Выведение промышленного сорта ели финской с использованием химических маркеров

Автор: Рогозин М.В.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Ботаника
Статья в выпуске: 3, 2016 года.
Бесплатный доступ
Программа выведения промышленного сорта разработана интеграцией трех направлений исследований: селекции, моделирования хода роста, изучения химического состава хвои. Сделан обзор результатов испытаний 525 потомств ели в густых и редких тест-культурах с выявлением 107 контрастных по типу роста семей. У семей изучено содержание 12 химических элементов (ХЭ) в хвое. Обнаружено, что конкурентный или светолюбивый тип развития семьи маркируют концентрации ХЭ в ней, и при отборе по некоторым параметрам их концентраций доля лучших семей повышается в 1.8-2 раза. С учетом этих данных, а также на основе связей между признаками и условиями формирования матерей и ростом их потомства, разработана программа селекции ели финской. Она включает отбор плюс-деревьев в насаждениях-аналогах плантационных культур, т.е. точно в таких же условиях, в каких будут выращивать их потомство. Эта мера повышает высоту потомства в среднем на 9.6%, а частоту лучших семей в 6 раз. Далее в возрасте 4-8 лет тестируют семьи от 1 тыс. деревьев и выделяют ~70 деревьев-кандидатов в сорт. Предложенная программа с учетом применения химического анализа хвои в 4-5 раз сокращает затраты и в 3 раза - сроки выведения сорта. Экспертная оценка стоимости программы на 10 лет оценивается в 40-45 млн руб. на один лесосеменной район.
Ель финская, потомство, типы роста, кандидаты в сорта, хвоя, химические элементы
Короткий адрес: https://sciup.org/147204774
IDR: 147204774 | УДК: 630
Breeding industrial cultivars Finnish spruce using chemical markers
The program is designed for industrial breeding varieties integration of the three research areas: selection, modeling the course of growth, the study of the chemical composition of the needles. A review of previous work of the author, including 525 test results in dense spruce progeny and rare test cultures, identifying 107 contrasting the type of family growth. These families been studied the content of 12 chemical elements (CE) in the needles. It has been found that the competitive family or photophilic family is determined (marked) concentration of chemical elements therein. In view of these data, and on the basis of relations between the features and conditions of formation of the mothers and the growth of their progeny, developed a breeding program of the Finnish spruce. It includes the selection of plus trees in stands of-analogues of plantation crops, i.e. exactly in the same conditions in which progeny will grow. This measure increases the average height of the progeny to 9.6% and the best families frequency 6 times. Then at the age of 4-8 years are tested family from 1 thousand trees and selected candidates ~70 grade. The proposed program, taking into account innovative approaches described in 4-5 times and reduces costs by 3 times - deadlines bred varieties. Expert assessment of the cost of the the program for 10 years, is estimated at 40-45 mln. rubles for one forest seed region.
Текст научной статьи Выведение промышленного сорта ели финской с использованием химических маркеров
Ель доминирует в хвойных лесах Пермского края и имеет важное ресурсное значение. В лесах края растет естественный гибрид ели сибирской и ели европейской. так называемая ель финская (Picea х fennica (Regel) Кот.) с ареалом от Урала до Финляндии* Изучалась ее морфометрия [Правдин, 1975; Попов, 2005: Коренев. 2008]. генетические особенности [Ми лютин и др., 2009; Нечаева и др.* 2012], а также возможности селекции [Рогозин, 2013]. Данная работа интегрирует результаты наших 30-летних исследований в виде краткого обзора; далее проведено более детальное освещение этапа селекции на основе химических маркеров.
Программа выведения сорта ели финской была разработана в результате объединения* казалось бы, довольно разных по тематике исследований:
(С Рогозин М. В., 2016
-
а) селекции ели для плантационного выращивания;
-
6) моделирования развития (хода роста) деревьев и древостоев;
-
в) изучения химического состава хвои у семей ели финской.
Результаты работ по этим направлениям позволили нам прийти к пониманию существа эволюции ели финской, точнее о самом ее начале, о ее первом шаге, о первом поколении, которое очень по-разному реагирует на конкурентные условия выращивания. Объем работ по первому7 направлению представлен результатами 20-летних испытаний 525 семей в двух тест-кульлурах потомством от плюсовых и обычных деревьев 12 ценопопуляций Пермского края. По второму’ направлению изучен ход роста ели на 349 пробных площадях и составлены 19 моделей развития древостоев ели в зависимости от начальной густоты сс естественных насаждений и культур. По третьемуг направлению изучено содержание 12 химических элементов в хвое у 107 семей с разным типом роста в густых и редких тест-культура х? а также химический состав почвы вблизи каждого из дерева, от которого брали хвою на анализ. Исследования обобщены в ряде наших работ [Рогозин, Разин. 2012. 2015; Рогозин. 2013].
Цель данной статьи - анализ данных автора по селекции ели финской в Пермском крае с определением вектора ее селекции на основе выяснения признаков и условий развития материнских деревьев, коррелирующих с ростом потомства.
Селекция ели финской для плантационного выращивания
Популяции ели финской в рассматриваемом регионе имеют отличия в морфометрии. Естественные популяции (Е) отличаются большим проявлением признаков ели сибирской, а искусственные популяции, т.с. культуры (К), тяготеют к ели европейской. Отличия приводят к различиям в тенденциях естественного отбора. Это проявляется в том, что быстрорастущие семьи в популяциях Е получаются в равной степени у матерей, имеющих мелкие, средние и крупные семена (отбор стабилизирующий), тогда как в популяциях К они получаются от матерей со средними и крупными семенами (отбор движущий). Близкие тренды обнаружены и по размеру шишек [Рогозин, Разин, 2012].
Однако внутри этих популяций лучшее потомство согласованно производят матери с близкими показателями морфометрии, а именно, со сбегом ствола от 1.2 до 1.3 см/м. В пределах указанных параметров число лучших семей было больше в 1.4-1.7 раза, т е. отбор по этому признаку’ оказался стабилизирующим. Данная особенность оказалась очень важна для понимания общего характера наследуемости быстроты роста. Корреляция между высотой матерей и высотой их семей в потомстве К (всего 224 семьи) в возрасте 21 год была равна г
= 0.186. Однако теоретические расчеты селекционного сдвига по ней оказались не корректны в силу того, что тренд связи был в виде параболы. В результате у самых высоких родителей высота потомства и доля лучших семей были такими же, что и у средних по высоте родителей, и только самые низкие деревья производили слабое потомство [Рогозин. Разин. 2012, с. 137-140].
Далее удалось выяснить, что на высоту потомства решающим образом влияет густота фитоценоза. где отбирали плюс-деревья. Но влияет опа в 'Зависимости от второго фактора, а именно, густоты дочерних тест-ку льтур: в густых культурах лучше растут потомки из густых ценозов, а в более редких - потомства из редких родительских ценозов. По данным измерения высот 4.3 тыс. растений 152 семей от пяти популяций при выращивании в разреженных тест-культурах потомство от родительских ценозов малой густоты оказалось выше на 11.0%, чем от ценозов с большей густотой, и выше на 9.6% высоты контроля. Противоположные различия были получены в густых тест-кулыурах Влияла на рост потомства не только густота всего насаждения, но и густота микроценоза. Так, у 79 плюс-деревьев были учтены все деревья-соседи на площадке радиусом 3.3 м и выделены группы родителей со слабой и сильной конкуренцией, имевшие в среднем по 7.1 и 10.9 шт. соседей соответственно. Оказалось, что при слабом давлении конкуренции на матерой высота их сомой в тест-ку л ыурах возрастала на 4.6%, а частота лучших семей - в 2 раза [Рогозин. Разин. 2012. с. 147-150].
Все это доказывает сильнейший характер влияния на потомство тех условий, в которых формировались их родители. Можно говорить даже о своеобразной эпигенетической «памяти» у потомства, проявляющей себя в соответствии с программой, которую прошли их родители, и густота насаждения влияет на сс проявление решающим образом. Поэтому последствия самого первого шага в селекции ели финской (т е. ее искусственной эволюции) могут быть эффективно использованы при выращивании ее искусственных лесов.
При сравнении наших данных с результатами других авторов по селекции основных лесообразующих видов можно сослаться на обзор А. И. Ви-дякина [Видякин. 2010], который отмечал, что в опытах разных авторов доля элитных семей изменяется от 0 до 90%. Причинами таких разных опенок были межпопуляционные различия, некорректные ВИДЫ контроля, отсутствие различий с контролем и несовершенство методик полевых опытов. Так. в республике Коми в одном крупном опыте среди потомства 129 плюс-деревьев превышали контроль лишь 3% семей, а все их потомство росло на 12.7% хуж*е контроля: в других опытах в возрасте 5-18 лет превышения семей 190 плюс-деревьев колебались от нуля до минус 3.6% [Тур- кин. 2007].
Анализ влияния параметров родительских и дочерних ценопопуляций как раз и позволил нам объяснить некоторые биологические Причины таких колебаний.
Моделирование развития (хода роста) деревьев и древостоев
Из изложенного выше получается, что конкуренция является тем камнем преткновения, который определяет успешность реализации буквально всех программ в лесной селекции, и необходимо учитывать законы, по которым развиваются древостои. Моделирование их развития позволило сформулировать общий закон развития одноярусных древостоев, «в одинаковых условиях местопроизрастания пределы развития древостоев определяет их начальная густота; при ее изменчивости примерно от 0.5-0.7 до 200 тыс. шт./га чем она больше, тем раньше древостой достигает своих пределов по показателям сомкнутости крон, суммам объемов крон, полноте, запасу, производительности, устойчивости и долговечности по сравнению с древостоями с меньшей начальной густотой; но чем она меньше, тем позднее древостой лидирует по указанным параметрам и дольше сохраняет лидерство по ним в сравнении с древостоями с большей начальной густотой» [Рогозин. Разин, 2015].
Закон проверен практикой [Рябоконь, 1990: Плантационное..., 2007], служит основой революционных изменений в способах выращивания леса и подобен закон)' предельной численности популяции и закон)' популяционного максимума в экологии [Одум, 1986; Реймерс, 1994]. В полном соответствии с ними в развитии древостоя имеется максимум текущего прироста, который В густых ценозах наступает уже в 15-20, а в редких - в 40-45 лет. Поэтому’ развитие сообщества древесных растений четко делится на период прогресса, те. возрастания прироста, и на период регресса, т е. его падения. Причем изменить модель развития на более прогрессивную рубками ухода можно только в фазе прогресса [Рогозин, Разин, 2015].
В соответствии с этим ключевым моментом меняется в целом вся парадигма лесоводства, а в лесной селекции - принципы испытаний, которые сокращаются путем опознания тренда линии развития. Причем достаточна будет только часть этой линии, например, до 10-15 лет. И здесь возникает проблема типов роста, но решается она не увеличением сроков испытаний, о чем постоянно сокрушаются селекционеры. Ее решение лежит в иной плоскости, а именно, в моделях развития древостоев. Пока можно использовать наши 15 моделей развития ели в естественных насаждениях и 4 модели - в ее культурах для условий. типичных для этой породы [Рогозин, Разин. 2015]. Суть моделей можно представить как веер из линий, где каждая линия имеет точку перегиба, и положение этой точки определяет начальная густота. В таком понимании пик развития древостоев ели с модальной начальной густотой в 3-5 тыс, шт./га приходится на 30-35 лет, и это известно, в общем, давно: однако пик этот подвижен. При высокой густоте он наступает в 15—20, а при малой — только в 45 лет. Поэтому редкие по начальной густоте модели наиболее продуктивны в 55-60 лет. и именно они должны быть образцом плантационных культур Особенно важно здесь то. что даже небольшие различия в густоте «на старте» сразу запускают механизм развития ценоза на много лег вперед. И густоту следует регулировать постоянно.
Если этого не делать, то буквально все показатели древостоя будут следовать в русле модели, имеющей определенную текущую густоту; и после пика развития лучшие потомства отстанут в росте, а на их место выйдут ранее отстававшие. Поэтому" тип роста потомства следует задавать изначально, т е. не изучать у потомства типы роста, затягивая испытания до возраста рубки, а управлять ими [Рогозин, Разин, 2015].
Выше мы рассмотрели наши итоги так называемой «коррелятивной» селекции, когда селекционеры ищут обходные пути И ВЫВОДЯТ сорта не напрямую. Этот подход в свете новых данных нам удалось улучшить и предложить новые правила для отбора плюс-дсрсвьсв, в частности, отбирать их в насаждениях-аналогах плантационных культур в возрасте 40-60 лет, со сбегом ствола 1.3-1.4 см/м. В результате при отборе популяций-аналогов высота их потомства в дочерних ценозах повышалась в среднем на 9.6%. а число лучших семей увеличивалось в 6 раз. Далее, в этих более редких родительских ценозах можно задействовать уровень микроценоза с отбором матерей при ослабленной конкуренции со стороны соседей. В результате в их потомстве доля лучших семей возрастает еще в 1.5 раза [Рогозин, Разин, 2012].
Следует отметить, что при столь разном характере развития деревьев в густых и редких фитоценозах должны быть различия в их физиологии. Следуя в рассуждениях далее, разная физиология должна приводить к различиям в химическом составе хвои, и здесь появились первые обнадеживающие результаты. Удалось доказать, что потомства естественных популяций ели с преобладанием признаков ели сибирской накапливают в хвое достоверно больше элементов Мп. TL Р. Си, Ba, Sr, Pb, а при снижении содержания органики в почве повышают ЗОЛЬНОСТЬ хвои, в отличие От изученных нами более «европейских» искусственных ценопо-пуляций, которые в своем потомстве снижают их концентрацию в хвое; при этом семьи последних обладают повышенной скоростью роста [Рогозин и др., 2014] В целом же оказалось, что химические элементы хвои у потомства связаны с быстротой роста семей намного сильнее, чем влияние высоты и сбега ствола, массы семени и размера шишек материнского дерева. при использовании которых удалось повысить частоту' лучших семей в 1*4-1.7 раза, а высоту потомства - на 3*0% [Рогозин, Разин, 2015].
Бос описанные выше влияния вполне могут быть объяснены эпигенетикой ИЛИ* ИНЫМИ словами* материнским эффектом и взаимодействием генотип-среда, влияющими на экспрессию генов и* таким образом* на фенотипы потомства. Это было известно генетикам давно, но только в последнее время появились методы, позволяющие изучать ИХ на полногеномном уровне [Крутовский, 2015].
Ниже мы приводим часть результатов исследований по химическому составу хвои ели.
Материал и методы исследования
Изучалось потомство ели финской из семян от свободного опыления* выращенное на двух участках тест-кулыур* заложенных в 1991 г. в Пермском крае. Первый участок создан в кв. 41 Ильинского лесничества. Почва среднесутлинистая, обедненная гумусом в результате сдвига по1геы при корчевке (биотоп С2). Посадка проведена по схеме 2.5* 1.0 м. Представлены потомства: из естественных популяций от 301 плюс-дерева и из культур от 224 деревьев, в том числе 152 плюсовые и 72 - обычные деревья. Контроль - семена от 30 стуча иных деревьев из всех популяций. По-садочный материал выращивали в теплице и далее в школе. Измерения проведены в 21 году 16 755 растении* в том числе 1 435 шт. в контроле*
Потомство этих же деревьев изучали также на втором участке тест-культур, заложенном на супеси в биотопе Б2* где посадки создавали 3-летними сеянцами на вырубке по раскорчеванным полосам тремя рядами по схеме 0.7*0 7 м* с расстоянием между полосами 8-20 м. Такая схема мало подходила для опыта, но позволила выяснить реакцию семей на конкуренцию. Этот участок, названный «густые культуры»* был заложен в кв. 5 Юго-Камского лесничества Оханского лесхоза на площади 8 га. Высажено 16.1 тыс* растений на 1 711 делянок, потомством 462 деревьев с тем же контролем. В 2010 г* в 23-летнем возрасте деревья были измерены и проведены осветления культур с вырубкой 15 м3/га древесины лиственных пород. Культуры сильно затенялись березой и осиной, которые превышали высоту7 ели на 1-5 м* т* е. на этом участке ель испытывала не только конкуренцию от растений своего вида в рядах* но и угнетение со стороны лиственных пород. Сохранилось только 34% растений, поэтому7 оставили семьи* где было 6 растений и более. Выборка на семью составила в среднем 13 шт* с ошибкой ее высоты ±8.4%. На Ильинском участке измеряли 29 растений на семью с ошибкой высоты ±5*4%* В итоге по два измерения высот оказались для 393 семьи и 21 вариант контроля*
В результате анализа этих пар семей были выделены 107 семей с разной реакцией на густоту выращивания. От этих деревьев на первом участке тест-культур выбирали по одному типичному дереву и брали образец хвои 1-3 лет из средней части кроны при объеме срезанных ветвей 1*0-1.5 л [Рогозин и др*, 2014]. Далее проводили атомноабсорбционный анализ хвои по 30 элементам на дифракционном спектрографе СТЭ-1 методом испарения из кратера угольного электрода [Методы.. 1989]
Предполагалось, что особенности почвы вблизи деревьев как-то могут повлиять на химический состав их хвои* Поэтому вблизи выбранных деревьев* в радиусе 1*0 м от ствола в трех местах делали прикопки до глубины 15-18 см и брали объединенный образец почвы на обычный почвеннохимический [ Арину шкина* 1962] и на атомноабсорбционный анализ [Методы*.., 1989], всего 111 образцов.
Результаты и их обсуждение
-
1, Влияние суммы элементов в хвое на рост семем
Из 216 семей от деревьев из естественных популяций и из 177 семей от деревьев из культур по данным об их высотах в 21 год в редких и в 23 года в густых тест-кулыурах были выделены разные типы роста семей. Если при увеличении густоты культур семья снижала высоту; то ее назвали «семья-светолюб». Если же она со увеличивала - это была «семья-кошуренгник»* те. плохо растущая в редких культурах, но хорошо - в густых. Всего из 393 семей, представленных в редких и густых тест-кулыурах, для анализа хвои выбрали 55 семей из естественных популяций и 52 семьи - из ценопопуляций культур, отличающихся контрастным типом роста. В результате атомно-абсорбционного анализа хвои были получены данные по 12 химическим элементам (ХЭ): Ni* Cr* Мп* V, Ti, Р, Сп, Zn, Ba, Sr* Zr* Pb. Сравнение их показало* что в потомстве естественных популяций (потомство Е) более активно накапливают ХЭ семьи-конкуренгники Сумма всех 12 элементов у них оказалась в 1*3 раза выше* чем у ссмсй-свстолюбов, и по 8 элементам из 12 повышение было достоверно. Однако в потомстве культур (потомство К) такого согласованного повышения ХЭ не наблюдалось. Всего лишь три элемента достоверно увеличили концентрации в семьях-конкурентниках: Ni. Мп и Zr — в 1.19-1.4 раза. Полученные данные о химическом составе хвои позволяют предполагать существование неких физиологических различий между их потомствами. Семьи-конкурентники из естественных популяций, видимо, формируют более морозоустойчивую хвою, накапливая в ней в 1.3 раза больше микроэлементов и повышая ее зольность в 1.08 раза. В потомствах культур, которые являются потомками более «европейской» ели финской, семьи делают это менее ак- тивно и по большинству микроэлементов их повышения недостоверны. Подробную интерпретацию этих данных мы приводили в предыдущей статье [Рогозин и др.. 2014].
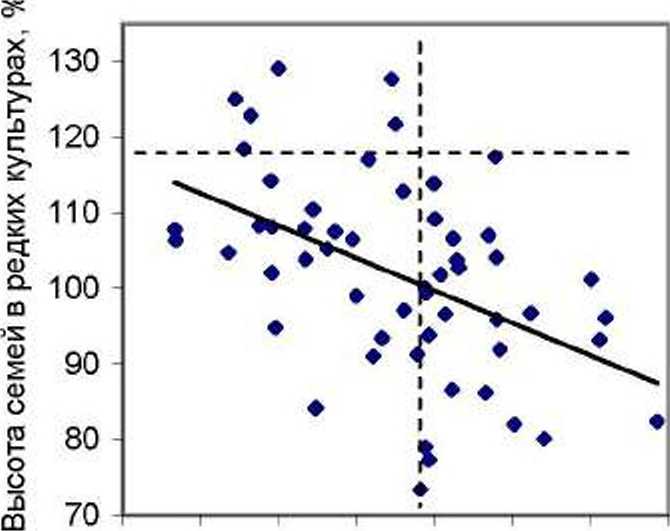
Далее мы проанализировали влияние суммы концентраций указанных выше 12 ХЭ на рост семей в разных по густоте тест-культу рах. Рассмотрим два поля корреляции, поместив их рядом: для случая редких культур слева, а для петых - справа (рис. 1).
0,6 0,8 1,0 1,2 1,4 1,6 1,8 2,0
Сумма ХЭ хвои потомства Е, г / кг
Рис. 1. Влияние суммы концентраций химических элементов в хвое ели на рост семей в редких (слева) и в густых тест-культу рах (справа) в потомстве естественных популяций
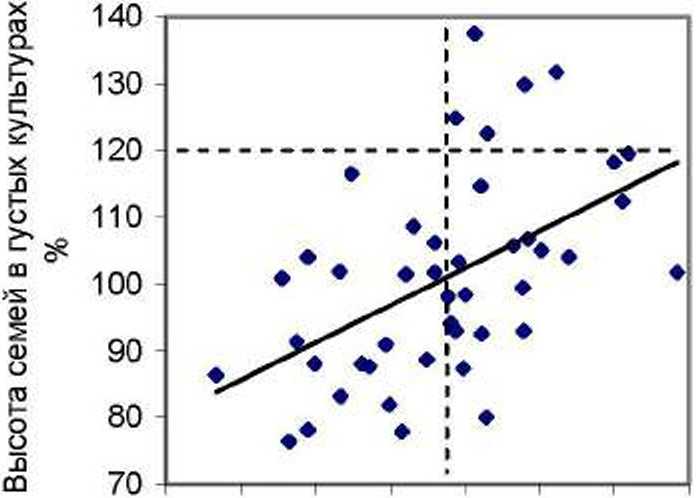
0,6 0,8 1,0 1,2 1,4 1,6 1,8 2,0 Сумма ХЭ хвои потомства Е, г / кг
На участке редких культур, где брали образцы хвои. т.е. на левом графике, связь между (уммой концентраций ХЭ и ростом семей отрицательна (г = -0.47±0.10). тогда как в густых культурах на правом графике (где хвою нс изучали) она стала положительной (г = 0.51±0.11). Различия между ними достоверны на самом высоком уровне (1 = 6.6 > t o.oi - 2.6).
Так как из густых культур хвою на химический анализ мы нс отбирали, то объяснить влияние ХЭ в ней на высоту, семей в том насаждении, где эта хвоя «нс работала», можно, по-вцдимому, тем. что уровень их содержания в хвое отражает, во-первых. особенности ее физиологи и. во-вторых, особенности генотипа. Поэтому можно полагать, что химические элементы в хвое ели в их суммарной концентрации и каждый по отдельности проявляют себя как некие химические признаки (хемопризнаки) дерева, увеличение концентраций которых приводит к снижению роста семей в редких культурах и, наоборот, увеличивает интенсивность их роста в густых, т е. идет проявление эпигенетических взаимодействий генотипа и среды. Отметим, что, в противовес всяким ожиданиям, в редких тест-культурах рост семей усиливался при снижении их концентраций. Почем}' это происходит - пока неясно и нужны дальнейшие исследования. Мы уже привыкли считать, что рост растений чаще всего улучшается при подкормке удобрениями. т е. растения испытывают некий «голод» по ряду' элементов. У деревьев ели получилось это не так, и они почему-то росли иногда лучше при их малом содержании в хвое.
Для оценки возможного практического эффекта можно имитировать отбор семей по сумме концентраций ХЭ. ориентируясь на значение 1.35 г/кг, и отбирать в редких тест-культурах семьи с малым, а в густых культурах - с большим содержанием суммы ХЭ в хвое (отделены вертикальной линией на графиках). После этого частота лучших семей (точки выше горизонтальной линии) составит на левом графике 5/27 = 0.185. что по отношению к исходной их частоте, равной 5/55 = 0.091. окажется выше в 2.03 раза. На правом графике исходная их частота была 5/44=0.114. а новая оказалась 5/24 = 0.208, что выше исходной в 1.82 раза.
Отобранные по указанным критериям семьи имеют среднюю высоту' в редких культурах 108.4±2.2 %, а в густых 107.2±3.1 %. и это неплохой результат. Если же учесть, что для получения образцов хвои потомства можно выращивать всего 3-4 года, а нс 20-30 лет. то при отсутствии взаимодействий «генотип-возраст» по изучаемым хемопризнакам вполне возможна ранняя диагностика: разумеется, данное взаимодействие следует выяснить далее путем повторных наблюдений в разном возрасте семей.
-
2. Влияние содержания химических элементов в почве на их содержание в хвое
Образец почвы для анализа формировали из трех прикопок, взятых с глубины 5-15 см на расстоянии 1.0 м от дерева. Содержание ХЭ в почве рассчитывали после ее сжигания, при котором выгорали все органические примеси. Всего на объекте проанализировано 111 образцов, для которых получены данные о содержании 17 химических элементов и их основные статистики (табл. 1 и 2).
В подавляющем числе образцов (85%) содержание органики находилось в пределах 3-8% при изменчивости показателя от 2.1 до 12.3% и его вариации 30.6% (табл. 3).
Данный показатель характеризует в какой-то мере количество гумуса в почве. Нас же интересовали элементы в минеральном остатке, и мы использовали обратный ему показатель, те. зольность, который по-лучали после сжигания в муфельной печи образца почвы в виде ее минерального остатка. На этот оста ток далее переводились полученные концентрации ХЭ. Например, при содержании органических включений 10% зольность почвы будет 100-10=90% или 0.90 от 1.0: соответственно, содержание ХЭ в почве делили на 0.90 и получали их концентрацию в минеральной части почвы
Таблица 1
Химические элементы в почве вблизи растений потомства естественных популяций ели финской*
|
№ се-мьи |
Микроэлементы в почве, мг/кг 17 элементов |
Орга-ника в почве, % |
Образцы хвои |
Высота семьи, % |
|||||||||||||||||
|
Зольность. % |
Сумма МЭ. мг'кг |
||||||||||||||||||||
|
Ki |
Со |
Сг |
Мп |
V |
Ti |
Su |
Си |
Zn |
РЬ |
Ва |
Sn |
Zr |
Ga |
Y |
Yb |
Nb |
|||||
|
/ |
49 |
20 |
176 |
683 |
146 |
6826 |
98 |
49 |
98 |
18 |
390 |
2,9 |
488 |
15 |
59 |
5.9 |
18 |
2.48 |
6.43 |
980 |
102.12 |
|
2 |
48 |
15 |
174 |
872 |
97 |
5810 |
9.7 |
48 |
87 |
15 |
291 |
3.9 |
387 |
15 |
48 |
4.8 |
15 |
3.16 |
6.23 |
1400 |
109.13 |
|
3 |
57 |
17 |
189 |
663 |
142 |
4734 |
9.5 |
47 |
95 |
14 |
284 |
3,8 |
284 |
17 |
38 |
3,8 |
9 |
5,32 |
5Л1 |
888 |
124,99 |
|
4 |
82 |
18 |
182 |
908 |
164 |
4542 |
9Л |
64 |
136 |
18 |
454 |
3.6 |
273 |
16 |
36 |
3,6 |
14 |
9,16 |
5.32 |
870 |
104,71 |
|
5 |
88 |
18 |
175 |
877 |
158 |
3510 |
8.8 |
53 |
132 |
18 |
351 |
3.5 |
263 |
16 |
35 |
3.5 |
13 |
12.26 |
5.77 |
1064 |
107.88 |
|
6 |
80 |
18 |
159 |
1329 |
177 |
4430 |
13,3 |
53 |
133 |
18 |
354 |
3,5 |
354 |
16 |
35 |
3,5 |
13 |
11,41 |
5,9 |
999 |
128,99 |
|
7 |
54 |
16 |
181 |
817 |
163 |
4537 |
8.2 |
64 |
91 |
16 |
272 |
3,6 |
181 |
14 |
36 |
3.6 |
9 |
9.26 |
6.2 |
1189 |
106.54 |
|
Я |
63 |
18 |
135 |
901 |
162 |
4506 |
8.1 |
63 |
162 |
16 |
270 |
2.7 |
270 |
18 |
36 |
3.6 |
9 |
9.88 |
5.43 |
980 |
114.25 |
|
9 |
38 |
14 |
143 |
856 |
143 |
4755 |
9.5 |
48 |
95 |
14 |
380 |
2,9 |
285 |
14 |
38 |
3,8 |
14 |
4,91 |
5.97 |
1144 |
107,51 |
|
10 |
38 |
14 |
142 |
663 |
142 |
4738 |
9.5 |
47 |
95 |
14 |
379 |
2,8 |
284 |
14 |
38 |
3,8 |
14 |
5,24 |
5.06 |
910 |
118,35 |
|
11 |
65 |
19 |
186 |
931 |
186 |
5583 |
14.0 |
47 |
186 |
19 |
279 |
3.7 |
372 |
17 |
37 |
3.7 |
9 |
6.95 |
5.65 |
1446 |
106.62 |
|
12 |
47 |
14 |
171 |
664 |
142 |
4740 |
9.5 |
38 |
95 |
14 |
379 |
2,8 |
190 |
17 |
38 |
3,8 |
14 |
5,20 |
5,4 |
1087 |
110,46 |
|
13 |
48 |
14 |
171 |
665 |
143 |
4751 |
9.5 |
48 |
86 |
14 |
380 |
2,9 |
285 |
17 |
38 |
3,8 |
14 |
4,99 |
5.24 |
1123 |
105,19 |
|
14 |
46 |
14 |
138 |
644 |
138 |
4598 |
8.3 |
55 |
83 |
9 |
368 |
2.8 |
276 |
14 |
28 |
2.8 |
8 |
8.04 |
6.83 |
1457 |
103.76 |
|
15 |
48 |
14 |
143 |
572 |
143 |
5724 |
9.5 |
48 |
86 |
10 |
382 |
2.9 |
382 |
14 |
38 |
3,8 |
14 |
4,60 |
5.54 |
1318 |
112,79 |
|
16 |
48 |
14 |
143 |
668 |
143 |
4768 |
9.5 |
38 |
86 |
14 |
381 |
2,9 |
286 |
14 |
29 |
2,9 |
14 |
4,64 |
6.38 |
1555 |
117,33 |
|
17 |
47 |
17 |
170 |
851 |
142 |
3780 |
8.5 |
47 |
85 |
14 |
378 |
2.8 |
378 |
14 |
38 |
3.8 |
14 |
5.49 |
6.65 |
1557 |
104.12 |
|
18 |
48 |
17 |
143 |
667 |
143 |
5714 |
8.6 |
48 |
86 |
14 |
381 |
3.8 |
286 |
14 |
38 |
3.8 |
19 |
4.77 |
5.71 |
1289 |
127.64 |
|
19 |
48 |
19 |
145 |
675 |
145 |
5788 |
8.7 |
58 |
96 |
10 |
386 |
3,9 |
386 |
17 |
39 |
3,9 |
17 |
3,53 |
6.05 |
1298 |
121,70 |
|
20 |
48 |
17 |
143 |
855 |
143 |
3801 |
86 |
57 |
95 |
17 |
285 |
2,9 |
380 |
17 |
29 |
2.9 |
14 |
4.97 |
4.26 |
981 |
108.18 |
|
21 |
47 |
14 |
142 |
665 |
95 |
3799 |
9.5 |
47 |
95 |
14 |
380 |
2.8 |
380 |
14 |
28 |
2.8 |
14 |
5.02 |
6.1 |
928 |
122.84 |
|
22 |
55 |
16 |
137 |
640 |
137 |
3657 |
6.4 |
46 |
91 |
14 |
366 |
2,7 |
274 |
18 |
27 |
2,7 |
14 |
8,58 |
5.97 |
735 |
106,35 |
|
23 |
63 |
18 |
134 |
626 |
134 |
4471 |
8,9 |
54 |
161 |
16 |
447 |
3,6 |
268 |
18 |
36 |
2.7 |
13 |
10.59 |
5.05 |
734 |
107.81 |
|
24 |
48 |
14 |
145 |
578 |
96 |
3855 |
8.7 |
48 |
87 |
14 |
289 |
2.9 |
289 |
17 |
39 |
2.9 |
14 |
3.62 |
|||
|
25 |
47 |
14 |
169 |
845 |
141 |
5633 |
14,1 |
56 |
94 |
14 |
376 |
2,8 |
282 |
17 |
28 |
2,8 |
17 |
6,11 |
6.69 |
1560 |
95,91 |
|
26 |
48 |
14 |
173 |
577 |
96 |
5773 |
9.6 |
38 |
96 |
29 |
385 |
2.9 |
481 |
14 |
29 |
3,8 |
14 |
3,79 |
|||
|
27 |
48 |
14 |
173 |
673 |
144 |
5765 |
14.4 |
48 |
144 |
19 |
480 |
2.9 |
384 |
17 |
38 |
3.8 |
17 |
3.92 |
6.84 |
1645 |
96.68 |
|
28 |
29 |
7 |
129 |
501 |
107 |
3575 |
7.2 |
36 |
72 |
14 |
358 |
2,1 |
358 |
13 |
29 |
2,9 |
13 |
4,96 |
6.54 |
1679 |
80,19 |
|
29 |
38 |
10 |
144 |
481 |
96 |
5773 |
14,4 |
58 |
144 |
17 |
481 |
2,9 |
385 |
17 |
29 |
3.8 |
19 |
3.79 |
6.89 |
1568 |
91.88 |
|
30 |
47 |
14 |
142 |
851 |
142 |
4727 |
14.2 |
47 |
95 |
17 |
378 |
2.8 |
378 |
17 |
28 |
3.8 |
14 |
5.47 |
5.62 |
1095 |
84.25 |
|
31 |
56 |
17 |
169 |
565 |
141 |
4707 |
14,1 |
66 |
141 |
28 |
377 |
2,8 |
377 |
19 |
28 |
2,8 |
14 |
5,87 |
7.29 |
1838 |
96,16 |
|
32 |
56 |
19 |
169 |
659 |
141 |
5650 |
14,1 |
47 |
141 |
19 |
377 |
2,8 |
377 |
19 |
28 |
3,8 |
19 |
5,84 |
5,8 |
1385 |
77.34 |
|
33 |
47 |
17 |
170 |
663 |
95 |
5680 |
14.2 |
57 |
85 |
19 |
473 |
1.9 |
284 |
17 |
38 |
4.7 |
19 |
5,33 |
|||
|
34 |
57 |
19 |
172 |
668 |
143 |
3817 |
14,3 |
57 |
95 |
17 |
382 |
3,8 |
382 |
17 |
29 |
3,8 |
17 |
4,58 |
7.88 |
1824 |
93,17 |
|
35 |
48 |
14 |
171 |
570 |
95 |
3802 |
14,3 |
67 |
143 |
17 |
285 |
2,9 |
380 |
17 |
29 |
3,8 |
17 |
4,95 |
6.13 |
1384 |
93,81 |
|
36 |
47 |
14 |
168 |
652 |
140 |
4657 |
14.0 |
56 |
140 |
19 |
466 |
3.7 |
373 |
17 |
37 |
4.7 |
14 |
6.87 |
6.04 |
1413 |
101.83 |
|
37 |
47 |
9 |
189 |
663 |
142 |
5680 |
14,2 |
57 |
95 |
17 |
473 |
2,8 |
379 |
17 |
28 |
3,8 |
17 |
5,34 |
5,7 |
1356 |
91,26 |
|
38 |
48 |
10 |
192 |
576 |
96 |
4799 |
9.6 |
48 |
86 |
17 |
384 |
2,9 |
480 |
17 |
38 |
3,8 |
19 |
4,04 |
|||
|
39 |
57 |
14 |
171 |
664 |
142 |
4744 |
14.2 |
57 |
142 |
17 |
474 |
3.8 |
380 |
17 |
38 |
3.8 |
19 |
5.12 |
7.24 |
1801 |
101.23 |
|
40 |
56 |
19 |
186 |
930 |
139 |
4648 |
13.9 |
37 |
84 |
14 |
465 |
3.7 |
372 |
17 |
37 |
3,7 |
14 |
7,05 |
5.57 |
1375 |
78.95 |
|
41 |
47 |
14 |
187 |
844 |
141 |
5624 |
8.4 |
47 |
94 |
19 |
375 |
1,9 |
375 |
17 |
28 |
3,7 |
14 |
6,26 |
5.36 |
1320 |
97Д2 |
|
42 |
47 |
17 |
171 |
664 |
142 |
5689 |
14.2 |
47 |
95 |
19 |
379 |
3.8 |
379 |
14 |
38 |
4,7 |
17 |
5.19 |
5.22 |
1362 |
73.36 |
|
43 |
57 |
17 |
189 |
850 |
142 |
5669 |
9.4 |
66 |
94 |
19 |
472 |
2.8 |
378 |
17 |
28 |
3.8 |
17 |
5.51 |
5.28 |
1377 |
99.38 |
|
44 |
48 |
14 |
172 |
669 |
143 |
5731 |
9.6 |
48 |
96 |
29 |
478 |
2,9 |
382 |
17 |
29 |
3,8 |
19 |
4,48 |
6Л2 |
1443 |
86,54 |
|
45 |
48 |
17 |
194 |
873 |
145 |
5817 |
9,7 |
58 |
97 |
29 |
485 |
3,9 |
388 |
19 |
29 |
3.9 |
17 |
3.05 |
|||
|
46 |
48 |
14 |
174 |
483 |
145 |
5798 |
8.7 |
48 |
97 |
17 |
387 |
2.9 |
387 |
17 |
29 |
3.9 |
17 |
3,37 |
|||
|
47 |
48 |
14 |
171 |
570 |
143 |
5701 |
14,3 |
48 |
95 |
38 |
380 |
2,9 |
380 |
17 |
48 |
4,8 |
17 |
4,98 |
7.58 |
1970 |
82,39 |
|
48 |
56 |
17 |
188 |
563 |
141 |
4693 |
9,4 |
56 |
141 |
28 |
563 |
2,8 |
375 |
19 |
47 |
4,7 |
14 |
6.1 5 |
6.8 |
1605 |
82.06 |
|
49 |
48 |
19 |
173 |
865 |
86 |
3843 |
9.6 |
38 |
67 |
8 |
173 |
1.9 |
288 |
14 |
29 |
2,9 |
14 |
3.92 |
6.19 |
1263 |
93.44 |
|
50 |
38 |
17 |
95 |
856 |
95 |
4754 |
14,3 |
48 |
67 |
14 |
171 |
1,9 |
171 |
14 |
29 |
2,9 |
14 |
4,93 |
6.09 |
1242 |
90,99 |
|
51 |
39 |
15 |
97 |
678 |
87 |
.3874 |
8,7 |
48 |
68 |
15 |
174 |
1,9 |
174 |
15 |
29 |
2.9 |
17 |
3.14 |
|||
|
52 |
48 |
17 |
96 |
675 |
96 |
5789 |
9.6 |
48 |
87 |
14 |
289 |
2.9 |
289 |
14 |
29 |
2,9 |
17 |
3.51 |
6.41 |
1531 |
86.28 |
|
53 |
48 |
14 |
145 |
579 |
96 |
4821 |
9.6 |
48 |
67 |
10 |
193 |
2,9 |
289 |
14 |
29 |
2,9 |
17 |
3,58 |
|||
|
Окончание табл. 1 |
|||||||||||||||||||||
|
№ се-MLH |
Микроэлементы в почве, мг/кг 17 элементов |
Орга-ника в почве, % |
Образцы хвои |
Высота семьи. % |
|||||||||||||||||
|
Зольность, % |
Сумма МЭ. мг'кг |
||||||||||||||||||||
|
М |
Со |
СТ |
Мп |
\ |
Ti |
Sc |
Си |
Zn |
Pb |
Ва |
Sn |
Zi |
Ga |
Y |
Yb |
Nb |
|||||
|
54 |
49 |
15 |
88 |
681 |
88 |
4863 |
9.7 |
39 |
58 |
10 |
292 |
1,9 |
389 |
15 |
29 |
2.9 |
15 |
2.74 |
- |
||
|
55 |
49 |
15 |
146 |
582 |
146 |
5825 |
8.7 |
49 |
68 |
15 |
291 |
3,9 |
388 |
10 |
39 |
3,9 |
15 |
2,92 |
5,91 |
1537 |
106,99 |
|
56 |
57 |
19 |
144 |
862 |
144 |
5747 |
9.6 |
57 |
96 |
17 |
383 |
3,8 |
383 |
17 |
29 |
2,9 |
17 |
4,22 |
6.99 |
1397 |
113,83 |
|
Статистики для почвы, хвои и семей потомства естественных популяций |
|||||||||||||||||||||
|
п |
56 |
56 |
56 |
56 |
56 |
56 |
56 |
56 |
56 |
56 |
56 |
56 |
56 |
56 |
56 |
56 |
56 |
56 |
47 |
47 |
47 |
|
51,0 |
15.6 |
158,6 |
716,0 |
132,2 |
49475 |
ЮЛ |
зол |
102,2 |
17,0 |
368 |
3,02 |
338.7 |
16,1 |
34,1 |
3.6 |
15,1 |
5,45 |
6,05 |
1317.1 |
101,58 |
|
|
6 |
ЮЛ |
2,6 |
25.6 |
150,3 |
24,6 |
^92,8 |
2,5 |
7,6 |
28,1 |
5,5 |
84,6 |
0,58 |
71,0 |
1,8 |
6,5 |
0.7 |
2.7 |
2,17 |
0,73 |
295,50 |
13.76 |
|
V,% |
19,8 |
167 |
16,1 |
21,0 |
18,6 |
16,0 |
23,3 |
14,9 |
27,5 |
32,3 |
23,0 |
19,3 |
21,0 |
11,4 |
18,9 |
18 |
17,9 |
39,8 |
12,1 |
22,4 |
13,6 |
|
* Данные по химическому составу почвы получены к.б.н. Л.В. Кувшинской в 2013 г |
|||||||||||||||||||||
Таблица 2
Химические элементы в почве вблизи растений из потомства культур ели финской*
|
№ се- МКН |
Микроэлементы в почве, мг/кг 17 элементов |
Орга-ника в почве, % |
Образцы хвои |
Высота семьи, % |
|||||||||||||||||
|
Зольность, |
Сумма МЭ. mj/kj |
||||||||||||||||||||
|
Ni |
Со |
Сг |
Мл |
V |
Ti |
Sc |
Си |
Zn |
Pb |
Ba |
Sn |
Zr |
Ga |
Y |
Yb |
Nb |
|||||
|
57 |
48 |
17 |
143 |
668 |
95 |
3818 |
8,6 |
48 |
67 |
14 |
286 |
2,9 |
286 |
17 |
29 |
2,9 |
14 |
4,56 |
. |
||
|
58 |
38 |
14 |
141 |
660 |
85 |
3769 |
8,5 |
38 |
85 |
9 |
283 |
L9 |
188 |
14 |
28 |
2,8 |
14 |
5,78 |
6.65 |
1335 |
110.04 |
|
59 |
38 |
17 |
96 |
671 |
86 |
3836 |
9,6 |
48 |
86 |
14 |
144 |
1,9 |
192 |
14 |
29 |
2,9 |
14 |
4Л0 |
6 |
1448 |
116,71 |
|
60 |
29 |
15 |
97 |
581 |
87 |
5809 |
9,7 |
48 |
68 |
15 |
174 |
1,9 |
194 |
17 |
29 |
2,9 |
10 |
3,19 |
5Л7 |
1123 |
124,62 |
|
61 |
47 |
14 |
141 |
565 |
141 |
4708 |
9,4 |
66 |
141 |
14 |
282 |
2,8 |
377 |
17 |
28 |
2,8 |
9 |
5.84 |
5,04 |
1122 |
128,37 |
|
62 |
48 |
17 |
143 |
669 |
96 |
4782 |
9,6 |
67 |
96 |
14 |
287 |
2,9 |
287 |
17 |
29 |
2,9 |
14 |
4,36 |
4,79 |
1112 |
126,46 |
|
63 |
29 |
17 |
97 |
679 |
87 |
4850 |
9,7 |
48 |
58 |
9 |
145 |
1,9 |
194 |
15 |
39 |
2,9 |
10 |
3,01 |
4,7 |
1085 |
146,16 |
|
64 |
39 |
14 |
145 |
578 |
87 |
5783 |
14,5 |
39 |
58 |
14 |
145 |
2,9 |
289 |
14 |
39 |
2.9 |
14 |
3,61 |
5.08 |
1186 |
120.56 |
|
65 |
46 |
18 |
165 |
826 |
138 |
5509 |
9,2 |
55 |
92 |
17 |
367 |
2,8 |
275 |
14 |
28 |
2,8 |
9 |
8Л9 |
6,48 |
880 |
107,81 |
|
66 |
48 |
19 |
173 |
578 |
96 |
5776 |
9,6 |
58 |
67 |
14 |
385 |
2,9 |
289 |
17 |
39 |
2,9 |
10 |
3,74 |
4,64 |
908 |
143,27 |
|
67 |
47 |
14 |
142 |
664 |
95 |
3796 |
8,5 |
47 |
57 |
9 |
190 |
1,9 |
171 |
17 |
38 |
2,8 |
9 |
5,09 |
4,76 |
1079 |
133,78 |
|
68 |
38 |
9 |
142 |
851 |
95 |
5671 |
14,2 |
66 |
95 |
17 |
378 |
2,8 |
473 |
17 |
38 |
4,7 |
17 |
5,48 |
- |
- |
- |
|
69 |
38 |
9 |
144 |
576 |
96 |
5758 |
14,4 |
58 |
96 |
17 |
384 |
3,8 |
384 |
17 |
38 |
3,8 |
19 |
4,04 |
5,92 |
1283 |
99,49 |
|
70 |
47 |
14 |
142 |
662 |
95 |
5673 |
14,2 |
57 |
142 |
19 |
378 |
2,8 |
284 |
17 |
38 |
4,7 |
19 |
5,45 |
- |
||
|
71 |
47 |
14 |
142 |
661 |
94 |
4723 |
9,4 |
57 |
142 |
19 |
378 |
1,9 |
283 |
17 |
28 |
3,8 |
19 |
5,54 |
6,26 |
1377 |
102,23 |
|
72 |
47 |
14 |
142 |
570 |
95 |
5698 |
14.2 |
66 |
95 |
19 |
475 |
3.8 |
380 |
17 |
28 |
4,7 |
19 |
5,04 |
5.57 |
1107 |
82.40 |
|
73 |
48 |
10 |
143 |
573 |
95 |
4771 |
14,3 |
57 |
95 |
19 |
286 |
3,8 |
382 |
17 |
38 |
3,8 |
17 |
4,58 |
5,37 |
1203 |
97,93 |
|
74 |
48 |
14 |
143 |
668 |
95 |
4772 |
9,5 |
48 |
95 |
14 |
382 |
2,9 |
382 |
19 |
38 |
3,8 |
17 |
4,57 |
6,57 |
1555 |
89,56 |
|
75 |
48 |
14 |
143 |
667 |
95 |
4765 |
9.5 |
57 |
95 |
19 |
381 |
3,8 |
381 |
17 |
29 |
3.8 |
14 |
4,70 |
6,05 |
1449 |
86,10 |
|
76 |
48 |
14 |
143 |
669 |
96 |
4777 |
9,6 |
67 |
96 |
17 |
382 |
1,9 |
287 |
14 |
29 |
3,8 |
17 |
4,46 |
5,07 |
1194 |
103,97 |
|
77 |
48 |
14 |
172 |
859 |
143 |
5729 |
9,5 |
67 |
143 |
19 |
382 |
2,9 |
382 |
19 |
38 |
3,8 |
17 |
4,51 |
5.77 |
1292 |
94.30 |
|
78 |
48 |
14 |
171 |
667 |
143 |
5716 |
9,5 |
57 |
143 |
14 |
476 |
3,8 |
381 |
17 |
29 |
3,8 |
19 |
4,73 |
5.4 |
1224 |
101,61 |
|
79 |
49 |
20 |
176 |
683 |
146 |
6826 |
9,8 |
49 |
98 |
18 |
390 |
2,9 |
488 |
15 |
59 |
5,9 |
18 |
2,48 |
5,33 |
1076 |
116,21 |
|
80 |
47 |
17 |
94 |
659 |
85 |
4705 |
9,4 |
47 |
56 |
9 |
282 |
1,9 |
376 |
14 |
28 |
2,8 |
9 |
5,90 |
|||
|
81 |
47 |
19 |
93 |
653 |
93 |
4666 |
14,0 |
47 |
93 |
14 |
280 |
2,8 |
373 |
14 |
28 |
2,8 |
9 |
6,68 |
- |
- |
|
|
82 |
56 |
19 |
139 |
836 |
139 |
4645 |
9,3 |
56 |
139 |
17 |
372 |
2,8 |
279 |
17 |
37 |
3,7 |
9 |
7,11 |
- |
- |
- |
|
83 |
48 |
17 |
96 |
579 |
96 |
4823 |
9,6 |
48 |
68 |
14 |
289 |
2,9 |
289 |
14 |
39 |
3,9 |
10 |
3.55 |
3,76 |
904 |
106,20 |
|
84 |
56 |
19 |
141 |
658 |
141 |
4703 |
9,4 |
47 |
141 |
19 |
376 |
3,8 |
376 |
14 |
38 |
3,8 |
17 |
5,95 |
- |
- |
113,85 |
|
85 |
48 |
19 |
143 |
669 |
143 |
5735 |
9,6 |
48 |
96 |
17 |
478 |
3,8 |
478 |
14 |
38 |
2,9 |
17 |
4,41 |
4,19 |
930 |
117,15 |
|
86 |
56 |
19 |
140 |
562 |
140 |
4682 |
9,4 |
47 |
94 |
14 |
281 |
2,8 |
375 |
14 |
28 |
2.8 |
8 |
6,36 |
|||
|
87 |
48 |
19 |
145 |
677 |
97 |
5807 |
14,5 |
58 |
97 |
17 |
387 |
3,9 |
484 |
15 |
39 |
3,9 |
17 |
3,22 |
4,31 |
996 |
112,34 |
|
88 |
39 |
17 |
97 |
483 |
87 |
4831 |
9,7 |
39 |
58 |
17 |
174 |
2,9 |
290 |
14 |
29 |
2,9 |
9 |
3,38 |
5,63 |
1159 |
98,79 |
|
89 |
57 |
19 |
171 |
854 |
171 |
4747 |
8,5 |
57 |
95 |
19 |
380 |
3,8 |
380 |
17 |
38 |
3,8 |
19 |
5,06 |
4,97 |
1233 |
106,56 |
|
90 |
48 |
17 |
144 |
671 |
144 |
4793 |
9,6 |
58 |
67 |
17 |
383 |
3,8 |
479 |
10 |
38 |
3,8 |
19 |
4Л5 |
4,47 |
1030 |
102,52 |
|
91 |
58 |
17 |
192 |
673 |
144 |
4808 |
9.6 |
67 |
96 |
19 |
385 |
4,8 |
385 |
14 |
29 |
1,9 |
17 |
3,84 |
5,7 |
1302 |
95,07 |
|
92 |
68 |
17 |
194 |
680 |
146 |
4858 |
14,6 |
68 |
97 |
29 |
486 |
4,9 |
389 |
15 |
39 |
3,9 |
19 |
2,84 |
|||
|
93 |
90 |
27 |
179 |
806 |
179 |
4479 |
16,1 |
81 |
179 |
27 |
448 |
6,3 |
269 |
18 |
36 |
3,6 |
18 |
10,42 |
5,72 |
1050 |
129,37 |
|
94 |
48 |
17 |
144 |
864 |
144 |
5758 |
5,8 |
58 |
96 |
19 |
480 |
3.8 |
288 |
14 |
29 |
2,9 |
10 |
4.03 |
6.42 |
1563 |
111.07 |
|
95 |
57 |
19 |
172 |
668 |
143 |
3817 |
14,3 |
57 |
95 |
17 |
382 |
3,8 |
382 |
17 |
29 |
3,8 |
17 |
4,58 |
|||
|
96 |
49 |
18 |
176 |
683 |
98 |
5858 |
9,8 |
39 |
88 |
18 |
488 |
2,9 |
488 |
10 |
39 |
3,9 |
15 |
2.36 |
4,68 |
1219 |
87,85 |
|
97 |
48 |
14 |
171 |
570 |
95 |
3802 |
14,3 |
67 |
143 |
17 |
285 |
2,9 |
380 |
17 |
29 |
3.8 |
17 |
4.95 |
5,48 |
1291 |
85,57 |
|
98 |
48 |
17 |
144 |
862 |
144 |
5749 |
8,6 |
57 |
96 |
14 |
479 |
3,8 |
287 |
14 |
29 |
2,9 |
17 |
4,18 |
7,03 |
1649 |
90,82 |
|
99 |
58 |
19 |
173 |
962 |
144 |
4808 |
9,6 |
58 |
144 |
19 |
481 |
4,8 |
288 |
17 |
29 |
2,9 |
17 |
3,85 |
4.08 |
936 |
97.15 |
Окончание табл. 2
|
№ се-мьм |
Микроэлементы в почве, мг/кг 17 элементов |
Орга-ника в почве. % |
Образцы хвои |
Высота семьи, % |
|||||||||||||||||
|
Зольность, % |
Сумма МЭ, мг/кг |
||||||||||||||||||||
|
Ni |
Со |
Сг |
Мп |
V |
Ti |
Sc |
Си |
7,п |
Ph |
Ra |
Sn |
Zr |
Ga |
Y |
Yb |
Nb |
|||||
|
100 |
56 |
19 |
169 |
938 |
141 |
5628 |
4,7 |
47 |
94 |
19 |
375 |
3,8 |
375 |
17 |
28 |
1,9 |
17 |
6.20 |
- |
||
|
101 |
48 |
14 |
173 |
673 |
144 |
4810 |
8,7 |
48 |
87 |
14 |
385 |
2,9 |
385 |
14 |
29 |
2,9 |
14 |
3,81 |
5,17 |
1166 |
92,53 |
|
102 |
47 |
19 |
171 |
854 |
142 |
5690 |
9,5 |
57 |
95 |
14 |
474 |
3,8 |
379 |
14 |
38 |
3,8 |
17 |
5,16 |
5,25 |
1284 |
95,84 |
|
103 |
57 |
19 |
171 |
854 |
142 |
4746 |
9,5 |
57 |
95 |
19 |
475 |
3,8 |
285 |
14 |
28 |
2,8 |
9 |
5.08 |
4,04 |
909 |
88,72 |
|
104 |
57 |
19 |
171 |
855 |
143 |
5701 |
14,3 |
57 |
95 |
19 |
475 |
3,8 |
380 |
14 |
29 |
2,9 |
10 |
4,98 |
- |
- |
|
|
105 |
58 |
19 |
174 |
675 |
145 |
6751 |
9,6 |
58 |
96 |
19 |
579 |
4,8 |
482 |
17 |
39 |
3,9 |
14 |
3,56 |
4,51 |
993 |
89,63 |
|
106 |
48 |
10 |
192 |
576 |
96 |
4799 |
9,6 |
48 |
86 |
17 |
384 |
2,9 |
480 |
17 |
38 |
3,8 |
19 |
4.04 |
5,13 |
1208 |
98,68 |
|
107 |
48 |
19 |
145 |
675 |
145 |
5783 |
9,6 |
48 |
96 |
14 |
386 |
.3,9 |
386 |
14 |
29 |
1,9 |
17 |
3,61 |
5,43 |
1219 |
94,97 |
|
108 |
48 |
14 |
96 |
575 |
96 |
3835 |
8,6 |
48 |
86 |
9 |
288 |
2,9 |
288 |
10 |
19 |
1,9 |
17 |
4,13 |
5,53 |
1342 |
94,75 |
|
109 |
47 |
17 |
142 |
664 |
142 |
4744 |
6,6 |
47 |
85 |
14 |
285 |
3,8 |
285 |
14 |
38 |
3,8 |
14 |
5.13 |
5,55 |
1270 |
83,17 |
|
110 |
47 |
17 |
171 |
664 |
142 |
5689 |
14,2 |
47 |
95 |
19 |
379 |
3,8 |
379 |
14 |
38 |
4.7 |
17 |
5.19 |
5.9 |
1286 |
79,16 |
|
Ш |
47 |
9 |
189 |
663 |
142 |
5680 |
14.2 |
57 |
95 |
17 |
473 |
2,8 |
379 |
17 |
28 |
3,8 |
17 |
5,34 |
5,3 |
1284 |
94,84 |
|
для поивы. хвои и семей потомства культур |
|||||||||||||||||||||
|
п |
55 |
55 |
55 |
55 |
55 |
55 |
55 |
55 |
55 |
55 |
55 |
55 |
55 |
55 |
55 |
55 |
55 |
43 |
43 |
44 |
|
|
X |
49,1 |
17,1 |
147,4 |
6792 |
118,4 |
4987,4 |
11,3 |
54,4 |
96,6 |
17,2 |
356,6 |
4,20 |
341.1 |
16.2 |
33,7 |
4,32 |
15,6 |
5,6 |
6,2 |
1166,0 |
103,2 |
|
6 |
8.8,4 |
3.24 |
27.52 |
1082 |
27.1 |
730.2 |
2.57 |
8,77 |
26.6 |
3.72 |
99.98 |
0.89 |
82,64 |
2,08 |
6.19 |
0.78 |
3,69 |
1,38 |
0,75 |
184.25 |
16.32 |
|
v;% |
18,1 |
18,9 |
18,7 |
15,9 |
22.9 |
14,6 |
22,7 |
16,1 |
27,5 |
21,6 |
28,0 |
21,3 |
24,2 |
12.9 |
18,4 |
18,0 |
24,5 |
12.2 |
15,8 |
15,8 |
|
* Данные по химическом) составу почвы получены к.б.н. Л,В. Кувшинской в 2013 г.
Органика в образцах почвы в непитательных культурах слн
Таблица 3
|
Органические включения, % |
2.1-3.0 |
3.1-4.0 |
4.1-5.0 |
5.1-6.0 |
6.1-8.0 |
8.1-10.0 |
10.1-12.3 |
Итого |
|
Количество проб, шт. |
8 |
26 |
31 |
26 |
11 |
5 |
4 |
111 |
|
Количество проб, % |
7.2 |
23 4 |
27.9 |
23.4 |
9.9 |
4.5 |
3.6 |
100.0 |
Органика в почве - наиболее просто определяемый показатель, и было интересно выяснить, как она влияет на содержание ХЭ в хвое, а также на ее зольность в популяциях.
Для этого общая выборка семей была разделена на две части: одна для потомства из естественных популяций (56 образцов) и другая - для потомства из культур (55 образцов). Общая ведомость содержания микроэлементов приведена в табл. 1 и 2.
По этим данным мы выяснили, отличается ли почва вблизи деревьев из потомства культур (потомство К) от почвы вблизи потомства из естественных популяций (потомство Е). В конце упомянутых таблиц помещены статистики и их отношения, сравнение которых показало, что в целом отличия малы. Среднее содержание ХЭ в почве вблизи деревьев из потомства К, по отношению к их содержанию у потомства Е колебалось от 0.94 до 1.08 и в целом оказалось несколько ниже (0.987), а отношения вариаций ХЭ колебались от 0.75 до 1.41 и в целом оказались несколько выше (1.088). Различия оказались несущественны и мы приводим их, чтобы показать практически одинаковый м икроэлементный состав почвы под обоими потомствами.
Далее, из 111 образцов почвы было проведено сравнение химического состава 90 образцов почвы и 90 образцов хвои, в т.ч. 47 из потомства естественных популяций и 43 - из культур. Оставшиеся образцы почвы (21 шт.) не имели соответствующей пары по хвое, так как на ее анализ не хватило финансовых средств (эти образцы хвои пока не проанализированы и представляют группу семей-универсалов с быстрым ростом как в редких, так и в густых испытательных культурах).
Оказалось, что влияние по существу одинаковой по составу ХЭ почвы на химический состав хвои в сравниваемых потомствах были различны. Повышение органики в почве приводило к тому; что в потомстве естественных популяций зольность хвои снижалась, а в потомстве культур, наоборот, происходило ее повышение (рис. 2).
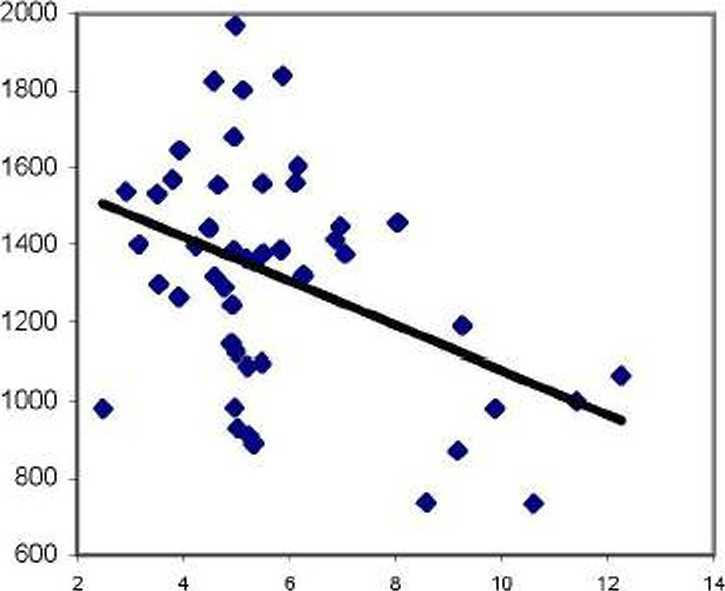
Разные тенденции «утяжеления» хвои зольным остатком выражаются в достоверных различиях (t^ = 3.0 > to,05= 2.0) между корреляциями для левого (г = -0.250 ± 0.137) и для правого полей (г = 0.334 ±0.135).
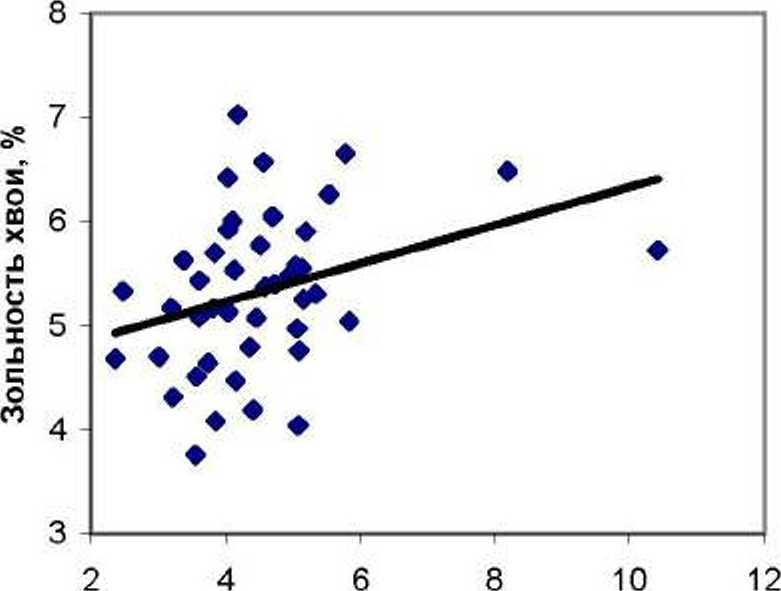
Влияние органики в почве на сумму ХЭ в хвое оказалось несколько иным: в потомстве естественных популяций при увеличении органики в почве сумма ХЭ в хвое снижалась (г = - 0.42±0.12), тогда как в потомстве культур этот показатель имел недостоверную связь с органикой (г = — 0.05 ± 0.15) и оставался практически на одном уровне, при достоверном различии между7 этими коэффициентами При 1ф = 1.91> to.LO= 1 66 (рис. 3).
Несмотря на достоверные различия между рассмотренными четырьмя корреляциями, можно критически отнестись К Трендам ВЛИЯНИЯ Органики на зольность и на содержание ХЭ в хвое, так как для органики кластер значений 8-12% представлен малым количеством образцов: 7 шт. для попу ля- ций Е и 2 шт. для популяций К (рис. 2). Кроме того. различия между последней парой корреляций были достоверны лишь при пониженном уровне статистических (при t0 ю = 1.66).
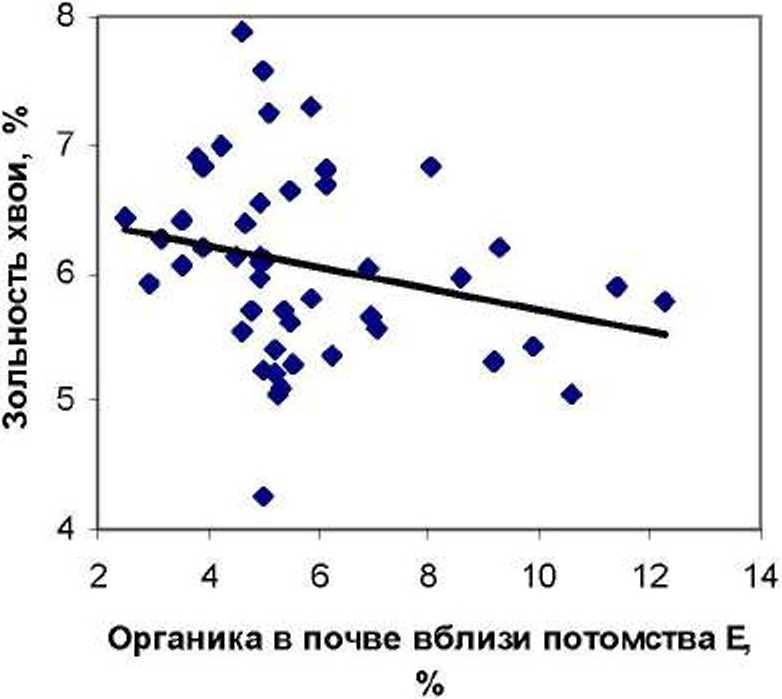
Рис. 2. Влияние содержания органики в почве вблизи деревьев на зольность хвои ели в потомстве естественных популяций (слева) и в потомстве культур (справа)
Органика в почве вблизи потомства К, %
Органика в почве вблизи потомства Е, %
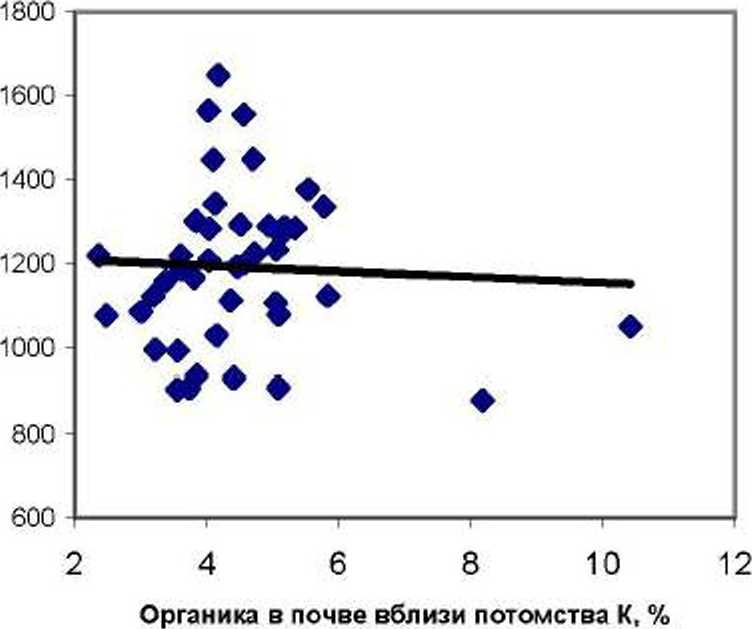
Рис. 3. Влияние содержания органики в почве вблизи деревьев на суммарное содержание микроэлементов в хвое в потомстве естественных популяций (слева) и в потомстве культур (справа)
Поэтому*, не удовлетворившись этими четырьмя графиками, мы проанализировали еще и влияние суммы ХЭ в почве на сумму ХЭ в хвое, предполагая при этом, что с увеличением суммарного содержания ХЭ в почве их содержание в хвое также должно увели чиваться.
И вновь тренды связей оказались различны. В потомстве Е корреляция суммы ХЭ в почве с суммой ХЭ в хвое оказалась положительной и недостоверной (г = 0.215 ± 0.139). а в потомстве К наоборот. она оказалась отрицательна (г = - 0.145 ± 0.149). Несмотря на недостоверность обеих корреляций, их разная направленность приводит к достоверным различиям между ними при Ц= 1.76 > t0 ю= 1.66. Если же выборки потомств Е и К объединить. то связь окажется в целом близка к нулю.
Таким образом, зольность и химический состав хвои дерева варьируют во многом самостоятельно, и они почти нс связаны с вариацией химического состава почвы вблизи растения, имеющей в целом по участку выравненный агрофон.
Коль скоро разнообразие почвы по органике и по сумме ХЭ вблизи деревьев слабо и не всегда, но все-таки влияет на сумму, и. вероятно, на состав ХЭ в хвое, то влияние это нужно как-то убрать, чтобы прояснить главные для нас моменты в селекции. а именно: как отдельные химические элементы в хвое, их сумма, а также зольность хвои влияют на рост отдельных семей и потомства в целом.
Для этого мы поступили самым простым образом - показатели каждого ХЭ в хвое дерева разделили на его значение в почве вблизи этого дерева. Для этого значение ХЭ в почве, полученное вблизи конкретного дерева, перевели в относительные величины и выразили их в долях единицы от среднего содержания элемента в выборке. Далее эти относительные величины ХЭ в хвое и их связь с рос- том семей из потомства Е и К анализировали повторно.
Однако полученные по этим относительным значениям корреляции отличались от рассчитанных ранее несущественно. Не было обнаружено и достоверных корреляций между содержанием отдельных ХЭ в почве и в хвое, коррелируемых по отдельности. Убедившись окончательно, что химический состав субстрата вблизи деревьев не влияет на состав ХЭ в хвое, мы пришли к заключению, что изменчивость содержания химических элементов в хвое у отдельных семей практически не зависит от их содержания в почве.
-
3. Влияние содержания химических элементов в почве па рост потомства ели
Выше отмечалось. что почва вблизи деревьев не имела достоверных отличий по массе органики и по изучаемым элементам, т. е. почву под деревьями по этим характеристикам можно считать одинаковой. Эти две характеристики почвы (масса органики и сумма 17 элементов) изменчивы, и это может как-то повлиять на развитие деревьев, хотя на содержание ХЭ в их хвое почва практически не повлияла.
Органика в почве варьирует от 24.9% вблизи потомства культур и до 39.8% — в образцах, взятых вблизи потомства естественных популяций. Сумма ХЭ имеет одинаковую вариацию на уровне 12.5-12.6% (см. табл. 1 и 2). Мы проанализировали влияние этих показателей почвы на рост 90 семей, из которых 47 были из потомства естественных популяций и 43 — из потомства культур, которое оказалось следующим.
Корреляция органики в почве с высотой семей была недостоверна при г = 0.19±0.14 в потомстве Е и при г = 0.07±0.15 в потомстве К. Связь суммы 17 химических элементов почвы с высотой семей также оказалась недостоверна при г = 0.08±0.15 в потомстве Е и при г - - 0.19±0.15 в потомстве К. По-видимому, для доказательства их влияния на рост ели нужны значительно большие колебания этих двух характеристик почвы, чем наблюдалось в нашем опыте. Вероятно, это влияние проявится, если почва вообще будет разной. Для нас же важно было доказать наличие или отсутствие влияния некоторых ее различий под деревьями на содержание химических элементов в хвое.
Можно также полагать следующее. Во-первых, почва на этой бывшей вырубке после развития на ней высаженных культур ели спустя 20 лет стала более или менее однородной: во-вторых, почва относится к одному типу (эдатоп СзХ и колебания упомянутых ее характеристик (органики и суммы ХЭ) являются их естественной изменчивостью в пределах однородного биотопа, поэтому некоторые различия в составе почвы, имевшие место под от дельными деревьями, не повлияли на рост деревьев ели.
Таким образом, можно сделать вывод о том. что различия в концентрациях химических элементов и в содержании органики в достаточно однородной почве вблизи деревьев в опытных культурах, относящихся к одному биотопу7, имели естественную изменчивость и значимо на рост деревьев и на химический состав хвои у ели финской не повлияли.
Ранее [Рогозин. Разин, 2012] нами были обнаружены морфометрические отличия между родительскими естественными популяциями ели финской и ее культурами, которые характеризовали первые как более «сибирские», а вторые - как более «европейские» популяции этого естественного гибрида между елью сибирской и елью европейской. В данной работе выяснилось, что эти отличия имеют свое «химическое» продолжение в их потомствах, которые различаются по концентрациям химических элементов в хвое, а главное, имеют разную направленность трендов их накопления в хвое при разной устойчивости роста потомства. Они показывают, что в селекции ели имеется возможность эффективно использовать также и «химические» признаки (хемопризнаки), в частности. сочетание некоторых отдельных химических элементов и зольности хвои, изучение влияния которых уже выходит за пределы объемов данной статьи и будет рассмотрено нами в последующем.
Заключение
Программа выведения промышленного сорта ели финской основана на признании эпигенетического влияния материнских популяций, предусматривает обязательное научное сопровождение и массовые испытания потомства сотен семей. Только при таких условиях возможно обнаружение слабых связей, типичных для лесной селекции и необходимых для ее текущей корректировки. Отбор плюс-деревьев проводят в насаждениях -аналогах плантационных культур, т е. точно в таких же условиях, В каких будут выращивать ИХ ПОТОМСТВО. Применяют ранние оценки в 4—8 лет и ступенчатые испытания потомства. Из потомства от 500-1000 деревьев выделяют -70 деревьев-кандидатов в сорт. Изучение содержания химических элементов в хвое показало. что конкурентный или светолюбивый тип развития семьи определяют концентрации химических элементов в ней, и при отборе семей по некоторым параметрам их концентраций доля лучших семей повышается в 1.8—2 раза. Предложенная программа с учетом применения химического анализа хвои в 4-5 раз сокращает затраты и в 3 раза — сроки выведения сорта. Экспертная оценка стоимости программы на 10 лет оценивается в 40-45 млн руб. на один лесосеменной район.
Работа выполнена при финансовой поддержке задания 2014/153 государственных работ в сфере научной деятельности в рамках базовой части гос-задания Минобрнауки России, проект 144 № ГР 01201461915.
Список литературы Выведение промышленного сорта ели финской с использованием химических маркеров
- Аринушкина Е.В. Руководство по химическому анализу почв. М.: Изд-во МГУ, 1962. 491 с
- Видякин А.И. Эффективность плюсовой селекции древесных растений//Хвойные бореальной зоны. 2010. № 1-2. С. 18-24
- Коренев И.А. Продуктивность ели в связи с морфологической изменчивостью вида в подзоне южной тайги: дис.. к.с/х.н. Кострома, 2008. 125 с
- Крутовский К.В. Геномные и эпигеномные механизмы адаптации лесных древесных видов//Сохранение лесных генетических ресурсов Сибири: материалы 4-го междунар. совещания. Барнаул, 2015. С. 93-94
- Методы анализа минерального вещества, применяющиеся в центральной лаборатории ПО «Уралгеология». Свердловск, 1989. 81 с
- Милютин Л.И. и др. Сравнительный анализ структуры популяций ели европейской (Беларусь) и ели сибирской (Средняя Сибирь)//Современное состояние, проблемы и перспективы лесовосстановления и лесоразведения на генетико-селекционной основе: материалы междунар. науч. конф. Гомель, 2009. С. 108-112
- Нечаева Ю.С. и др. Анализ полиморфизма ДНК Picea obovata Ledeb. в Пермском крае//Биологические системы: устойчивость, принципы и механизмы функционирования: материалы IV Всерос. науч. -практ. конф. Н. Тагил, 2012. С. 93-96
- Одум Ю. Экология. М.: Мир, 1986. Т. 1. 328 с.; Т. 2. 376 с
- Плантационное лесоводство/под ред. И.В. Шутова. СПб.: Изд-во Политех. ун-та, 2007. 366 с
- Правдин Л.Ф. Ель европейская и ель сибирская в СССР. М.: Наука,1975. 199 с
- Попов П.П. Ель европейская и ель сибирская. Новосибирск: Наука, 2005. 230 с
- Реймерс Н. Ф. Экология (теории, законы, правила, принципы и гипотезы). М.: Журнал «Россия молодая», 1994. 367 с
- Рогозин М.В. Селекция сосны обыкновенной для плантационного выращивания : монография. Пермь, ПГНИУ, 2013. 200 с. (2,02 Мб). URL: http://elibrary.ru (дата обращения: 20.11.13)
- Рогозин М.В., Жекина Н.В., Комаров С.С., Кув шинская Л.В. Микроэлементы хвои в потомстве культур и естественных популяций ели финской//Вестник Пермского университета. Серия Биология. 2014. Вып. 3. С. 44-50
- Рогозин М.В., Разин Г.С. Лесные культуры Теплоуховых в имении Строгановых на Урале: история, законы развития, селекция ели. : Изд. второе. Пермь: ПГНИУ, 2012 -210 с. (6,75 Мб). URL: http://elibrary.ru (дата обращения: 20.11.13)
- Рогозин М.В., Разин Г.С. Развитие древостоев. Модели, законы, гипотезы : монография/под ред. М.В. Рогозина. Пермь, 2015. 277 с. (11 Мб). URL: http://elibrary.ru/item.asp?id=24420793 (дата обращения: 10.11.15)
- Рябоконь А. П. Динамика сортиментной структуры сосновой древесины при различных режимах выращивания//Лесное хозяйство. 1990. № 2. С. 48-50
- Туркин А.А. Испытание потомства плюсовых деревьев сосны обыкновенной на примере Республики Коми: дис.. к.с/х.н. Сыктывкар, 2007. 144 с
