Выделение и идентификация клубеньковых бактерий гуара Cyamopsis tetragonoloba (L.) Taub
 Taub")
Автор: Кузнецова И.Г., Сазанова А.Л., Сафронова В.И., Попова Ж.П., Соколова Д.В., Тихомирова Н.Ю., Оследкин Ю.С., Карлов Д.С., Белимов А.А.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Выделение и идентификация клубеньковых бактерий
Статья в выпуске: 6 т.53, 2018 года.
Бесплатный доступ
Циамопсис четырехкрыльниковый, или гуар ( Cyamopsis tetragonoloba, сем. Fabaceae ), - однолетняя зернобобовая культура, перспективная для возделывания в России. Его плоды содержат значительное количество жирных масел и белков, зеленые бобы могут служить ценным пищевым продуктом, это древняя кормовая культура (в настоящее время используется гуаровая мука и немолотый гранулированный корм), но наиболее востребовано это растение как источник гуаровой камеди, которая представляет собой полисахарид, образованный галактозой и маннозой (галактоманнан) и содержится в эндосперме семян. Гуар в основном выращивают в Индии, однако из-за высокого спроса на камедь это растение пытаются культивировать в других районах с подходящим климатом: в США, Судане, Афганистане, Кении, Пакистане, Австралии, а также на юге России. Известно, что продуктивность бобовых культур зависит не только от климатических условий, но и от эффективности симбиоза с ризобиями, которая в том числе определяется азотфиксирующей активностью и конкурентоспособностью микросимбионтов, а также их комплементарностью к определенному сорту...
Гуар, клубеньковые бактерии, ген 16s ррнк, its-регион, гены "домашнего хозяйства", симбиоз
Короткий адрес: https://sciup.org/142220076
IDR: 142220076 | УДК: 579.64:631.461.52 | DOI: 10.15389/agrobiology.2018.6.1285rus
Isolation and identification of root nodule bacteria from guar Cyamopsis tetragonoloba (L.) Taub
Cyamopsis tetragonoloba (guar) belongs to the family Fabaceae and is one of the promising crops for cultivation in Russia. Beans contain a large number of protein and fatty oil content, green beans can serve as a valuable source of food and feeds (as seed flour and not ground granulated feeds), but the plant is more in demand as a source of guar gum, which is a polysaccharide formed by galactose and mannose (galactomannan) and is contained in the endosperm of the seeds of this plant. Guar gum is widely used in various industries: food, textile, cosmetic, oil and other. Guar comes from India, where approximately 80 % of the world's production of guar gum is obtained. However, due to high demand, the plant is cultivated throughout the world in areas with a suitable climate (the USA, Sudan, Kenya, Pakistan, Australia), including in the south of the Russian Federation. It is known that the productivity of leguminous crops depends not only on climatic conditions, but also on the effectiveness of symbiosis of plants with nodule bacteria (rhizobia), which is determined by the nitrogen-fixing activity and competitiveness of strains, as well as their complementarity to a particular variety...
Текст научной статьи Выделение и идентификация клубеньковых бактерий гуара Cyamopsis tetragonoloba (L.) Taub
Циамопсис четырехкрыльниковый Cyamopsis tetragonoloba (L.) Taub. (гуар, гороховое дерево, или индийская акация) — однолетняя зернобобовая культура с высоким содержанием белка из трибы Indigofereae семейства Fabaceae . Помимо C. tetragonoloba , род Cyamopsis включает еще 4 вида (С. dentata , С. psoraloides , С. senegalensis и С. serrate ) с меньшим промышленным значением (1, 2). Выращивается как овощная культура и может употребляться на корм животным. Гуар — азотфиксирующее растение и служит хорошим предшественником в севообороте. Особую ценность представляет гуаровая камедь (используется как натуральный загуститель и эмульгатор в пищевой, медицинской, текстильной и целлюлозно-бумажной промышленности, при производстве косметики, взрывчатых веществ, а также как поверхностно-активное вещество с высокой вязкостью в
Исследование выполнено при поддержке Минобрнауки в рамках проекта RFMEFI60417X0168 (соглашение ¹ 14.604.21.0168). Долгосрочное хранение штаммов осуществляется в рамках Программы по развитию и инвентаризации биоресурсных коллекций научными организациями.
угольной и нефтегазовой промышленности) (3), спрос на которую постоянно растет: по данным за 2014-2016 годы, ежегодная потребность в камеди составляет около 1,5 млн т, а в 2016 году импорт гуаровой камеди в Россию превысил 15 тыс. т (4). Родина циамопсиса и основной поставщик гуаровой камеди — Индия, хотя растение также культивируется в Пакистане, Судане, Африке, Австралии, Цейлоне, Афганистане и США (1). В Россию гуар был завезен в середине 1920-х годов (5), но широкого распространения не нашел из-за недостаточных знаний о технологии ее возделывания (6). В последние годы интерес к промышленному выращиванию гуара отмечается в Северо-Кавказском регионе России, в Краснодарском крае, Ростовской области и в Крыму (3).
Известно, что производительность бобовых культур зависит от эффективности их симбиоза с клубеньковыми бактериями, которую в том числе определяет азотфиксирующая активность, вирулентность, конкурентоспособность, а также комплементарность (специфичность) штаммов микросимбионтов к определенному сорту растения. Применение активных штаммов в качестве инокулянтов обеспечивает интенсивную азот-фиксацию, способствует усилению фотосинтеза и, как следствие, приводит к увеличению урожайности растений (7). Использование ризобий для инокуляции особенно важно при возделывании бобовых на новых территориях, где в почве отсутствуют необходимые микросимбионты. Например, при попытке выращивания сои культурной ( Glycine max ) в нетипичных для вида географических зонах России клубеньки на корнях практически не формировались, и для повышения урожая и содержания белка в растительной массе и зерне потребовалось внесение с семенами специфических микросимбионтов (8). Согласно исследованиям прошлых лет, инокуляция произрастающего в Судане гуара штаммами Bradyrhizobium spр. оказывала заметное положительное влияние на развитие растений, значительно увеличивала количество клубеньков, сухую массу растений, общее содержание азота и урожай семян (9).
Мы полагаем, что для успешной интродукции гуара в России требуется (наравне с изучением подходящих почвенно-климатических условий и разработкой технологий возделывания) исследование микросимбионтов этой культуры и последующая селекция эффективных штаммов.
В настоящей работе мы впервые выделили из клубеньков гуара ри-зобии вида Bradyrhizobium elkanii . С помощью секвенирования последовательностей 16S рДНК, ITS-региона и генов «домашнего хозяйства» atpD , dnaK и recA у изолятов определена таксономическая принадлежность и описана генетическая разнородность.
Цель работы состояла в получении и филогенетическом анализе микросимбионтов растений Cyamopsis tetragonoloba .
Методика. Семена гуара (получены во Всероссийском институте генетических ресурсов растений им. Н.И. Вавилова, г. Санкт-Петербург) скарифицировали и поверхностно стерилизовали в 98 % H 2 SO 4 в течение 10 мин, затем тщательно промывали стерильной водопроводной водой и проращивали на фильтровальной бумаге в чашках Петри при температуре 25 ° C в темноте в течение 3 сут. Проростки переносили в пластиковые сосуды (12 сосудов по 4 проростка), содержащие 100 г стерильного вермикулита. В каждый сосуд добавляли 5 мл водной вытяжки, полученной из 12 образцов почв, собранных в провинции Раджастан (Индия).
Растения культивировали в течение 45 сут в фитокомнате с относительной влажностью 60 % при 4-уровневом режиме освещенности и температуры: ночь (темнота, 18 ° C, 8 ч), утро (200 мкмоль • м - 2 • с - 1, 20 ° C, 2 ч), 1286
день (400 мкмоль • м - 2 • с - 1 , 23 ° C, 12 ч), вечер (200 мкмоль • м - 2 • с - 1, 20 ° C, 2 ч). Для освещения использовались лампы L 36W/77 FLUORA («Osram», Германия). По окончании эксперимента корни растений вынимали из вермикулита и промывали водопроводной водой. Образовавшиеся клубеньки отделяли от корней, поверхностно стерилизовали 1 мин в 70 % этаноле и гомогенизировали в стерильной водопроводной воде.
Штаммы ризобий выделяли из гомогенатов клубеньков по традиционной методике (10) и с использованием модифицированного маннит-но-дрожжевого агара YMSA (11) с добавлением 0,5 % янтарной кислоты. Культуры выращивали при 28 ° С. Все изоляты депонированы в Ведомственной коллекции полезных микроорганизмов сельскохозяйственного назначения (ВКСМ) и размещены на Станции низкотемпературного автоматизированного хранения биологических образцов (Liconic Instruments, Лихтенштейн) (12). Информация о штаммах доступна в Интернет-базе данных ВКСМ (13).
Видовую принадлежность штаммов определяли на основе секвенирования гена 16S рРНК (rrs), а также последовательности между генами 16S и 23S рРНК (ITS-региона). Для амплификации 16S рДНК (фрагмент гена около 1500 п.н.) использовали праймеры fD1 (5´-AGAGTTTGATC-CTGGCTCAG-3´) и rD1 (5´-AAGGAGGTGATCCAGCC-3´), для амплификации ITS региона (800 п.н.) — праймеры FGPL-132 (5'-CCGGGTTT-CCCCATTCGG-3') и FGPS1490-72 (5'-TGCGGCTGGATCCCCTCCTT-3´). Для уточнения таксономического положения и изучения генетической гетерогенности штаммов были секвенированы гены «домашнего хозяйства» atpD, recA и dnaK. Амплификацию генов «домашнего хозяйства» проводили с помощью праймеров atpD352F и atpD871R, recA63FD и recA504RD (14), dnaK1466Fd и dnaK1777Rd (15). Полученные ПЦР-продукты выделяли из геля и очищали (16) для последующего секвенирования (генетический анализатор ABI PRISM 3500xl, «Applied Biosystems», США). Поиск гомологичных последовательностей выполняли с помощью базы данных NCBI GenBank и программы BLAST (17). Филогенетическое дерево конструировали в программе MEGA5 методом Neighbor-Joining (18). Пары последовательностей сравнивали по числу различающихся нуклеотидов. Чтобы оценить уровни поддержки кластеров, проводили бутстреп-анализ на основе 1000 повторов. Полученные последовательности депонированы в базе данных GenBank под номерами МH938226-MH938235 для гена rrs, MH938704-MH938713 — для последовательности ITS-региона, MH982271-MH982280 — для гена atpD, MH982261-MH982270 — для гена dnaK и MH982251-MH982260 — для гена recA.
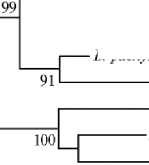
Результаты. Из корневых клубеньков растений гуара было выделено 10 бактериальных штаммов: по одному штамму из каждого образца почв. В двух образцах почв на корнях растений клубеньков не обнаружили. Анализ нуклеотидной последовательности гена 16S рРНК показал, что все штаммы принадлежат к роду Bradyrhizobium и формируют монофилетический статистически достоверный кластер с уровнем поддержки 99 % (рис. 1). Помимо изолятов, кластер включал типовые штаммы B. elkanii USDA 76T, B. jicamae PAC68T, B. lablabi CCBAU 23086T, B. pachyrhizi PAC48T и B. tro-piciagri SEMIA 6148T. Однако максимальная степень rrs-гомологии у новых изолятов (100 %) наблюдалась только с двумя типовыми штаммами — B. el-kanii USDA 76T и B. pachyrhizi PAC48T (табл.). Со штаммом B. tropiciagri SEMIA 6148Т сходство по rrs гену было ниже и варьировало от 98,4 до 99,3 %, со штаммами B. jicamae PAC68T и B. lablabi CCBAU 23086T rrs- гомология была ниже 99 % (см. табл.).
Bradyrhizobium pachyrhiy PAC48T (AY624135) ----------------В. tropiciagri SEMIA 6148T (AY904753)
Cte-503
Cte-501
Cte-497
Cte-496
^ Cte-499
Cte-502
-----B. jicamae PAC68T (NR043036)
Cte-495
____________ 99 Cte-498
Cte-500
Cte-504
B. elkanii USDA 76T (U35000) — B. lablabi CCBAU 230861 (GU433448)
B. retamae Rol9T (KC247085)
[— B. japonicum USDA 6T (U69638)

ft daqingense CCBAU I5774T (HQ231274)
----------B. yuanmingense B071T (AF193818)
•B. canariense ВТA- IT (AJ558025)
gj ,B. huanghuaihaiense CCBAU 233O3T (HQ231463)
S. iriomotense EK05T (AB300992)
r B. betae PL7HG1T (AY372184)
0.002
B. rifense CTAW71T (EU561074)
91 La cytisi CTAW11T (EU561065)
Рис. 1. rrs -Филограмма штаммов, выделенных из клубеньков гуара Cyamopsis tetragonoloba, а также представителей родственных видов рода Bradyrhizobium . Типовые штаммы отмечены литерой Т. Штаммы гуара отмечены жирным шрифтом (метод Neighbour-Joining).
Гомология (%) последовательностей генов у изолятов, выделенных из клубеньков гуара Cyamopsis tetragonoloba, и у ближайших типовых штаммов рода Bradyrhizobium
|
Типовой штамм |
Локус |
Изолят С |
|||||||||
|
495 |
496 |
497 |
498 |
499 |
500 |
501 1 |
502 |
503 |
504 |
||
|
B. elkanii USDA76T |
rrs |
100 |
100 |
100 |
100 |
100 |
100 |
100 |
100 |
100 |
100 |
|
ITS |
92,2 |
93,0 |
92,3 |
92,6 |
92,9 |
92,1 |
94,2 |
91,3 |
94,3 |
94,3 |
|
|
atpD |
96,1 |
96,5 |
97,4 |
97,8 |
97,8 |
96,8 |
96,0 |
96,2 |
96,3 |
97,0 |
|
|
dnaK |
98,8 |
98,8 |
98,4 |
97,9 |
98,4 |
98,7 |
98,4 |
98,7 |
98,7 |
99,2 |
|
|
recA |
94,4 |
95,0 |
94,5 |
94,5 |
94,7 |
94,5 |
95,7 |
94,4 |
96,1 |
94,6 |
|
|
B. pachyrhizi PAC48Т |
rrs |
100 |
100 |
100 |
100 |
100 |
100 |
100 |
100 |
100 |
100 |
|
ITS |
90,6 |
89,8 |
90,6 |
90,7 |
90,8 |
90,2 |
87,9 |
90,6 |
87,9 |
88,3 |
|
|
atpD |
96,1 |
95,3 |
95,1 |
95,7 |
95,1 |
94,9 |
95,7 |
95,8 |
95,8 |
95,1 |
|
|
dnaK |
99,6 |
99,6 |
99,2 |
98,6 |
99,1 |
99,0 |
98,7 |
99,3 |
99,0 |
99,2 |
|
|
recA |
93,8 |
93,9 |
93,9 |
93,9 |
93,6 |
93,8 |
94,8 |
93,9 |
94,7 |
93,6 |
|
|
B. tropiciagri SEMIA6148T |
rrs |
99,3 |
99,3 |
98,8 |
99,0 |
99,1 |
99,2 |
98,4 |
99,3 |
99,3 |
99,2 |
|
ITS |
92,1 |
91,9 |
91,8 |
91,5 |
91,9 |
91,5 |
91,4 |
91,1 |
90,8 |
90,8 |
|
|
atpD |
96,9 |
95,7 |
95,7 |
96,2 |
96,3 |
95,3 |
96,6 |
96,8 |
96,8 |
95,3 |
|
|
dnaK |
96,6 |
96,7 |
96,3 |
96,7 |
96,8 |
96,7 |
96,1 |
97,0 |
96,4 |
96,5 |
|
|
recA |
94,7 |
95,2 |
95,2 |
94,8 |
94,4 |
94,8 |
95,5 |
94,6 |
95,9 |
94,4 |
|
Для уточнения видовой принадлежности штаммов, а также изучения их генетической разнородности был проведен анализ последовательностей трех генов «домашнего хозяйства»: atpD и recA, кодирующих соответственно β -субъединицу мембранной АТФ-синтазы и ДНК-рекомбиназу
(14), а также гена dnaK, который кодирует шаперон, предотвращающий агрегацию белков и обеспечивающий их рефолдинг при тепловом повреждении (15). Дендрограмма, построенная на основе объединенных последовательностей генов atpD , dnaK и recA , представлена на рисунке 2.
Cte-497
Cte-500
----Cte-496
Ctc-499
Cte-504
----Ctc-498
Cte-495
98l Ctc-502
Cte-501
tool Cte-503
Bradyrhitobium dkanii USDA 76T

B. pachyrhiy PAC48T
B. troptciagri SEMIA 6148T
-
B. retamae Rol9T
B. labiabi CCBAU 23086T
B. jicamae PAC68T
-
B. daqingense CCBAU 15774T
-
B. japonicum USDA 6T
B. canariense LMG 22265T
B. betae LMG 219871
0,01
Рис. 2. Филограмма, построенная на основе объединенных последовательностей генов atpD , dnaK и recA у штаммов, выделенных из клубеньков гуара Cyamopsis tetragonoloba , а также представителей родственных видов рода Bradyrhizobium . Типовые штаммы отмечены литерой Т. Штаммы гуара отмечены жирным шрифтом (метод Neighbour-Joining).
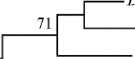
Все изоляты кластеризовались при уровне поддержки 100 % вместе с типовыми штаммами B. elkanii USDA 76Т, B. pachyrhizi PAC48Т и B. tropiciagri SEMIA 6148T. В пределах этой группы были выявлены два статистически достоверных субкластера, сформированных изолятами Cte-501 и Cte-503, а также Cte-495 и Cte-502 (уровни поддержки субкластеров соответственно 100 и 98 %). Максимальная гомология по гену atpD между новыми изолятами и типовыми штаммами составила 97,8 % для B. elkanii, 96,9 % — для B. tropiciagri и 96,1 % — для B. рachyrhizi (табл.) . Гомология по гену dnaK варьировала от 98,4 до 99,2 % для B. elkanii , от 98,7 до 99,6 % — для B. рachyrhizi и от 96,1 до 97,0 % — для B. tropiciagri . По гену recA величина максимальной гомологии между изолятами и типовыми штаммами равнялась 96,1 % для B. elkanii , 95,9 % — для B. tropiciagri и 94,8 % — для B. рachyrhizi (см. табл. 1).
Анализ последовательности ITS-региона часто используют в качестве дополнительного метода идентификации микроорганизмов (11, 19).
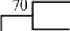
По результатам выполненного нами сравнения по ITS-региону все изоля-ты кластеризовались вместе с типовым штаммом B. elkanii USDA 76Т при уровне статистической поддержки 100 % (рис. 3). Максимальная гомология по ITS-региону между новыми изолятами и типовыми штаммами B. elkanii, B. tropiciagri и B. рachyrhizi составляла соответственно 94,3, 92,1 и 90,8 % (см. табл.).
pCte-498 — Cte-504
Cte-499
Cte-496
Cte-501
Cle-503
Ctc-500
Cte-495
Cte-497
0.01
Cte-502
-------Bradyrhteobium elkanii USDA 76T (AB509379)
B. paehyrhfci PAC 48 T (AY628092)
-B. troptciagrt SEMIA 61481 (FJ391100)
-----A retamae Rol9T (KF638356)
B. jicamae PAC68(AY628094) --------------B. lablabl CCBAU 23086T (GU433583)
B. cytisi CTAW11T (KC247124)
—B. rifense CT№N1W (KC247123)
Рис. 3. ITS-филограмма штаммов, выделенных из клубеньков гуара Cyamopsis tetragonoloba, а также представителей родственных видов рода Bradyrhizobium . Типовые штаммы отмечены литерой Т. Штаммы гуара отмечены жирным шрифтом (метод Neighbour-Joining).
Название вида Bradyrhizobium elkanii предложено в 1992 году (20) для гомологичной группы штаммов в пределах существующего вида B. ja-ponicum, описанного для азотфиксирующих микросимбионтов сои Glycine max (21). Оба вида относятся к доминирующему роду, способному ноду-лировать большинство видов трибы дроковых Genisteae (22), к наиболее известным представителям которой относятся люпин ( Lupinus ), ракитник ( Cytisus ) и дрок ( Genista ). Однако штаммы B. elkanii были также обнаружены в клубеньках бобовых растений из трибы Indigofereae , к которой относится и гуар: у вигны ( Vigna unguiculata, V. radiate ), палисандра ( Dalbergia odorifera ) и десмодиума ( Desmodium incanum ) (23-25) . Ранее были описаны два штамма — XBD2 SARCC-388 и ENNRI 16A, выделенные из циамо-псиса, способные к эффективной азотфиксации и идентифицированные соответственно как B. japonicum и Bradyrhizobium sp. (26, 27). Однако в основном исследования гуара проводятся для изучения и улучшения свойств камеди (28), устойчивости к болезням и селекции высокопродуктивных
сортов (2, 29). Видимо, это связано с достаточным количеством аборигенных штаммов-микросимбионтов в местах традиционного возделывания культуры и отсутствием необходимости в инокуляции растений. В связи с этим следует подчеркнуть, что при интродукции гуара на новые территории симбиотические растительно-микробные взаимоотношения, благодаря которым культура получает азотное питание, становятся важнейшим фактором повышения ее продуктивности.
Таким образом, по результатам секвенирования гена rrs десять штаммов, выделенных в вегетационном опыте из корневых клубеньков гуара, отнесены к роду Bradyrhizobium. С помощью дополнительных методов молекулярно-генетической идентификации (секвенирования последовательности ITS-региона и трех генов «домашнего хозяйства» dnaK, recA и atpD ) удалось уточнить таксономическое положение изолятов и показать их принадлежность к виду Bradyrhizobium elkanii. Ранее представители этого вида не были описаны как микросимбионты циамопсиса четырех-крыльникового. Секвенирование ITS-региона и генов «домашнего хозяйства» продемонстрировало генетическую разнородность природной популяции микросимбионтов гуара, что может свидетельствовать о различиях в симбиотических взаимоотношениях между растениями и изолированными штаммами. Дальнейшее изучение генетического разнообразия, морфологокультуральных, физиолого-биохимических, хозяйственно полезных свойств клубеньковых бактерий гуара позволит расширить знания о филогении микросимбионтов этой относительно новой для России культуры, а также проводить селекцию эффективных штаммов, улучшающих рост растений и качество продукции в новых регионах возделывания.
Авторы благодарны П.А. Белимовой за ценную помощь в проведении вегетационного опыта.
Список литературы Выделение и идентификация клубеньковых бактерий гуара Cyamopsis tetragonoloba (L.) Taub
- Лебедь Д.В., Костенкова Е.В., Волошин М.И. Агрономическое обоснование размещения посевов Cyamopsis tetragonoloba L. на юге европейской части России. Таврический вестник аграрной науки, 2017, 1(9): 53-63.
- Дзюбенко Н.И., Дзюбенко Е.А., Потокина Е.К., Булынцев С.В. Гуар Cyamopsis tetrago-nolоba (L.) Taub.: характеристика, применение, генетические ресурсы и возможность интродукции в России (обзор). Сельскохозяйственная биология, 2017, 52(6): 1116-1128 ( ) DOI: 10.15389/agrobiology.2017.6.1116rus
- Булынцев С.В., Вальяникова Т.И., Силаева О.И., Копоть Е.И., Пимонов К. И. Гуар -новая бобовая культура для России. Мат. Всероссийской науч.-прак. конф. «Инновации в технологиях возделывания сельскохозяйственных культур». Донской ГАУ, пос. Персиановский, 2017: 167-172.
- Старцев В.И., Ливанская Г.А., Куликов М.А. Перспективы возделывания гуара Cyamopsis tetragonoloba L. в России. Вестник Российского государственного аграрного заочного университета, 2017, 24(29): 11-15.
- Вавилов Н.И. Интродукция растений в советское время и ее результаты. Избранные труды. Том V. М.-Л., 1965: 674-689.
- Волошин М.И., Лебедь Д.В., Брусенцов А.С. Результаты интродукции нового бобового растения -гуара (Cyamopsis tetragonoloba (L.)Taub.). Труды Кубанского государственного аграрного университета, 2016, 1(58): 84-91.
- Стамбульская У.Я. Влияние бактерий Rhizobium leguminosarum bv. viciae на морфометрические показатели роста гороха. Режим доступа: http://bio-x.ru/articles/simbioz-bakteriy-rhizobium-leguminosarum-i-rasteniy-goroha. Без даты.
- Жакеева М.Б., Бекенова У.С., Жумадилова Ж.Ш., Шорабаев Е.Ж., Абдиева К.М., Саданов А.К. Влияние разных доз биопрепаратов на урожайность и биометрический показатель люцерны. Современные проблемы науки и образования, 2015, 5. Режим доступа: http://www.science-education.ru/ru/article/view?id=21887. Без даты.
- Elsheikh El.A.Al., Ibrahim K. The effect of Bradyrhizobium inoculation on yield and seed quality of guar (Cyamopsis tetragonoloba L.). Food Chem., 1999, 65(2): 183-187 ( )
- DOI: 10.1016/S0308-8146(98)00192-7
- Novikova N., Safronova V. Transconjugants of Agrobacterium radiobacter harboring sym genes of Rhizobium galegae can form an effective symbiosis with Medicago sativa. FEMS Microbiol. Lett., 1992, 93(3): 261-268 ( )
- DOI: 10.1016/0378-1097(92)90472-Z
- Safronova V.I., Kuznetsova I.G., Sazanova A.L., Kimeklis A.K., Belimov A.A., Andronov E.E., Pinaev A.G., Chizhevskaya E.P., Pukhaev A.R., Popov K.P., Willems A., Tikhonovich I.A. Bosea vaviloviae sp. nov., a new species of slow-growing rhizobia isolated from nodules of the relict species Vavilovia formosa (Stev.) Fed. Antonie van Leeuwenhoek, 2015, 107(4): 911-920 ( )
- DOI: 10.1007/s10482-015-0383-9
- Safronova V.I., Tikhonovich I.A. Automated cryobank of microorganisms: Unique possibilities for long-term authorized depositing of commercial microbial strains. In: Microbes in applied research: current advances and challenges/A. Mendez-Vilas (ed.). World Scientific Publishing Co, Hackensack, 2012.
- Электронная база данных Ведомственной коллекции полезных микроорганизмов сельскохозяйственного назначения (ВКСМ). Режим доступа: http://www.arriam.spb.ru. Без даты.
- Gaunt M.W., Turner S.L., Rigottier-Gois L., Lloyd-Macgilp S.A., Young J.P. Phylogenies of atpD and recA support the small subunit rRNA-based classification of rhizobia. Int. J. Syst. Evol. Micr., 2001, 51(6): 2037-2048 ( )
- DOI: 10.1099/00207713-51-6-2037
- Stepkowski T., Czaplińska M., Miedzinska K., Moulin L. The variable part of the dnaK gene as an alternative marker for phylogenetic studies of rhizobia and related alpha Proteobacteria. Syst. Appl. Microbiol., 2003, 26(4): 483-494 ( )
- DOI: 10.1078/072320203770865765
- Stępkowski T., Zak M., Moulin L., Króliczak J., Golińska B., Narożna D., Safronova V.I., Mądrzak C.J. Bradyrhizobium canariense and Bradyrhizobium japonicum are the two dominant rhizobium species in root nodules of lupin and serradella plants growing in Europe. Syst. Appl. Microbiol., 2011, 34(5): 368-375 ( )
- DOI: 10.1016/j.syapm.2011.03.002
- GenBank sequence database. The National Center for Biotechnology Information. Available https://blast.ncbi.nlm.nih.gov/Blast.cgi. Без даты.
- Tamura K., Peterson D., Peterson N., Stecher G., Nei M., Kumar S. MEGA5: Molecular evolutionary genetics analysis using Maximum Likelihood, evolutionary distance, and Maximum Parismony methods. Mol. Biol. Evol., 2011, 28(10): 2731-2739 ( )
- DOI: 10.1093/molbev/msr121
- Safronova V.I., Sazanova A.L., Kuznetsova I.G., Belimov A.A., Andronov E.E., Chirak E.R., Popova J.P., Verkhozina A.V., Willems A., Tikhonovich I.A. Phyllobacterium zundukense sp. nov., a novel species of rhizobia isolated from root nodules of the legume species Oxytropis triphylla (Pall.) Pers. Int. J. Syst. Evol. Micr., 2018, 68(5): 1644-1651 ( )
- DOI: 10.1099/ijsem.0.002722
- Kuykendall L.D., Saxena B., Devine T.E., Udell S.E. Genetic diversity in Bradyrhizobium japonicum Jordan 1982 and a proposal for Bradyrhizobium elkanii sp. nov. Can. J. Microbiol., 1992, 38(6): 501-505 ( )
- DOI: 10.1139/m92-082
- Jordan D.C. Transfer of Rhizobium japonicum Buchanan 1980 to Bradyrhizobium sp. nov., a genus of slow-growing, root nodule bacteria from leguminous plants. Int. J. Syst. Bacteriol., 1982, 32(1): 136-139 ( )
- DOI: 10.1099/00207713-32-1-136
- Stępkowski Т., Banasiewicz J., Granada C.E., Andrews M., Passaglia L.M.P. Phylogeny and phylogeography of rhizobial symbionts nodulating legumes of the Tribe Genisteae. Genes, 2018, 9(3): 163 ( )
- DOI: 10.3390/genes9030163
- Zhang Y.F., Chang E.T., Tian C.F., Wang F.Q., Han L.L., Chen W.F., Chen W.X. Bradyrhizobium elkanii, Bradyrhizobium yuanmingense and Bradyrhizobium japonicum are the main rhizobia associated with Vigna unguiculata and Vigna radiate in the subtropical region of China. FEMS Microbiol. Lett., 2008, 285(2): 146-154 ( )
- DOI: 10.1111/j.1574-6968.2008.01169.x
- Lu J., Yang F., Wang S., Ma H., Liang J., Chen Y. Co-existence of Rhizobia and diverse non-rhizobial bacteria in the rhizosphere and nodules of Dalbergia odorifera seedlings inoculated with Bradyrhizobium elkanii, Rhizobium multihospitium-like and Burkholderia pyrrocinia-like strains. Front. Microbiol., 2017, 21(8): 2255 ( )
- DOI: 10.3389/fmicb.2017.02255
- Toniutti M.A., Fornasero L.V., Albicoro F.J., Martini M.C., Draghi W, Alvarez F., Lagares A., Pensiero J.F., Del Papa M.F. Nitrogen-fixing rhizobial strains isolated from Desmodium incanum DC in Argentina: phylogeny, biodiversity and symbiotic ability. Syst. Appl. Microbiol., 2017, 40(5): 297-307 ( )
- DOI: 10.1016/j.syapm.2017.04.004
- Hassen A.I., Bopape F.L., Trytsman M. Nodulation study and characterization of rhizobial microsymbionts of forage and pasture legumes in South Africa. World Journal of Agricultural Research, 2014, 2(3): 93-100 ( )
- DOI: 10.12691/wjar-2-3-2
- Mohamed Ahmed T.H., Elsheikne A.E., Mahdi A.A. The in vitro compatibility of some Rhizobium and Bradyrhizobium strains with fungicides. African Crop Science Conference Proceedings. Egypt, 2007, 18: 1171-1178 ( )
- DOI: 10.13140/2.1.3933.9208
- Sharma G., Sharma S., Kumar A., Al-Muhtaseb A.H., Naushad M., Ghfar A.A., Mola G.T., Stadler F.J. Guar gum and its composites as potential materials for diverse applications: A review. Carbohydrate Polym., 2018, 199: 534-545 ( )
- DOI: 10.1016/j.carbpol.2018.07.053
- Muthuselvi R., Shanthi A., Praneetha S. Mean performance of cluster bean (Cyamopsis tetragonoloba) genotypes for yield and quality parameters. International Journal of Chemical Studies, 2018, 6(2): 3626-3629.
