данном географическом пункте, симметричная полидактилия является массовой аномалий, которая отмечалась в 1997 г. (n=8; 5,93±2,03; N=135). В других регионах массовая симметричная полидактилия (c развитием дополнительного одного и двух пальцев) была зарегистрирована у серой жабы в Окском заповеднике (Рязанская обл.,) в выборке собранной в период размножения (8-9 апреля 1975; n=11) с частотой 7,2 % (N = 153) [18]. В г. Казани, отмечена только симметричная полидактилия в форме удвоения фа-
Рис. 1. Полимелия задней конечности озерной лягушки Rana ridibunda (популяция «Кольцово»)
Полимелия – развитие дополнительных конечностей. Отмечена в форме развития 1 и 2-х дополнительных конечностей (рис. 1). Для Средней Волги, отмечен 1 экз. озерной лягушки с полимелией [18]. В г. Казани, отмечена у 1 экз. озерной лягушки [8]. Данная аномалия отмечена у краснобрюхой жерлянки и зеленой жабы [13]. Одним из факторов, вызывающие полимелию (массовую) относят заражение пояса конечностей метацеркариями трематод [21]. Однако по нашим данным [14], цисты трематод в зоне развития дополнительных конечностей не обнаружены.
Эктромелия – недоразвитие конечностей. В Волжском бассейне отмечена у озерной лягушки, а также у краснобрюхой жерлянки, обыкновенной чесночницы, серой жабы [13].
Симметричная полидактилия. При рассмотрении проявления аберраций билатеральных признаков, симметричные нарушения доминируют при развитии добавочных пальцев — развитие 5 дополнительного пальца на передних, 6 – 7 на задних конечностях. Отмечен 1 случай симметричной полидактилии с развитием дополнительных пальцев на всех конечностях. В популяции озерной лягушки, обитающей на территории Мордовенской пой- ланг первого пальца, дополнительного палец между первым и вторым пальцами, сочетанием первого и второго вариантов [8]. Несимметричная полидактилия встречена у краснобрюхой жерлянки (n=2; 1,68±1,18 %), травяной (n=1; 0,68±0,68 %), остромордой (n=1; 0,70±0,70 %) и озерной (n=10; 0,59±0,18 %) лягушек.
Несимметричная полидактилия. В период исследования отмечена только в районе контроля – популяции «Брусяны». В 1996–1997 гг. регистрировались в пойменных водоемах Мордовенской поймы. Несимметричная полидактилия (развитие 6-го пальца на правой ноге), отмечена из порядка 4 тыс. просмотренных экземпляров в регионе г. Казани. В том же районе отмечены 3 экз. озерной лягушки с несимметричной полидактилией [18]. В Окском заповеднике 2 экз. с полидактилией, обнаружены в выборке из почти 10 тыс. особей прудовой (съедобной) Rana lessonae/esculenta лягушек [18].
Эктродактилия – недоразвитие пальцев. У озерной лягушки отмечена на передних в популяции «Брусяны», так и на задних конечностях, в популяции «Васильевские острова». У других видов отмечалась для передних конечностей обыкновенной чесночницы (n=1; 0,94±0,94 %) и зеленой жабы (n=1; 1,15±1,14 %) [13]. Для задних конечностей остромордой (n=1; 0,70±0,70) и травяной (n=1;
0,68±0,68) лягушек. Эктродактилия в г. Казани отмечена у 2 экз. зеленой жабы (2 экз.) и остромордой лягушки (1 экз.) [8].
Брахидактилия – укороченная длина пальцы. Отмечена во всех популяциях, кроме «Кольцово». У других видов брахидактилия несимметричная задних конечностей отмечена у серой жабы (n=1; 0,91±0,90), травяной (n=1; 0,68±0,68) и прудовой (n=1; 0,40±0,40) лягушек [13].
Клинодактилия – искривленные пальцы. Редкая аномалия, отмечена единично в популяции «Мор-дово». В Волжском бассейне отмечена у травяной (n=1; 0,68±0,68) и прудовой (n=1; 0,40±0,40) лягушек [13].
Отсутствие (недоразвитие) пяточного бугра. Отмечены только у озерной лягушки в популяции «Кольцово».
Циклопия – отсутствие глаз; Отмечена у озерной лягушки (n=2; 0,12±0,08). В г. Казани обнаружена у одной особи прудовой лягушки [8]. Аномалии – симметричное отсутствие век (n=1; 0,06±0,06 %) и несимметричное глаз (недоразвитие), отмечены только у метаморфизирующих сеголетков.
Отсутствие зрачка.Редкая аномалия, отмеченная в Волжском бассейне только у озерной лягушки [13].
Из таблицы видно, что наиболее высокая частота аномалий отмечена в популяции «Васильевские острова».словиях высокой антропогенной нагрузки. Также высокий уровень встречаемости аберраций, около 7%, наблюдается в условиях средней антропопрессии в выборке из пруда у с. Кольцово.
Различия по частоте встречаемости аномалий не являются статистически достоверными. Распределение общей частоты встречаемости аберраций и изменения величины флуктуирующей асимметрии заметно отличаются, что не позволяет оценивать состояние популяции по данному параметру. Но общая тенденция – «повышения встречаемости аномалий с ростом антропогенной нагрузки», отмеченная другими исследователями [4, 20] сохраняется. Однако, на отсутствие связи между показателем «общая частота аномалий» и нарушением стабильности развития указывает Е. Е. Коваленко [9].
Другой, более объективный показатель – разнообразие аномалий – анализируется в лабораторных [9] и природных популяциях [3]. В качестве критерия нами выбран показатель фенотипического разнообразия μ и доля редких фенотипов h [6]. Обычно, в популяции в качестве нормального фенотипа, рассматривается доминирующий тип строения, без видимых отклонений (93 – 99%) (p1). Остальные особи с видимыми отклонениями, возникающими на эмбриональной и личиночной стадии развития, – морфологическими аномалиями включены в группы типов с ненормальным строением (p2 + ...pm-1), где m – число вариантов фенотипа, включая и особей без отклонений (анализируется весь ряд фенотипов, а не только аномальные). Таким образом, данные показатели оценивают одновременно частоту встречаемости и разнообразие аномалий в выборке.
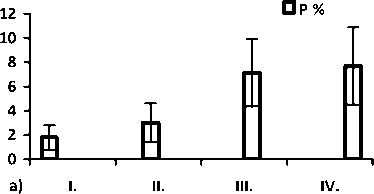
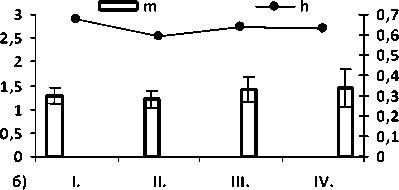
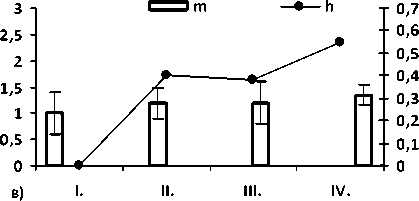
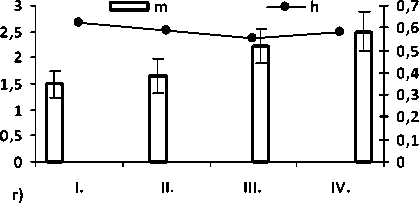
Рис. 2. Значение общей частоты аномалий по общей выборке а); величина показателей разнообразия µ и доли редких фенотипов h по морфологическим аномалиям среди: б) неполовозрелых (juv.), в) половозрелых особей (ad.) и г) по общей выборке (juv. + ad.). Цифрами обозначены популяции: I – Брусяны (контроль); II – Мордово; III – Кольцово; IV –Васильевские острова.
Значение параметров разнообразия и доли редких типов аномалий в районе исследования представлено на рис. 1. Из графика видно, что в условиях высокой антропопрессии выше разнообразие аномалий, чем в условиях средней и низкой антро-попрессии.
Среди сеголетков высоко разнообразие уродств в пруду в окр. с. Кольцово, где личиночное развитие проходит в «экстремальных» условиях – высокой плотности (водоемом сильно пересыхает к началу метаморфоза, уровень воды падает на 0,5 – 0,8 м) и резких перепадах температуры, в отличие от пойменных более крупных водоемов, где водохранилище и растительность формируют благоприятные микроклиматические условия. Достаточно редкая встречаемость особей с аномалиями, среди прошедших метаморфоз особей связана с низкой жизнеспособностью особей [21].
Таблица 2. Оценка сходства ( r ) и величина критерия идентичности (I) по спектру морфологических аномалий в районе исследования
|
Популяции |
I – критерий идентичности |
|||||
|
Брусяны |
Мордово |
Кольцово |
Васильевские о-ва |
|||
|
1 у |
Брусяны |
Juv |
– |
2,92 |
9,53* (9,49) |
6,51 |
|
Ad. |
1,58 |
1,20 |
1,88 |
|||
|
Juv.+Ad. |
5,39 |
10,17 |
6,91 |
|||
|
Мордово |
Juv |
0,983±0,010 |
– |
2,98 |
5,30 |
|
|
Ad. |
0,976±0,024 |
2,72 |
9,49* |
|||
|
Juv.+Ad. |
0,984±0,009 |
9,34 |
3,91 |
|||
|
Кольцово |
Juv |
0,945±0,020 |
0,966±0,011 |
– |
5,59 |
|
|
Ad. |
0,964±0,035 |
0,940±0,031 |
2,68 |
|||
|
Juv.+Ad. |
0,955±0,012 |
0,949±0,016 |
8,91 |
|||
|
Васильевские о-ва |
Juv |
0,949±0,024 |
0,951±0,019 |
0,944±0,023 |
– |
|
|
Ad. |
0,962±0,030 |
0,939±0,039 |
0,927±0,043 |
|||
|
Juv.+Ad. |
0,962±0,012 |
0,971±0,012 |
0,939±0,020 |
|||
Примечание. * различия достоверны на статистически значимом уровне (Р<0,05).
Таким образом, установлена зависимость разнообразия аномалий (µ) от степени антропогенного воздействия на популяцию. Наибольшие различия по показателям разнообразия (µ) между популяциями из контроля и зоны наибольшего антропогенного воздействия различаются в 1,7 раз. Подобный уровень различий (в 1,5 – 2 раза) позволяет оценить показатель разнообразия аномалий, как «хороший биоиндикатор» (4 балла) по шкале предложенной О. В. Бухариным и соавторами [1].
Полученные нами данные согласуются с оценкой состояния популяции рыб по аберрациям личинок – интегральным показателем состояния популяции (ИПС) в районе исследования (Евланов и др., 1999): от 0,21 в 1996 г. и 2,22 в 1997 г. (окр. пос. Мордово) до 3,24 в 1997 г. (в районе устья р. Чапаевки – «Васильевские острова»). По опубликованным данным тенденция увеличения разнообразия типов аномалий с повышением антропогенной нагрузки отмечается для городских территорий Екатеринбурга [2], г. Пензы [7], г. Казани [8], и г. Челябинска [16], а также в промышленных районах восточной Украины [20].
Сравнение спектра аберраций разнообразия представлен в табл. 2. Из таблицы видно, что статистически значимые различия установлены между неполовозрелыми особями популяций Кольцово и Брусяны, а также половозрелыми популяций Мор-дово и Васильевских островов.
Высокое сходство по спектру аномалий отмечается для популяций «Мордово» с популяцией «Васильевских о-вов» (r > 0,97) и «Брусяны» (r > 0,98). Ниже сходство (r < 0,96) популяции «Брусяны» с «Кольцово» и «Васильевскими о-вами». Наименьшее сходство (r < 0,95) отмечается для популяции «Кольцово» с «Мордово» и «Васильевскими о-вами». Вероятно, на спектр аномалий, кроме антропогенного воздействия влияют и условия обитания. Так популяция «Кольцово» населяющий небольшой пруд, имеет наибольшие отличия от остальных популяция, с критерием идентичности (I>8,9).
Исследования поддержаны Грантом РФФИ (проект № 12-04-31774).
Список литературы Встречаемость и разнообразие морфологических аномалий популяций озерной лягушки (Anura, Amphibia) Среднего Поволжья
- Бухарин О.В., Захаров В.М., Зинченко Т.Д., Немцева Н.В., Розенберг Г.С., Шитиков В.К. Методы биомониторинга для оценки состояния антропогенно-нагруженной равнинной реки//Экология и пром-сть России. 2010, №11. С. 10-15.
- Вершинин В.Л. Экологические особенности популяций амфибий урбанизированных территорий. Автореф. дис. … д-ра биол. наук. Екатеринбург, 1997. 47 с.
- Вершинин В.Л. Морфологические аномалии амфибий городской черты//Экология. 1989. № 3. С. 58-66.
- Вершинин В.Л., Неустроева Н.С. Роль трематодной инвазии в специфике морфогенеза скелета бесхвостых амфибий на примере Rana arvalis Nilsson, 1842//Докл. Академии наук. 2011. Т. 440. № 2. С. 279-281.
- Евланов И.А., Минеев А.К., Розенберг Г.С. Оценка состояния пресноводных экосистем по морфологическим аномалиям у личинок рыб: методическое пособие. Тольятти: ИЭВБ РАН, 1999. 38 с.
- Животовский Л.А. Показатели популяционной изменчивости по полиморфным признакам//Фенетика популяций. М.: Наука, 1982. С. 38-44.
- Закс М.М. О морфологических аномалиях зеленых лягушек (Rana ridibunda, R. lessonae) г. Пензы//Изв. ПГПУ им. В.Г. Белинского. 2008. № 10 (14). С. 63-65.
- Замалетдинов Р.И. Морфологические аномалии в городских популяциях бесхвостых амфибий (на примере г.Казани)//Совр. герпетология. 2003, Т. 2. C. 148-153.
- Коваленко Е.Е. Методологические проблемы биоиндикации//Вопр. герпетологии. Пущино-Москва, 2001. С. 124-126.
- Леонтьева О.А., Семёнов Д.В. Земноводные как биоиндикаторы антропогенных изменений среды//Успехи современной биологии. 1997. Т. 117. № 6. С. 726-736.
- Литвинов Н.А., Файзулин А.И., Шураков А.И., Ганщук С.В. Анализ состояния кладок сибирского углозуба Salamandrella keyserlingii Dybowski, 1870 (Caudata, Amphibia) Предуралья//Поволж. экологич. журн. 2010. № 4. С. 438-441.
- Селезнев В.А., Цыкало В.А., Сергиенко Т.С. Содержание марганца в поверхностных водах Самарской области//10 лет Гос. комитету по охране окружающей среды Самарской области: Итоги научных исследований. Самара: Комитет по охране окружающей среды Самарской области, 1998. С. 108-117.
- Файзулин А.И. О морфологических аномалиях бесхвостых земноводных (Anura, Amphibia) Волжского бассейна//Працi Українського герпетологiчного товариства. 2011, N. 3. С. 201-207.
- Файзулин А.И., Чихляев И.В. Морфологические аномалии бесхвостых земноводных (Anura, Amphibia) Среднего Поволжья//Актуальные проблемы герпетологии и токсинологии (Сборник научных трудов) Вып. 9. Тольятти, 2006. С. 178-182.
- Файзулин А.И. Эколого-фаунистический анализ земноводных Среднего Поволжья и проблемы их охраны. Автореферат диссертации на соискание ученой степени кандидата биологических наук. Тольятти: ИЭВБ, 2004. 20 с.
- Чибилёв Е.А. Биология и экология зеленых и бурых лягушек Челябинской городской агломерации//Животные в антропогенном ландшафте: Материалы 1 Международной научно-практической конференции. Астрахань: Издательство Астраханского государственного университета, 2003. С. 73-76.
- Чубинишвили А.Т. Гомеостаз развития в популяциях озерной лягушки (Rana ridibunda Pall.), обитающих в условиях химического загрязнения в районе Средней Волги//Экология. 1998. № 1. С. 71-74.
- Borkin L.J., Pikulik M.M. The occurrence of polymely and polydactyly in natural populations of Anurans of the USSR//Amphibia-Reptilia. Vol. 7, № 3. 1986. P. 205-216.
- Dubois A. Anomalies and mutations in natusal populations of the Rana “esculenta” complex (Amphibia, Anura)//Mitt. Zool. Mus. Berlin. 1979. Bd. 55, H. 1. S. 59-87.
- Flax N.L., Borkin L.J. High of incidence in anurans in contaminated industrial areas (eastern Ukraine)//Herpetologia bonnensis. 1997. P. 119-123.
- Guex G.-D., Hotz H., Uzzell T., Semlitsch R. D., Beerli P., Pascolini R. Developmental disturbances in Rana esculenta tadpoles and metamorphs//Mitt. Zool. Mus. Berlin., 2001 Bd. 77, H. 1. S. 79-86.
- Machado C.,A., Schlüter K.A. Polydactyly and polymely in two populations of Rana temporaria and Pelophylax esculentus (Anura, Ranidae) in southern Germany.//Salamandra, 2010. V. 46(4). P. 239-242.
