Выравнивание почвенных условий для развития растений при деструкции растительных остатков микробными препаратами

Автор: Свиридова О.В., Воробьев Н.И., Проворов Н.А., Орлова О.В., Русакова И.В., Андронов Е.Е., Пищик В.Н., Попов А.А., Круглов Ю.В.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Растение и почва
Статья в выпуске: 5 т.51, 2016 года.
Бесплатный доступ
Развитие современного сельского хозяйства направлено на получение стабильно высоких урожаев и производство семян высокого качества. В связи с этим разрабатываются прецизионные агротехнологии, выравнивающие почвенные условия. Индивидуальные фенотипические характеристики растений определяются локальными почвенными условиями вблизи корневых систем. Так, дисперсия высот растений зависит от пространственного распределения энергетических ресурсов и питательных веществ в почве и ограничивается генетической нормой. Она может уменьшаться, когда почвенные условия выравниваются. Мы предположили, что эффект микробиологического выравнивания почвенных условий (МВПУ) наблюдается при деструкции растительных остатков с использованием микробного препарата. До настоящего времени исследования в этом направлении не проводились. В представленной работе были получены практически значимые результаты благодаря использованию оригинального фрактального анализа молекулярно-генетических частотных данных для почвенных микробных сообществ. Нашей целью стало экспериментальное и теоретическое изучение МВПУ эффекта, возникающего после деструкции растительных остатков с использованием микробных препаратов, разработанных во Всероссийском НИИ сельскохозяйственной микробиологии. В I опыте (2011-2014 годы) изучали дисперсию высот растений ячменя после деструкции соломы ячменя препаратом баркон. Во II опыте использовали данные молекулярно-генетического анализа почвенных микробных сообществ после деструкции растительных остатков тремя микробными препаратами (баркон, багс и омуг). Препарат баркон содержал консорциум бактерий и грибов; багс представлял собой консорциум целлюлозолитических микроорганизмов, полученный на основе биологически активного грунта; омуг - микробное удобрение, полученное после биотехнологической переработки птичьего помета. Функциональную активность микробных сетей, возникающих при деструкции растительных остатков с использованием микробных препаратов, изучили с помощью фрактального анализа молекулярно-генети-ческих данных микробных сообществ в почве. Мы получили фрактальный таксономический портрет микробных сообществ и индекс эффективности функционирования микробных сетевых образований, которые были сформированы во время деструкции растительных остатков. Использование баркона для деструкции растительных остатков в I опыте привело к постепенному выравниванию почвенных условий и уменьшению дисперсии высот растений. Без применения препарата дисперсия высот растений возрастала с каждым годом. Следовательно, микробные препараты могут инициировать эффективные микробные сети, которые способны сохранить энергетические ресурсы и питательные вещества, распределив их равномерно в почве. Молекулярно-генетические данные II опыта подтвердили, что эффективность функционирования микробных сетей после использования баркона, багса и омуга значительно увеличивалась за счет лучшей организации деструктивных процессов. Таким образом, деструкция растительных остатков с помощью биопрепаратов - необходимое и эффективное дополнение современных агротехнологий. Она приводит к восстановлению необходимого количества энергетических ресурсов и питательных веществ в почве, выравниванию ресурсов в почвенном пространстве, повышению устойчивости урожайности и улучшению качества получаемой растительной продукции.
Деструкция растительных остатков с использованием микробных биопрепаратов, микробные деструктивные сообщества в почве, дисперсия индивидуальных высот растений, фрактально-таксономический портрет микробного сообщества, индекс функциональной эффективности микробных деструктивных сетей
Короткий адрес: https://sciup.org/142213969
IDR: 142213969 | УДК: 633.1:579.64 | DOI: 10.15389/agrobiology.2016.5.664rus
The alignment of soil's conditions for plant's development during microbial destruction of plant's residues by microbial preparations
Modern agriculture is developing in the direction of producing consistently high yields and high quality seed production. In this regard, the precision agricultural technologies are develop for the leveling a soil conditions. Individual phenotypic characteristics of plant are determined by the local soil conditions near their root's systems. As a result, the variance of plant's height is dependent on the spatial distribution of energy resources and nutrients in the soil. The variance of plant's height restricted to the genetic norm for this characteristic and it can been reduced when a soil conditions are leveled. In experiments with planting alfalfa was been shown, that the variance of plant's mass may decrease with an increase in the efficiency of plant-microbial symbiosis. Perhaps the plant-microbial symbiosis is able to level a soil conditions and selectively stimulate the plants by using of the microbial metabolites. We assume that the effect of microbiological leveling soil conditions (MLSC) may be observed during destruction of plant's residues using microbial preparation. Previously, MLSC effect has not studied. Therefore, these theoretical and experimental researches are new. In addition, we have obtained new practically important results thanks to the use of the original fractal analysis of molecular-genetic data of the soil microbial community. The goal of this work was experimental and theoretical study of MPSC effect arising after destruction of plant's residues with the using of the microbial preparations, which was been developed in the All-Russian Research Institute of Agricultural Microbiology. To achieve this goal were used the data of two experiments. In the first experiment, the variances of the barley plant's height were been investigated after the destruction of plant residues using a microbial preparation Barkon. Out the second experiment used data of the molecular-genetic analysis of soil microbial communities after the destruction of plant's residues using three microbial preparations: Barkon, Bags and Omug. The preparation Barkon contains the consortium of bacteria and fungi; Bugs is a consortium of cellulolytic organisms, derived from biologically active soil; Omug is the microbial fertilizer obtained after biotechnological processing of poultry manure. The functional activity of microbial networks arising during the destruction of plant's residues with using of microbial preparations was been studied using fractal analysis of molecular genetic data of microbial communities in soil. With using of the fractal analysis was been obtained the fractal taxonomic portrait of microbial communities and the index of the functional efficiency of microbial network formations, which were formed during destruction of plant's residues. The first experiment showed that the using of preparations for the destruction of plant's residues leads to a gradual leveling of the soil's conditions and to reducing of the variance of plant's heights, which were been grown on these soils. Without these preparations, the dispersion of plant's heights increases with each successive year. From this, it follows that these preparations may initiate the effective microbial networks that are able to save the energy resources and the nutrients distributing them evenly in the soil. The molecular-genetic data from second experiment confirmed that the functional efficiency of the microbial networks after using preparations significantly increases due to better organization of destructive processes. The results of this study suggest that the destruction of plant's residues by using of the special preparations is a necessary and effective complement of the modern precision agro technologies. Thus, the microbial preparations for destruction plant's residues start processes which lead to the restoration of the required level of energy resources and nutrients in the soil, to the leveling of resources in soil's space, to the increasing the stabili
Текст научной статьи Выравнивание почвенных условий для развития растений при деструкции растительных остатков микробными препаратами
Развитие современного сельского хозяйства направлено на получение стабильно высоких урожаев и производство семян высокого качества. В связи с этим разрабатываются прецизионные агротехнологии, выравнивающие почвенные условия (1-5).
∗ Исследования выполняли с использованием оборудования ЦКП «Геномные технологии, протеомика и клеточная биология» (ФГБНУ Всероссийский НИИ сельскохозяйственной микробиологии, г. Санкт-Петербург—Пушкин). Работа поддержана Российским научным фондом, грант ¹ 14-26-00094.
Вегетация растений, начиная с прорастания семян, зависит от локального состояния почвы в небольшой зоне, окружающей семена и корневую систему. Каждое растение характеризуется индивидуальными показателями высоты стеблей, числа вегетативных органов, массы надземной и корневой частей и т.д. В результате в популяциях наблюдается дисперсия (варьирование) количественных фенотипических признаков. Сверху она ограничивается допустимой нормой реакции растений на изменение экологических факторов (6) и может снижаться при выравнивании почвенных условий (7-10). Снижение вариабельности фенотипических признаков растений было обнаружено в 3-летних опытах с люцерной посевной ( Medicago sativa ) (11). Каждый год соотношение генотипической и фенотипической составляющих дисперсии массы растений изменялось. Наблюдаемое возрастание сорто-штаммовой (генотипической) составляющей и уменьшение случайной (фенотипической), возможно, было вызвано выравниванием почвенных условий в результате повышения эффективности микробно-растительного симбиоза, но также могло быть следствием адресной стимуляции микробными метаболитами (12, 13) тех растений в популяции, которые оказались в менее благоприятных условиях по сравнению с остальными.
Мы предполагаем, что эффект микробиологического выравнивания почвенных условий (МВПУ) связан с деструкцией растительных остатков микробными биопрепаратами. В этом случае происходит выборочное накопление микроорганизмами питательных ресурсов в почвенных микронишах, где изначально их было недостаточно. Возможно, МВПУ — следствие диффузии низкомолекулярных питательных веществ, возникающей при деструкции растительных остатков в зоны с пониженным содержанием питательных ресурсов. Очевидно, что на таких выравненных почвах растения должны показывать пониженную вариабельность количественных фенотипических признаков и высокое качество урожая (14).
Влияют ли на процессы МВПУ микробиологические препараты, вызывающие деструкцию растительных остатков в почве в предвегетацион-ный период года (без растений), до настоящего времени не исследовалось. Мы впервые описали эти эффекты, получив практически значимые результаты благодаря использованию оригинального фрактального анализа молекулярно-генетических частотных данных для почвенных микробных сообществ и коэффициента варьирования индивидуальных высот растений, выращенных на соответствующих почвах.
Цель настоящей работы — экспериментальное и теоретическое изучение эффекта микробиологического выравнивания почвенных условий при применении микробиологических биопрепаратов, инициирующих функционирование микробных сетей в почвах.
Методика. В I опыте (проведен в 2011-2014 годах) использовали экспериментальный образец микробиологического биопрепарата баркон для гумификации растительных остатков (разработан во Всероссийском НИИ сельскохозяйственной микробиологии — ВНИИСХМ), который состоял из консорциума бактерий и грибов (15). В сосуды с дерново-подзолистой почвой (по 3,5 кг, Сгум. 2,0 %, Nобщ. 0,19 %, рНсол. 5,6) вносили солому ячменя (СЯ) на глубину 0-3 см. Этот опыт включал следующие варианты: 1-й — контроль (без внесения СЯ), 2-й — внесение СЯ без инокуляции барконом, 3-й — внесение СЯ, инокулированной барконом (1 мл/10 г соломы). Измельченную СЯ (воздушно-сухая масса — 10 г/сосуд) во 2-м и 3-м вариантах вносили в сосуды 3 раза — осенью 2011, 2012 и 2013 годов. В каждом варианте было по 4 сосуда. Минеральные удобрения в течение опыта в сосуды не вносили. Растения ячменя (Hordeum L.) высаживали в сосуды весной 2012, 2013 и 2014 годов (по 30 шт/сосуд) и по окончании вегетации (осенью 2012, 2013 и 2014 годов) измеряли высоту надземной части растений (100-120 шт/вариант опыта). Данные обрабатывали методом дисперсионного анализа (16) и вычисляли коэффициент варьирования этого фенотипического признака для каждого варианта опыта.
Во II (модельном) опыте молекулярно-генетические данные по микробным сообществам использовали для изучения сетевой организации микроорганизмов, составляющих основу биопрепаратов для деструкции соло- мы злаковых (17). В сосуды на 1 кг окультуренной дерново-подзолистой почвы (Сгум. 4,02 %; Nобщ. 0,316 %; pHсол. 5,63) вносили по 3 г измельченной соломы ржи (СР) тремя способами — поверхностно либо заделывая в слое 0-3 см и 9-12 см. Для ускорения разложения солому обрабатывали препаратами багс, баркон и омуг согласно рекомендациям разработчиков (ВНИИСХМ). Багс представляет собой консорциум целлюлозолитических микроорганизмов на основе биологически активного грунта (18); омуг — микробное удобрение, полученное после биотехнологической переработки птичьего помета (19).
На 60-е сут проводили молекулярно-генетический анализ 12 почвенных образцов с использованием стандартной методики (19) и системы генетического анализа CEQ 8000 («Beckman Coulter International S.A.», Швейцария). Данные по частоте встречаемости оперативных таксономических единиц (ОТЕ) в почвенных образцах были подвергнуты модифицированному фрактальному анализу (21-26) для получения информации о характеристиках образующихся микробных сетевых структур. Таксономические данные в исследовании не использовались. Информация о таксо- номическом анализе этих же данных представлена ранее (17).
Результаты. В I опыте (1-й и 2-й варианты) коэффициент вариации высот растений (ВВР) ячменя возрастал от первого года наблюдения к третьему (табл. 1). Вероятно, общее количество питательных ресурсов в почве снижалось, вследствие чего повышалась вариабельность почвенных условий и индивидуальных высот растений.
1. Коэффициенты вариации высот растений (ВВР) ячменя при выращивании на почве с внесением растительных остатков в виде соломы ячменя (СЯ) и в зависимости от инокуляции биопрепаратом баркон
|
Вариант 1 |
Год |
Коэффициент ВВР, % |
|
Контроль (без внесения |
2012 |
19,8 |
|
СЯ) |
2013 |
25,5 |
|
2014 |
27,0 |
|
|
Внесение СЯ без инокуля- |
2012 |
13,2 |
|
ции барконом |
2013 |
14,9 |
|
2014 |
16,3 |
|
|
Внесение СЯ с инокуля- |
2012 |
17,1 |
|
цией барконом |
2013 |
14,5 |
|
2014 |
16,0 |
|
|
Доверительный интервал |
±0,2 |
В 3-м варианте коэффициент ВВР с годами, напротив, уменьшался (см. табл. 1). То есть применение биопрепарата постепенно повышало эффективность функционирования почвенного микробного деструктивного сообщества. Накопление питательных ресурсов для растений и выравнивание почвенных условий может длиться несколько лет (27).
Вполне вероятно, что при этом образуются низкомо- лекулярные питательные вещества, диффундирующие в микрониши с их пониженной концентрацией, что также могло приводить к нивелированию исходных различий между микронишами.
Микробиологическая деструкция растительных остатков может быть представлена как поэтапное деление органических молекул на все большее число уменьшающихся по размерам фрагментов. В качестве математического объекта, отображающего деструкцию молекул, предлагается исполь- зовать степенные ряды дробных чисел, относящиеся к фрактальным числовым рядам (28, 29). Например, ряд чисел (1, 1/2, 1/4, 1/8, ...) — это фрактальное множество, так как он образован по фиксированному для всех чисел множества правилу: каждое следующее число ряда в 2 раза меньше предыдущего.
Числа из фрактальных рядов условно можно сопоставить с размерами фрагментов молекул. Тогда позиция во фрактальном ряду будет означать номер этапа деструкции, а текущее число ряда — размеры фрагментов молекул, которые подвергаются деструкции на соответствующем этапе. Реальные процессы деструкции происходят со сменой правил фрагментации молекул. На одних этапах число фрагментов может удваиваться, на других — утраиваться, то есть мы предполагаем, что процессы микробиологической деструкции мультифрактальны (30-34).
Наши исследования показали, что для математического моделирования фрактальными рядами сложных деструктивных процессов органических молекул достаточно использовать только три первых члена фрактальных рядов. Это ограничение значительно упрощает идентификацию сложных деструктивных процессов с заранее неизвестными правилами фрагментации и числом этапов деструкции.
Мы считаем, что деструкция органических молекул растительных остатков микроорганизмами осуществляется при образовании мультифрак-тальных сетевых структур микроорганизмов (35, 36). При этом оказывается задействовано необходимое количество микроорганизмов, пропорциональное числу фрагментов, подвергающихся деструкции. Такое допущение позволяет проводить фрактальный анализ микробиологических деструктивных процессов по частотам встречаемости ОТЕ с использованием следующего математического правила, которое связывает частоты трех ОТЕ:
HeiL-HepI
ln{p max) ^(^max)
где pk — частота встречаемости микроорганизмов k -й ОТЕ, осуществляющих деструкцию органических молекул на k -м этапе ( k = 0, 1, 2); p max — частота встречаемости микроорганизмов доминирующей ОТЕ в почвенном сообществе; a — постоянный коэффициент, фрактальный индекс первичного деструктивного комплекса микроорганизмов (ПДК).
Логарифмическая зависимость [1] дает возможность выделить и идентифицировать три ОТЕ, объединенных в ПДК, в котором микроорганизмы осуществляют деструкцию органических молекул по единому правилу фрагментации. Для обнаружения нескольких ПДК в микробных сообществах требуется перебрать все возможные комбинации по три ОТЕ и выбрать те из них, которые удовлетворяют следующим равенствам:
ln ( P1) _ ln ( P 0 )
ln ( p max ) ln ( P max ) ln ( P 2 ) _ ln ( P 1 )
+ a
_ ln ( p max ) ln ( P max )
+ a ( ± d )
где 1 > p 0 > p 1 > p 2 — частоты ОТЕ, образующих ПДК, микроорганизмы которых осуществляют деструкцию органических молекул в три этапа по единому правилу фрагментации; 0 < d < 0,01 — допустимые пределы погрешности фрактального индекса ПДК для его идентификации, зависящие от точности измерения частотных параметров ОТЕ.
На рисунке (А) представлен упорядоченный ряд нормированных логарифмов частот встречаемости 22 ОТЕ из II опыта (вариант с применени-
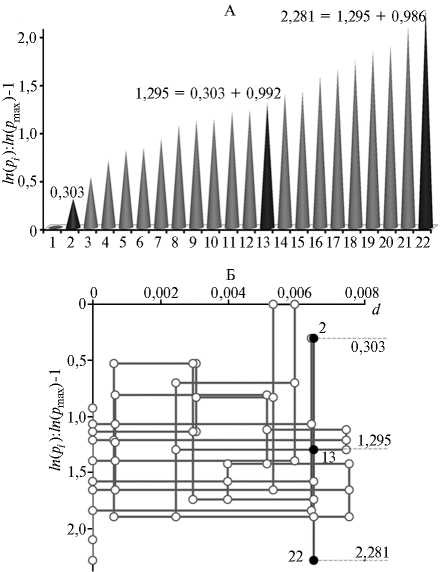
Упорядоченный ряд частот встречаемости отдельных таксономических единиц (ОТЕ) , выделенных молекулярно-генетическими методами (А) , а также фрактально-таксономический портрет бактериального почвенного сообщества при поверхностном расположении на почве соломы ржи, инокулированной биопрепаратом бар-кон (Б) : 1-22 — порядковые номера ОТЕ, d — погрешность фрактального индекса ПДК (первичный деструктивный комплекс) [2]; ln ( p max) , ln ( pi ) — натуральные логарифмы частот встречаемости ОТЕ соответственно с максимальной частотой встречаемости и порядковым номером i в ряду частот. ОТЕ ¹¹ 2, 13, 22 удовлетворяют равенствам [2].
ем баркона и расположением инокулированной СР на поверхности почвы). Для примера показаны три ОТЕ, частоты встречаемости которых удовлетворяют равенствам [2]. Следовательно, соответствующие микроорганизмы участвовали в деструкции растительных остатков по единому правилу фрагментации и образовывали ПДК.
Для визуализации всех ПДК в сообществе микроорганизмов была предложена следующая методика построения фрактально таксономического портрета микробных сообществ (см. рис., Б) (37). На портрете каждые три кружка, соединенные вертикальными линиями, представляют три ОТЕ в составе одного ПДК. Так отображается трехэтапная деструкция органических молекул соответствующими микроорганизмами по одному из правил фрагментации. Чем больше расстояние между кружками в ПДК, тем выше коэффициент деления на фрагменты на соответствующих этапах микробиологической деструкции и фрактальный индекс ПДК. Го- ризонтальные линии, соединяющие одни и те же ОТЕ, указывают на микроорганизмы, присутствующие одновременно в нескольких ПДК. При этом проекция каждого ПДК на горизонтальную ось d указывает на погрешность фрактального индекса соответствующего ПДК [2].
В нашем опыте фрактально-таксономический портрет деструктивного микробного сообщества (см. рис., Б) представлял собой сложное сетевое образование, состоявшее из 21 ПДК, образованных 22 ОТЕ. ОТЕ ¹ 7 и ¹ 22 вошли в ПДК по одному разу, остальные — по два раза и более. Следовательно, некоторые группы микроорганизмов могли функционировать одновременно в нескольких ПДК с разными правилами фрагментации молекул. Эти микроорганизмы, возможно, одновременно синтезировали несколько ферментов, что могло приводить к уменьшению скорости синтеза и снижению эффективности функционирования всего деструктивного процесса. Поэтому индекс эффективности функционирования микробных сетевых образований (ЭСО) мы предлагаем вычислять по следующей формуле:
I NET = ln ( NM ) : ln ( 3 ' N F ) ,
где INET — индекс ЭСО; NM — общее число ОТЕ, образовавших ПДК; NF — число ПДК в микробном сообществе.
Наибольшее значение индекса ЭСО равно единице, в микробном сообществе все ОТЕ присутствуют только в одном ПДК. То есть такое микробное деструктивное сообщество функционирует с максимальной специфичностью (одна группа микроорганизмов — одна деструктивная функция).
В таблице 2 приведены индексы ЭСО, характеризующие эффективность деструкции растительных остатков почвенными микробными сообществами, которые были инициированы микроорганизмами, входящими в состав препаратов багс, баркон и омуг. В контроле индекс ЭСО оказался меньше, чем в остальных вариантах, то есть микробиологические процессы были неорганизованы и малоэффективны. Использование биопрепаратов заметно повышало эффективность функционирования микробных сообществ, что подтверждалось возрастанием индекса ЭСО по отношению к контролю и вариантам опыта без их применения.
2. Индекс эффективной сетевой организации (ЭСО) деструктивных микробных сообществ в почве при внесении соломы ржи (СР) на разную глубину и в зависимости от инокуляции различными биопрепаратами
|
Вариант |
Глубина закладки СР в почву, см |
п I NET (индекс ЭСО) [3] |
|
Контроль (без внесения СР) |
0,634 |
|
|
Внесение СР без инокуляции био- |
0 |
0,662 |
|
препаратами |
0-3 |
0,676 |
|
9-12 |
0,700 |
|
|
Внесение СР, инокулированной |
0 |
0,746 |
|
барконом |
0-3 |
0,701 |
|
9-12 |
0,669 |
|
|
Внесение СР, инокулированной |
0 |
0,686 |
|
багсом |
0-3 |
0,689 |
|
9-12 |
0,669 |
|
|
Внесение СР, инокулированной |
0 |
0,630 |
|
омугом |
0-3 |
0,705 |
|
9-12 |
0,753 |
|
|
Доверительный интервал |
±0,003 |
При закладке соломы в верхние слои почвы, наибольшие значения индекса ЭСО наблюдались при применении биопрепаратов баркон и багс. Возможно, образование сетевых структур, инициированных этими биопрепаратами, происходило с привлечением аэробных почвенных микроорганизмов как поставщиков энергетических ресурсов (38). Так, в состав омуга входили микроорганизмы из кишечника птицы, то есть из анаэробной среды. При использовании омуга наибольшее значение индекса ЭСО наблюдалось при глубокой (на 9-12 см) закладке соломы в почву.
Сопоставляя данные I и II опытов, можно утверждать следующее. Основная причина отрицательной динамики коэффициента ВВР по годам в 3-м варианте I опыта заключалась в постепенном возрастании эффективности функционирования сетевых деструктивных микробных образований, инициированных биопрепаратом. Возможно, длительная настройка микробного сообщества на оптимальную деструктивную деятельность была направлена не только на деструкцию растительных остатков, но и на восстановление необходимого количества энергетических ресурсов и питательных веществ в почве, а также их выравнивание в почвенном пространстве.
Таким образом, деструкция растительных остатков с помощью специальных микробиологических биопрепаратов позволяет эффективно и с меньшими затратами решать задачи выравнивания количества энергетических ресурсов и питательных веществ в почве. Кроме того, инициация деструктивных микробиологических процессов способствует восстановлению необходимого уровня энергетических ресурсов и содержания питатель- ных веществ в почве, повышению стабильности урожаев и улучшению качества получаемой растительной продукции.
Список литературы Выравнивание почвенных условий для развития растений при деструкции растительных остатков микробными препаратами
- Якушев В.П., Якушев В.В. Информационное обеспечение точного земледелия. СПб, 2007 (ISBN 5-86763-181-8).
- McBratney A., Whelan B., Ancev T., Bouma J. Future directions of Precision Agriculture. Precision Agriculture, 2005, 6: 7-23 ( ) DOI: 10.1007/s11119-005-0681-8
- Zhang X., Shi L., Jia X., Seielstad G., Helgason C. Zone mapping application for precision-farming: A decision support tool for variable rate application. Precision Agriculture, 2010, 11(2): 103-114 ( ) DOI: 10.1007/s11119-009-9130-4
- Gebbers R., Adamchuk V.I. Precision agriculture and food security. Science, 2010, 327: 828-831 ( ) DOI: 10.1126/science.1183899
- Nikkilä R., Seilonen I., Koskinen K. Software architecture for farm management information systems in precision agriculture. Comput. Electron. Agric., 2010, 70(2): 328-336 ( ) DOI: 10.1016/j.compag.2009.08.013
- Инге-Вечтомов С.Г. Генетика с основами селекции: учебник для студентов высших учебных заведений. СПб, 2010.
- Griffiths A.J.F., Miller J.H., Suzuki D.T., Lewontin R.C., Gelbart W.M. Norm of reaction and phenotypic distribution. An introduction to genetic analysis. NY, 2000 (ISBN 0-7167-3520-2).
- Авдеев Ю.И., Морозова Л.В., Авдеев А.Ю., Кигашпаева О.П. Закон внутривидовой фенотипической изменчивости количественных признаков растений. Астраханский вестник экологического образования, 2014, 28(2): 114-120.
- Авдеев Ю.И. Генетический анализ количественных признаков растений. Астрахань, 2003.
- Гужов Ю.Л., Гнейм А.Р. Закономерности варьирования количественных признаков у гороха, обусловленные модификациями и генетическими различиями. Генетика, 1982, 18(2): 82-99.
- Тихонович И.А., Проворов Н.А. Симбиозы растений и микроорганизмов: молекулярная генетика агросистем будущего. СПб, 2009.
- Lugtenberg B.J.J., de Weger L.A., Bennett J.W. Microbial stimulation of plant growth and protection from disease. Curr. Opin. Biotech., 1991, 2(3): 457-464 ( ) DOI: 10.1016/S0958-1669(05)80156-9
- Мордухова Е.А., Кочетков В.В., Поликарпова Ф.Я., Боронин А.М. Синтез индолил-3-уксусной кислоты ризосферными псевдомонадами: влияние плазмид биодеградации нафталина. Прикладная биохимия и микробиология, 1998, 34(3): 287-292.
- Завалин А.А. Биопрепараты, удобрения и урожай. М., 2005.
- Воробьев Н.И., Свиридова О.В., Попов А.А., Русакова И.В., Петров В.Б. Граф-анализ генно-метаболических сетей микроорганизмов, трансформирующих растительные остатки в гумусовые вещества. Сельскохозяйственная биология, 2011, 3: 88-93.
- Воробьев Н.И., Проворов Н.А., Пищик В.Н., Свиридова О.В. Программа двухфакторного дисперсионного анализа биологических данных. Программа зарегистрирована в ФГУ ФИПС в отделе регистрации программ для ЭВМ № 2014661477 от 30.10.2014. Свидетельство о государственной регистрации программы для ЭВМ № 2014661477 от 30.10.2014. Режим доступа: http://www1.fips.ru/wps/portal/Registers/. Без даты.
- Орлова О.В., Андронов Е.Е., Воробьев Н.И., Колодяжный А.Ю., Москалевская Ю.П., Патыка Н.В., Свиридова О.В. Состав и функционирование микробного сообщества при разложении соломы злаковых культур в дерново-подзолистой почве. Сельскохозяйственная биология, 2015, 50(3): 305-314 ( , 10.15389/agrobiology.2015.3.305eng) DOI: 10.15389/agrobiology.2015.3.305rus
- Круглов Ю.В., Пароменская Л.Н. Микробиологические факторы биоремедиации почвы, загрязненной гербицидом прометрином. Сельскохозяйственная биология, 2011, 3: 76-80.
- Архипченко И.А., Барболина И.И., Дерикс П. Новая стратегия переработки отходов животноводства для получения биоудобрений. Доклады Россельхозакадемии, 1998, 6: 18-19.
- Брюханов А.А., Нетрусов А.И., Рыбак К.Е. Молекулярная микробиология. М., 2012 (ISBN 978-5-211-05486-8).
- Воробьев Н.И., Проворов Н.А., Свиридова О.В. Фрактальный анализ адаптационных свойств растительных компонентов биосистем. Мат. IV Всероссийской науч.-практ. конф. «Биологические системы: устойчивость, принципы и механизмы функционирования». Нижний Тагил, 2012, вып. 1: 103-106.
- Воскресенская О.Л., Скочилова Е.А., Копылова Т.И., Алябышева Е.А., Сарбаева Е.В. Организм и среда: факториальная экология. Йошкар-Ола, 2005 (ISBN 5-94808-149-4).
- Losa G.A., Merlini D., Nonnenmacher T.F., Weibel E.R. Fractals in biology and medicine. Vol. IV. Berlin, 2005.
- Akbar S. Fractal analysis in chemistry and biology. An Introduction. Lambert Academic Publisher, 2010 (ISBN: 978-3-8433-7905-2).
- Jones C.L., Jelinek H.F. Wavelet packet fractal analysis of neuronal morphology. Methods, 2001, 24(4): 347-358 ( ) DOI: 10.1006/meth.2001.1205
- Ristanović D., Nedeljkov V., Stefanović B.D., Milošević N.T., Grgurević M., Stulić V. Fractal and nonfractal analysis of cell images: comparison and application to neuronal dendritic arborization. Biological Cybernetics, 2002, 87(4): 278-288 ( ) DOI: 10.1007/s00422-002-0342-1
- Свиридова О.В., Воробьев Н.И., Попов А.А. Пролонгированное действие гумификации соломы микроорганизмами биопрепарата Баркон на адаптационные свойства растений ячменя. Мат. III Межд. науч. Интернет-конф. «Биотехнология. Взгляд в будущее». Казань, 2014, 2: 102-106.
- Шредер М. Фракталы, хаос, степенные законы. Миниатюры из бесконечного рая. Ижевск, 2001 (ISBN 5-93972-041-2).
- Богатых Б.А. Фрактальная природа живого: системное исследование биологической эволюции и природы сознания. М., 2012 (ISBN 978-5-397-02429-7).
- Ezekiel S. Medical Image Segmintation using multifractal analysis. Proc. of the App. Informatics, 2003, 378: 220-224.
- Lopes R., Betrouni N. Fractal and multifractal analysis: a review. Medical Image Analysis, 2009, 13(4): 634-649 ( ) DOI: 10.1016/j.media.2009.05.003
- Seuront L. Fractals and multifractals in ecology and aquatic science. UK, CRC Press, 2009.
- Takahashi T., Murata T., Narita K., Hamada T., Kosaka H., Omori M., Takahashi K., Kimura H., Yoshida H., Wada Y. Multifractal analysis of deep white matter microstructural changes on MRI in relation to early-stage atherosclerosis. NeuroImage, 2006, 32(3): 1158-1166 ( ) DOI: 10.1016/j.neuroimage.2006.04.218
- Tian Y.-C., Yu Z.-G., Fidge C. Multifractal nature of network induced time delay in networked control systems. Physics Letters A, 2007, 361(1): 103-107 ( ) DOI: 10.1016/j.physleta.2006.09.046
- Young I.M., Crawford J.W. Interactions and self-organization in the soil-microbe complex. Science, 2004, 304: 1634-1637 ( ) DOI: 10.1126/science.1097394
- Crawford J.W., Deacon L., Grinev D., Harris J.A., Ritz K., Singh B.K., Young I. Microbial diversity affects self-organization of the soil-microbe system with consequences for function. Journal the Royal Society Interface, 2012, 9: 1302-1310 ( ) DOI: 10.1098/rsif.2011.0679
- Воробьев Н.И., Свиридова О.В., Патыка Н.В., Думова В.А., Мазиров М.А., Круглов Ю.В. Фрактально-таксономический портрет микробного сообщества как биоиндикатор вида почвенных деструктивных процессов. Мат. Межд. конф. «Биодиагностика в экологической оценке почв и сопредельных сред». М., 2013: 38.
- Нетрусов А.И., Котова И.Б. Микробиология. М., 2006.
