Выделение и характеристика штамма - деструктора дибутилфталата Rhodococcus sp. 5a-k4

Автор: Пьянкова А.А., Краева А.В., Нечаева Ю.И., Плотникова Е.Г.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Микробиология
Статья в выпуске: 3, 2024 года.
Бесплатный доступ
Изучена способность бактериального штамма 5A-K4, выделенного из ризосферы растений бескильницы расставленной (Puccinellia distans (Jacq.) Parl.), произрастающих на территории промышленных разработок Верхнекамского месторождения солей (Пермский край), к росту на дибутилфталате (ДБФ) в качестве единственного источника углерода и энергии. Штамм 5A-K4 идентифицирован как представитель рода Rhodococcus. Анализ гена 16 S рРНК исследуемого штамма показал наибольший уровень сходства (99.86%) с гомологичным геном R. erythropolis NBRC 15567T. Штамм эффективно рос на ДБФ в качестве субстрата в присутствии до 30 г/л NaCl в среде культивирования. Показано, что Rhodococcus sp. 5A-K4 способен к росту на высоких концентрациях ДБФ (до 12 г/л). В геноме штамма выявлены гены dpeH и mpeH, продукты которых участвуют на начальных этапах разложения ДБФ. Нуклеотидные последовательности генов dpeH и mpeH гомологичны последовательностям генов а/p гидролаз штаммов - представителей классов Actinomycetes и Bacilli. На основании полученных данных предложен путь разложения ДБФ штаммом 5A-K4. Таким образом, ризосферный штамм-деструктор Rhodococcus sp. 5A-K4 может быть использован в качестве бактериального агента при разработке методов фиторемедиации почв, загрязненных фталатами.
Бескильница расставленная, ризосфера, дибутилфталат, деструкция, rhodococcus
Короткий адрес: https://sciup.org/147246121
IDR: 147246121 | УДК: 579.26: | DOI: 10.17072/1994-9952-2024-3-309-317
Isolation and с haracterization of dibutyl phthalate-degrading strain Rhodococcus sp. 5a-k4
The study is devoted to the ability of the bacterial strain 5A-K4, isolated from the rhizosphere of Puccinellia distans (Jacq.) Parl. plants, growing on the territory of industrial development of the Verkhnekamsk salt deposit (Perm krai), to grow on dibutyl phthalate (DBP) as the single carbon source was studied. Based on analysis of the 16S rRNA gene, the strain was identified as a representative of the genus Rhodococcus. Strain 5A-K4 had the highest level of the 16S rRNA gene similarity (99.86%) with Rhodococcus erythropolis NBRC 15567T. The strain grew effectively on a DBP substrate without addition of salt and at a content of 30 g/L NaCl in the cultivation medium. It has been shown that Rhodococcus sp. 5A-K4 is capable of growth at high concentrations of DBP (up to 12 g/L). The genome of the strain 5A-K4 contained the dpeH and mpeH genes, the products of which are involved in the initial stages of DBP degradation. The nucleotide sequences of the dpeH and mpeH genes are homologous to the sequences of the а/p hydrolase genes of strains of the classes Actinomycetes and Bacilli. Based on the data obtained, a pathway for the degradation of DBP by strain 5A-K4 was proposed. Thus, the rhizosphere DBP degrading strain Rhodococcus sp. 5A-K4 can be used as a bacterial agent in the development of phytoremediation methods for soils contaminated with phthalates.
Текст научной статьи Выделение и характеристика штамма - деструктора дибутилфталата Rhodococcus sp. 5a-k4
Эфиры фталевой кислоты (ЭФК) – класс токсичных антропогенных соединений, которые преимущественно использовались в качестве добавок или пластификаторов для увеличения гибкости, прозрачности и долговечности пластиковых материалов [Lu М. et al., 2020]. Дибутилфталат (ДБФ) является одним из наиболее широко используемых ЭФК, применяется при производстве инсектицидов, упаковочных материалов, косметики, покрытий, одежды и изоляторов для электрооборудования [He, Niu, Lu, 2014]. ЭФК обладают потенциальной канцерогенностью, тератогенностью и мутагенностью [Gardner et al., 2016], в связи с чем, внесены Агентством по охране окружающей среды США, Европейским Союзом и Национальным центром экологического мониторинга Китая в список приоритетных загрязнителей [Lu М. et al., 2020].
В настоящее время ЭФК обнаружены во всем мире в различных средах, таких как вода, воздух, ил, почва и донные отложения [Gao, Wen, 2016]. Значительные концентрации фталатов выявлены в районах работы предприятий горнодобывающей промышленности в составе глинисто-солевых шламов, избыточных рассолов и отходов калийного производства, вследствие применения реагентов, продуктами трансформации которых являются фталаты [Бачурин, Одинцова, 2006].
Одним из основных путей деградации ЭФК в окружающей среде считается разложение с помощью микроорганизмов. Биодеградация ЭФК является эффективным, недорогим, безопасным и экологически чистым методом [Gao, Wen, 2016; Lu М. et al., 2020]. Среди штаммов, способных осуществлять деструкцию ДБФ, описаны бактерии рода Rhodococcus [Aleshchenkova et al., 1996; Li, Gu, 2005; Li et al., 2006; Chao, Cheng, 2007; Lu Y. et al., 2009; Jin et al., 2010; He, Niu, Lu, 2014; Yang et al., 2018; Wang et al., 2022]. Путь деградации эфиров фталевой кислоты у большинства аэробных бактерий, в том числе у представителей рода Rhodococcus , аналогичен и включает гидролиз диэфиров фталата до моноэфиров и последующую их деструкцию до орто -фталевой кислоты и бутанола [Chao et al., 2006; Wang et al., 2022]. Дальнейший путь разложения осуществляется через орто -фталевую кислоту, которая метаболизируется через образование протокатеховой кислоты и, далее – до основных продуктов жизнедеятельности микробной клетки [Vamsee-Krishna, Phale, 2008; Gao, Wen, 2016]. Однако, несмотря на растущее количество сообщений о штаммах, разлагающих ЭФК, генах и ферментах, связанных с деградацией, механизм гидролиза ЭФК требует дальнейшего изучения. Исследование механизмов деградации ЭФК, а также поиск эффективных штаммов-деструкторов будет способствовать разработке новых подходов к восстановлению окружающей среды.
Цель работы – характеристика штамма-деструктора дибутилфталата, выделенного из ризосферы растений бескильницы расставленной, произрастающих в районе промышленных разработок Верхнекамского месторождения солей (Пермский край).
Материалы и методы
Объекты исследования. В работе использован штамм 5A-K4, выделенный из ризосферы растений бескильницы расставленной (Puccinellia distans (Jacq.) Parl.), произрастающих на территории Верхнекамского промышленного региона (Пермский край). Образцы растений были отобраны на расстоянии 8 м от солеотвала (г. Соликамск, Пермский край, Россия), координаты участка отбора проб: N59.633686°, E56.760206°. Почва исследуемого участка супесчаная подзолистая, отличается повышенным содержанием хлоридов, сульфатов, бикарбонатов. Содержание ионов Na+, К+, Ca2+ составляло 17, 26, 88 мг/кг, соот- ветственно. В исследуемой почве выявлено наличие дибутилфталата/диоктилфталата в концентрации 1.0/1.4 мг/кг [Nazarov et al., 2024].
Среды и условия культивирования . Для культивирования бактерий использовали минеральную среду Раймонда ( МСР ) [Raymond, 1961]. В качестве субстратов использовали дибутилфталат (ДБФ), диметилфталат, диэтилфталат, орто- фталевую, протокатеховую, бензойную кислоты, нафталин, бифенил и бутанол в концентрации 1.0 г/л, а также салициловую и гентизиновую кислоты в концентрации 0.5 г/л. Для приготовления богатой среды Раймонда ( БСР ) в МСР добавляли 5 г/л триптона («VWR Life Science Amresco», США) и 2.5 г/л дрожжевого экстракта («Biospringer», Франция) в качестве ростовых субстратов. В работе использовали среды без добавления соли, а также среды с концентрацией NaCl до 70 г/л. Для приготовления плотных сред вносили агар («Helicon», Россия) до конечной концентрации 15 г/л.
Накопительное культивирование и выделение бактерий . 5 г образца ризосферной почвы ресус-пендировали в 10 мл МСР, затем 100 мкл полученной суспензии помещали в 15 мл МСР (30 г/л NaCl) с ДБФ в качестве единственного источника углерода и энергии (1 г/л) в колбу объемом 50 мл. Культивирование проводили на термостатируемом шейкере Environmental Shaker Incubator ES-20/60 («BioSan», Латвия) при 100 об./мин. в течение 3 месяцев, после чего осуществляли высев суспензии на агаризован-ную БСР с содержанием 30 г/л NaCl и инкубировали при 28°С до появления колоний.
Описание морфологии колоний бактерий проводили на седьмой день культивирования на агаризо-ванной БСР (30 г/л NaCl).
Выделение ДНК из чистой культуры бактерий проводили общепринятым методом [Ausbel et al., 1995].
Амплификацию гена 16S рРНК проводили с универсальными бактериальными праймерами 27F и 1492R [Lane, 1991] на амплификаторе C1000 TouchTM Thermal Cycler («Bio-Rad Laboratories», США).
Исследование генов деструкции ДБФ осуществляли путем амплификации фрагментов генов dpeH и mpeH , кодирующих дибутил- и монобутилэстеразу, соответственно, с последующим секвенированием и анализом полученных нуклеотидных последовательностей. Амплификацию генов dpeH и mpeH осуществляли с использовалнием праймеров: dpeH-F 5’-TAAGAAGGAGATATACA TATGCAGATCGTACTGGTGCACG-3’, dpeH-R 5’-GTGGTGGTGGTGGTGCTCGAGGGCAATGACGG GCGTCGAG-3’ и mpeH-F 5’-TAAGAAGGAGATATACATATGGATTCGATTTCACGCCC-3’, mpeH-R 5’-GTGGTGGTGGTGGTGCTCGAGGAGCGCCGTGATCCACCG-3’, соответственно [Lu et al., 2020].
Для детекции ПЦР-продуктов использовали горизонтальный электрофорез в 1%-ном агарозном геле с использованием 1× буфера ТВЕ (Трис – 10.8 г/л, борная кислота – 5.5 г/л, 0.5 М ЭДТА – 4 мл, вода дистиллированная – 79.7 мл/л) при комнатной температуре, напряжении 5–15 В/см в течение 20–40 мин. Агарозные гели окрашивали раствором бромистого этидия (0.5 мкг/мл) в течение 15 мин. и фотографировали в УФ-свете с помощью системы гельдокументирования BioDocAnalyze («Bio-Rad Laboratories», США). Для определения размеров полученных фрагментов использовали маркер длин ДНК 100+ bp DNA Ladder («Евроген», Россия).
Секвенирование и анализ генов. Определение нуклеотидных последовательностей генов 16S рРНК, dpeH и mpeH проводили с применением набора реактивов «GenSeq-100» («Синтол», Россия) на автоматическом секвенаторе Нанофор 05 («Синтол», Россия) согласно рекомендациям производителя. Анализ полученных последовательностей осуществляли с использованием программ Sequence Scanner v. 2.0, MEGA v. 7.0 . Поиск гомологичных последовательностей осуществляли по международным базам данных EzBioCloud и GenBank (http:/. Для построения филогенетических деревьев использовали метод «neighbor-joining» программы MEGA v. 7.0. Эволюционные расстояния рассчитывали с использованием метода «p-distance». Статистическую достоверность ветвления («bootstrap»-анализ) оценивали на основе 1 000 альтернативных деревьев (приведены значения выше 50%).
Рост бактерий на ДБФ оценивали при культивировании в жидкой МСР. Субстрат вносили до конечной концентрации 1.0, 3.0, 5.0, 7.0, 9.0, 10.0, 11.0, 12.0 г/л. В качестве инокулята добавляли культуру (ОП 600 =1.0), выращенную в жидкой МСР с ДБФ в концентрации 1.0 г/л. Культивирование осуществляли в колбах объемом 250 мл (объем среды – 100 мл) на термостатируемом шейкере Environmental Shaker Incubator ES-20/60 («BioSan», Латвия) при температуре 28°С и скорости вращения 140 об/мин. Оптическую плотность ( ОП ) культуральной жидкости определяли на спектрофотометре
UV-Visible BioSpec-mini («Shimadzu», Япония) при длине волны 600 нм в кювете с длиной оптического пути 1 см. Удельную скорость роста ( μ , ч-1) рассчитывали по стандартной формуле
μ = (lnB2 – lnB1)/(t2 – t1), где В1 и В2 – оптические плотности культуры в моменты времени t1 и t2, соответственно [Нетрусов, 2005].
Статистическая обработка результатов . Все эксперименты были выполнены в трехкратной повторности. Для обработки полученных данных использовали программу Microsoft Excel 2007.
Результаты и их обсуждение
Методом накопительного культивирования на МСР с 30 г/л NaCl и ДБФ в качестве субстрата из ризосферы растений бескильницы расставленной, отобранных с территории солеразработок (г. Соликамск, Пермский край), был изолирован бактериальный штамм 5A-K4.
При культивировании на богатой среде (БСР) штамм имел бежевые округлые непрозрачные колонии диаметром 2–3 мм с ровным краем, гладкой блестящей поверхностью, выпуклым профилем, мелкозернистой структурой и мягкой консистенцией. На основе анализа фрагмента гена 16S рРНК (735 п.н.) штамм 5A-K4 имел наибольшее сходство, на уровне 99.86%, со штаммом Rhodococcus erythropolis NBRC 15567T.
При культивировании на БСР при различных концентрациях NaCl в среде, показано, что штамм растет как на среде без добавления соли, так и при содержании NaCl до 50 г/л.
Поскольку штамм был выделен из накопительной культуры с ДБФ в качестве субстрата, была проверена его способность к росту на фталатах и на возможных продуктах их разложения ( орто -фталевой, протокатеховой кислотах и бутаноле). Установлено, что штамм способен к эффективному росту на среде с дибутилфталатом, диметилфталатом, диэтилфталатом, бутанолом и протокатеховой кислотой, но не растет на орто -фталевой кислоте. Также штамм может использовать в качестве единственного источника углерода бензойую, салициловую и гентизиновую кислоты.
Исследованы ростовые характеристики штамма при культивировании на ДБФ в качестве субстрата без соли, а также при различных концентрациях NaCl в среде (табл. 1). Наиболее высокие показатели максимальной ОП и удельной скорости штамм имел при росте на среде без соли. При культивировании в среде с добавлением 30 г/л NaCl штамм также демонстрировал эффективный рост (табл. 1). При добавлении в среду 40 г/л NaCl рост штамма на среде с ДБФ не наблюдался.
Таблица 1
Параметры роста штамма 5A-K4 в МСР с ДБФ (1.0 г/л) при разных концентрациях NaCl [Growth parameters of the strain 5A-K4 in MMR and DBP (1.0 g/L) at different concentrations of NaCl]
|
Концентрация NaCl, г/л |
Удельная скорость роста, ч-1 |
Максимальное значение ОП 600 |
|
без NaCl |
0.038±0.002 |
1.41 |
|
30 |
0.021±0.001 |
0.81 |
Так как штамм 5A-K4 демонстрировал эффективный рост на ДБФ в качестве субстрата (табл. 1), были исследованы ростовые характеристики штамма при культивировании с высокими концентрациями ДБФ в среде МСР без добавления соли. Показано, что штамм способен к росту при содержании ДБФ до 12 г/л (табл. 2). При увеличении концентрации ДБФ в среде культивирования лаг-фаза изменялась незначительно. Максимальное значение ОП зафиксировано при содержании 3 г ДБФ/л, а максимальное значение удельной скорости роста – при 1 г ДБФ/л (табл. 2).
Таблица 2
Параметры роста штамма 5A-K4 в МСР при разных концентрациях ДБФ [Growth parameters of the strain 5A-K4 in MMR at different concentrations of DBP]
|
Концентрация ДБФ, г/л |
Удельная скорость роста, ч-1 |
Максимальное значение ОП 600 |
|
1 |
0.084±0.004 |
1.13 |
|
3 |
0.060±0.003 |
1.89 |
|
5 |
0.047±0.002 |
1.76 |
|
7 |
0.042±0.003 |
1.57 |
|
9 |
0.028±0.002 |
1.38 |
|
10 |
0.016±0.002 |
1.15 |
|
11 |
0.010±0.001 |
1.02 |
|
12 |
0.007±0.001 |
0.72 |
Проведена амплификация генов dpeH и mpeH , продукты которых участвуют в двух последовательных стадиях гидролиза ДБФ [Lu et al., 2020]. С использованием праймеров, описанных [Lu M. et al., 2020], у исследуемого штамма 5A-K4 выявлено наличие обоих генов (рис. 1).
Проведены секвенирование и анализ амплифицированных генов dpeH и mpeH. Сравнение гена dpeH штамма 5A-K4 с гомологичными последовательностями из базы данных GenBank показало, что данная последовательность имеет наибольше сходство с последовательностями α/β гидролаз бактерий – представителей классов Actinomycetes и Bacilli. Сходство на уровне 99.85% сравниваемая последовательность имеет с геном α/β гидролазы штамма Rhodococcus pyridinivorans 5Ap, который был изолирован из ливийской почвы, загрязненной нефтью [Чернявская и др., 2016]. На филогенетическом дереве транслированных аминокислотных последовательностей (ТАП) данный штамм формирует отдельный кластер с исследуемым штаммом Rhodococcus sp. 5A-K4 (рис. 2). Также ген dpeH штамма 5A-K4 имеет сходство (99.54%) с гомологичными генами Bacillus subtilis DK01 и штаммами семейства Micrococcaceae (уровень сходства 99.39%) – Glutamicibacter sp. ZJUTW, Microbacterium paraoxydans 70447 и Microbacterium schleiferi A32-1 (рис. 2). Стоит отметить, что штамм Glutamicibacter sp. ZJUTW, выделенный из активного ила (Китай), использовался в составе бактериального консорциума для одновременного и эффективного разложения шести эфиров фталевой кислоты и демонстрировал наибольшую активность при деградации короткоцепочечных эфиров [Liu et al., 2023].
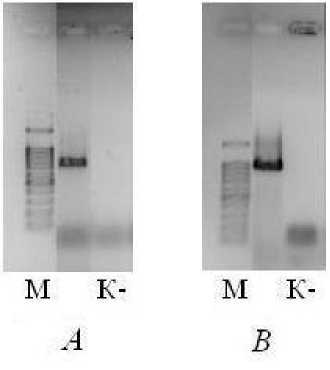
Рис. 1. Электрофореграмма результатов амплификации генов mpeH ( A ) и dpeH ( B ) штамма 5A-K4.
M – маркер 100+ bp DNA Ladder («Евроген», Россия), K- – отрицательный контроль
[Electropherogram of the mpeH ( A ) and dpeH ( B ) amplification results of the strain 5A-K4.
M – 100+ bp DNA Ladder marker (“Evrogen”, Russia), K- – negative control]
力 іЕ^ е Н ЛЕ2 ^ tf^ ^дгг^ sp. 5A4
加 I 山口 г идр 口 паз а еЛ 口曲酬比弊 用旧"由"也"期 в 口出 54 p (Q 口鹏汇 11516)
in L сф гидроп-ща Е&а 就 ін jzj-Mij DEE 1 (WSFLM643)
I 山卩 г ндриЕ.аза J 俄 Wrm5nЕ 財 Hum 戸占匕口: pti2Hls 7044*7 (QXE31721) 石 cdfl г itjijio Л-Щ.1 J й7сга& □ ctsnuyn 15aЛе 淞 г" A32-1 (QPED4173j
—сф гидр 口 паза GJumm2Msi w*r 5P. £ JUTWi : QEF09054 : i
__________I-------- 山卩 гидр 口 паза. 戍向□必比比也 sp. LW-X Y12 (С?х'Ш5б2ёГ ; і
SI------------------------------------------DpeH Micro bactenuj-гі 5P. PAE-1 (AZFty727)
Рис. 2. Положение dpeH -гена штамма Rhodococcus sp. 5A-K4 на филогенетическом дереве, построенном на основании сравнительного анализа ТАП генов α/β гидролаз.
Масштаб соответствует 5 аминокислотным заменам на каждые 1000 аминокислот. В скобках указаны номера в базе данных GenBank
[Positions of the gene dpeH of the strain Rhodococcus sp. 5A-K4 in the phylogenetic tree based on comparison of the amino acid sequences translated from α/β hydrolase genes.
The scale bar corresponds to 5 amino acid substitutions per 1000 amino acids. The GenBank accession numbers are given in parentheses]
Еще один кластер на филогенетическом дереве формируют ТАП α/β гидролаз штаммов Rhodococ-cus sp. LW-XY12 и Microbacterium sp. PAE-1. Уровень сходства нуклеотидной последовательности гена dpeH штамма 5A-K4 и последовательностей штаммов, входящих в этот кластер, составлял 98.47–98.93%. Штамм Rhodococcus sp. LW-XY12, изолированный из активного ила (Китай), способен к эффективному разложению ЭФК [Song et al., 2022]. Штамм Microbacterium sp. PAE-1, выделенный из сточных вод заво- да по производству пластмасс, способен к разложению ДБФ до орто-фталевой кислоты через монобутилфталат посредством двух последовательных стадий гидролиза с участием продуктов генов dpeH и mpeH [Lu M. et al., 2020]. Анализ полученной последовательности гена mpeH штамма 5A-K4 показал, что данная последовательность имеет наибольшее сходство с генами mpeH тех же штаммов (представителей классов Actinomycetes и Bacilli), с которыми близка и последовательность гена dpeH (рис. 2, 3). Кроме штаммов, с которыми выявлено сходство по гену dpeH, нуклеотидная последовательность гена mpeH штамма Rhodococcus sp. 5A-K4 имеет высокий процент сходства (98.97%) и с нуклеотидной последовательностью α/β гидролазы штамма Rhodococcus ruber YC-YT1, способного к эффективному разложению эфиров фталевой кислоты [Yang et al., 2018].
α/β гидролаза Rhodococcus sp. LW-XY12 (QXU56362)
α/β гидролаза Microbacterium schleiferi A32-1 (QPE04172)
-
54 MpeH Microbacterium sp. PAE-1 (AZP89726)
0.002
78 I 一 I α/β гидролаза Rhodococcus ruber YC-YT1 (AXY49591)
69 α/β гидролаза Microbacterium paraoxydans 70447 (QXE31720) α/β гидролаза Glutamicibacter sp. ZJUTW (QEP09053)
α/β гидролаза Bacillus subtilis DK01 (WGP04643)
I MpeH Rhodococcus sp. 5A-4
95 α/β гидролаза Rhodococcus pyridinivorans 5Ap (QOW01517)
Рис. 3. Положение mpeH -гена штамма Rhodococcus sp. 5A-K4 на филогенетическом дереве, построенном на основании сравнительного анализа ТАП генов α/β гидролаз.
Масштаб соответствует 2 аминокислотным заменам на каждые 1000 аминокислот. В скобках указаны номера в базе данных GenBank
[Positions of the gene mpeH of the strain Rhodococcus sp. 5A-K4 in the phylogenetic tree based on comparison of the amino acid sequences translated from α/β hydrolase genes.
The scale bar corresponds to 2 amino acid substitutions per 1000 amino acids. The GenBank accession numbers are given in parentheses]
Таким образом, у исследуемого штамма Rhodococcus sp . 5A-K4 показано наличие генов dpeH и mpeH , продукты которых участвуют в двух последовательных стадиях гидролиза ДБФ через монобутилфталат до орто- фталевой кислоты (рис. 4).
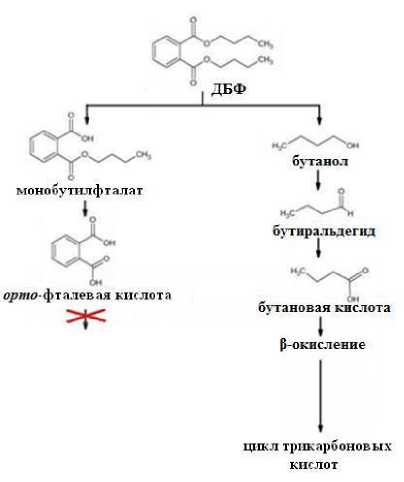
Рис. 4. Схема вероятного пути разложения ДБФ
[A scheme of probable degradation pathway of DBP]
Установлено, что штамм 5A-K4 не способен к росту на орто-фталевой кислоте – основном промежуточном метаболите разложения ДБФ у бактерий [Gao, Wen, 2016]. Было зафиксировано накопление орто-фталевой кислоты в культуральной жидкости при росте исследуемого штамма в МСР с ДБФ в качестве субстрата (данные не представлены). Поэтому можно предположить, что деструкция ДБФ у Rhodococcus sp. 5A-K4 идет по пути, представленному на рис. 4, через промежуточный продукт метаболизма ДБФ – бутанол [Mahajan et al., 2019]. Это предположение также подтверждается способностью штамма Rhodococcus sp. 5A-K4 к эффективному росту на бутаноле в качестве субстрата.
Заключение
В результате проведенных исследований из ризосферы растений бескильницы расставленной ( Puccinellia distans (Jacq.) Parl.), произрастающих на загрязненных/засоленных почвах района разработок Верхнекамского месторождения солей (Пермский край), изолирован штамм-деструктор дибутилфталата Rhodococcus sp. 5A-K4. Штамм способен к эффективному росту на среде с дибутилфталатом, а также диметил- и диэтилфталатом, бутанолом, протокатеховой, бензойной, салициловой и гентизиновой кислотами в качестве субстрата. В геноме штамма выявлены гены dpeH и mpeH , продукты которых участвуют в гидролизе ДБФ. Нуклеотидные последовательности генов dpeH и mpeH сходны с последовательностями генов α/β гидролаз бактерий – представителей классов Actinomycetes и Bacilli . Интересен факт, что штамм не растет на орто -фталевой кислоте, являющейся ключевым метаболитом при деструкции ДБФ у большинства изученных бактерий-деструкторов ЭФК. На основании полученных данных предложен путь разложения ДБФ штаммом 5A-K4 с использованием бутанола в качестве ключевого метаболита. Rhodococcus sp. 5A-K4 может использовать в качестве единственного источника углерода и энергии ДБФ при содержании 30 г/л NaCl в среде, а также способен к эффективному росту при концентрации ДБФ до 12 г/л. Таким образом, ризосферный деструктор ДБФ Rhodococcus sp. 5A-K4 представляет интерес для дальнейших исследований и возможности использования его в качестве бактериального агента при разработке методов фиторемедиации почв, загрязненных фталатами.
Список литературы Выделение и характеристика штамма - деструктора дибутилфталата Rhodococcus sp. 5a-k4
- Бачурин Б.А., Одинцова Т.А. Стойкие органические загрязнители в отходах горного производства // Современные экологические проблемы Севера. Апатиты: Изд-во Кольского НЦ РАН, 2006. Ч. 2. С. 7-9.
- Нетрусов А.И. Практикум по микробиологии. М.: Академия, 2005. 608 с.
- Чернявская М.И. и др. Первичный анализ генома бактерий-деструкторов нефти Rhodococcus pyridinivorans 5Ap // Труды БГУ. 2016. Т. 11, ч. 1. С. 219-223.
- Aleshchenkova Z.M. et al. The degradation of plasticizers by Rhodococcus erythropolis 40F // Mikrobiol. Z. 1996. Vol. 58(4). P. 34-38.
- Ausbel F.M. Short Protocols in Molecular Biology. 3rd ed. N.Y.: John Wiley & Sons, 1995. 450 p.
- Chao W.L., Cheng C.-Y. Effect of introduced phthalate-degrading bacteria on the diversity of indigenous bacterial communities during di-(2-ethylhexyl) phthalate (DEHP) degradation in a soil microcosm // Chemo-sphere. 2007. Vol. 67(3). P. 482-488.
- Chao W.L. et al. Degradation of di-butyl-phthalate by soil bacteria // Chemosphere. 2006. Vol. 63(8). P. 1377-1383.
- Gao D.-W., Wen Z.-D. Phthalate esters in the environment: a critical review of their occurrence, biodegradation, and removal during wastewater treatment processes // Science of the Total Environment. 2016. Vol. 541. P. 986-1001.
- Gardner S.T. et al. Assessing differences in toxicity and teratogenicity of three phthalates, diethyl phthalate, di-n-propyl phthalate, and di-n-butyl phthalate, using Xenopus laevis embryos // Journal of Toxicology and Environmental Health. 2016. Vol. 79(2). P. 71-82.
- He Z., Niu C., Lu Z. Individual or synchronous biodegradation of di-n-butyl phthalate and phenol by Rhodococcus ruber strain DP-2 // Journal of Hazardous Materials. 2014. Vol. 273. P. 104-109.
- Jin D.-C. et al. Biodegradation of di-n-butyl phthalate by Rhodococcus sp. JDC-11 and molecular detection of 3,4-phthalate dioxygenase gene // Journal of Microbiology and Biotechnology. 2010. Vol. 20(10) P. 1440-1445.
- Lane D.J. 16S/23S rRNA sequencing // Nucleic acid techniques in bacterial systematics. 1991. P. 115175.
- Li J. et al. Bioremediation of environmental endocrine disruptor di-n-butyl phthalate ester by Rhodococcus ruber // Chemosphere. 2006. Vol. 65(9). P. 1627-1633
- Li K., Gu J. Biodegradation of di-n-butyl phthalate by mangrove microorganism Rhodococcus ruber 1K // Ying Yong Sheng Tai Xue Bao. 2005. Vol. 16(8). P. 1566-1574.
- Liu T. et al. Synthetic bacterial consortia enhanced the degradation of mixed priority phthalate ester pollutants // Environmental Research. 2023. Vol. 235. P. 116-121.
- Lu M. et al. Degradation of dibutyl phthalate (DBP) by a bacterial consortium and characterization of two novel esterases capable of hydrolyzing PAEs sequentially // Ecotoxicology and Environmental Safety. 2020. Vol. 195. P. 1-9.
- Lu Y. et al. Biodegradation of dimethyl phthalate, diethyl phthalate and di-n-butyl phthalate by Rhodococcus sp. L4 isolated from activated sludge // Journal of Hazardous Materials. 2009. Vol. 168(2-3). P. 938-943.
- Mahajan R. et al. Biodegradation of di-n-butyl phthalate by psychrotolerant Sphingobium yanoikuyae strain P4 and protein structural analysis of carboxylesterase involved in the pathway // International Journal of Biological Macromolecules. 2019. Vol. 122. P. 806-816.
- Nazarov A.V. et al. Soil bacterial communities in the affected zone of salt dump (Solikamsk, Perm krai) // Eurasian Soil Science, 2024, Vol. 57(8). P. 1353-1361.
- Raymond R.L. Microbial oxidation of n-paraffinic hydrocarbons // Developments in Industrial Microbiology. 1961. Vol. 2(1). P. 23-32.
- Song X. et al. Biodegradation of phthalate acid esters by a versatile PAE-degrading strain Rhodococcus sp. LW-XY12 and associated genomic analysis // International Biodeterioration and Biodegradation. 2022. Vol. 170. P. 1-12.
- Vamsee-Krishna C., Phale P.S. Bacterial degradation of phthalate isomers and their esters // Indian Journal of Microbiology. 2008. Vol. 48. P. 19-34.
- Wang L. et al. Analysis of the performance of the efficient di-(2-ethylhexyl) phthalate-degrading bacterium Rhodococcus pyridinovorans DNHP-S2 and associated catabolic pathways // Chemosphere. 2022. Vol. 306. P. 135-142.
- Yang T. et al. Biodegradation of di-(2-ethylhexyl) phthalate by Rhodococcus ruber YC-YT1 in contaminated water and soil // International Journal of Environmental Research and Public Health. 2018. Vol. 15(5). P. 1-20.
