Взаимодействие эндогенных и экзогенных факторов в биологических ритмах южного одноперого терпуга Pleurogrammus azonus в водах Приморья

Автор: Вдовин А. Н., Четырбоцкий А. Н., Измятинский Д. В.
Журнал: Вестник Мурманского государственного технического университета @vestnik-mstu
Рубрика: Биологические науки
Статья в выпуске: 2 т.26, 2023 года.
Бесплатный доступ
Южный одноперый терпуг Pleurogrammus azonus является важным промысловым видом в водах России и Японии. В российских водах Японского моря регулярное изучение его биологии проводилось на акватории Приморского края, где выполнялись ежегодные траловые съемки. Сборы материала охватывали весь календарный год. В ходе исследования установлено, что в согласовании биоритмов южного одноперого терпуга решающую роль играют эндогенные ритмы (внутренние биологические часы). Это предположение подтверждается разной продолжительностью экзогенных и эндогенных ритмов: динамика температурного показателя имеет выраженную периодизацию, равную одному астрономическому году, а продолжительность цирканных ритмов в динамике физиологических процессов и главных компонент меняется с возрастом. Связь показателей соматического и генеративного роста с осцилляцией температуры подвержена возрастной изменчивости. У молоди эта связь явно не проcлеживается. В начале периода половозрелости терпуга проявляются значимые связи. Затем зависимость показателей роста от осцилляции температуры снижается, поскольку возрастает избирательная способность рыб к выбору температурного диапазона. Избирательная способность к условиям среды в большей степени определяется не индивидуальным, а групповым поведением вида.
Южный одноперый терпуг Pleurogrammus azonus, биоритмы, экзогенные и эндогенные факторы, соматический и генеративный рост, осцилляция температуры, Arabesque greenling Pleurogrammus azonus, biorhythms, exogenous and endogenous factors, somatic and generative growth, temperature oscillation
Короткий адрес: https://sciup.org/142237543
IDR: 142237543 | УДК: 597.556.334.1–13(265.54) | DOI: 10.21443/1560-9278-2023-26-2-99-111
Текст статьи Взаимодействие эндогенных и экзогенных факторов в биологических ритмах южного одноперого терпуга Pleurogrammus azonus в водах Приморья
Южный одноперый терпуг Pleurogrammus azonus – важный промысловый вид в водах России и Японии. В российских водах Японского моря регулярное изучение его биологии проводится на акватории Приморского края, где выполняются ежегодные траловые съемки.
Адаптация вида к изменчивости среды происходит через упорядоченное распределение физиологических процессов по времени ( Дольник, 1975; Шилов, 2001 и др.). Внутренние изменения нередко опережают, "предсказывают" внешние, что позволяет особи (группе особей) оптимально приспособиться к изменившимся внешним условиям.
Согласованность биологических ритмов внутри организма определяется взаимодействием эндогенных (физиологических) и экзогенных (экологических) ритмов ( Мина и др., 1976; 1980; Браун, 1977 и др.). В ряду экзогенных особо выделяются такие мощные факторы, как кормовая база и температура ( Бретт, 1983; Elliot, 1975 ).
Стадийность онтогенеза и ритмичность физиологических процессов изучена нами на примере южного одноперого терпуга Pleurogrammus azonus Jordan et Metz, 1913 (сем. Hexagrammidae) и представлена в работах ( Вдовин и др., 2018; 2019 ), где взаимодействие организма со средой детально не рассматривалось (приведены только отдельные факты). Настоящее исследование является продолжением указанных публикаций. Цель работы состоит в оценке влияния возрастной изменчивости биологического состояния терпуга на согласованность биологических ритмов с внешними экзогенными ритмами, которая определяется взаимодействием физиологических циклов с условиями среды.
Материалы и методы
Биологический материал собран в 1960–1996 гг. в морских водах Приморья (подзона Приморье, южнее м. Золотой); использованы пробы, взятые на научно-исследовательских и промысловых судах, а также рыбокомбинатах Приморья. Для биологического анализа было собрано 7 155 экземпляров. Содержание депозитного жира определено у 2 407 особей. У 1 112 экземпляров взвешивались гонады и пищевой комок. Пробы на содержание белка взяты у 906 рыб. Методы сбора проб и первичной обработки материала по физиологическим показателям (длина и масса тела, масса гонад, депозитного жира, белка и пищевого комка) представлены в наших предыдущих работах ( Вдовин и др., 1993; Vdovin et al., 2014; Вдовин и др., 2015а; б; 2018; 2019 ).
Показатели физиологических процессов имеют следующие обозначения: FL , см – стандартная длина; Ws , г – соматические значения массы тела; Q , г – масса гонад; Fa, г – общая масса депозитного жира, являющаяся суммой печеночного, полостного и мышечного жиров; Pr , г – масса белка; f o, г – масса пищевого комка. Удельные скорости физиологических процессов Ƴ рассчитаны по формуле (1а), указанной в работе А. Н. Вдовина и А. Н. Четырбоцкого ( 2018 ):
i x ( ti + a - xy ( t i )
-
( i ) X ( t) t i + 1 - t- ’
где X ( t ) – значения параметров физиологических процессов в момент времени t .
Измерения температуры воды проводились в ходе научно-исследовательских рейсов на гидрологических станциях в 1978–2018 гг.; они были выполнены совместно с тралениями (или с постановкой дрифтерных сетей) при наличии в уловах южного одноперого терпуга. Общее количество таких станций составило 6 679: 6 114 – на донных тралениях; 449 – на пелагических тралениях; 86 – на станциях дрифтерного лова. Количество измерений температуры было меньше, чем количество станций, так как: 1) не на всех станциях измерялась температура; 2) некоторые значения являлись артефактами. При исключении артефактов (нереально низких или высоких значений температур) ориентировались на опубликованные термические характеристики водных масс российских вод Японского моря ( Зуенко, 1994; 1998; 2008 ). Выборка промеров температуры у дна составила 3 170 измерений, у поверхности – 449 измерений; всего осуществлено 3 624 измерения. Показателем температурного фона служил размах вариации VR – разница между крайними значениями температуры. Другие параметры, представляющие какой-либо центр распределения значений температуры (средняя, мода, медиана), не показали значимых связей с интенсивностью физиологических процессов.
Для количественной оценки интенсивности физиологических процессов использовалась выборка наблюдений, которая характеризует 12 признаков: по 6 вышеприведенных физиологических показателей для самцов и самок. Ее построение выполнялось с помощью процедуры princomp системы MATLAB (Дьяконов и др., 2002). Целесообразность использования процедуры обусловлена потребностью построения небольшого числа интегральных показателей, которые обеспечивают хорошую аппроксимацию исходных коррелированных наблюдений. При работе с большим выборочным многопараметрическим числовым материалом наблюдается взаимозависимость между динамическими рядами отдельных показателей, где для представления парной связи между ними обычно используется коэффициент корреляции (диапазон его изменения от –1 до 1). В случае его высокого значения (не ниже 0,5) целесообразно на базе таких показателей сформировать новый набор интегральных показателей, куда каждый из исходных показателей вносит определенный вклад. Этот вклад оценивается на основании матрицы корреляции между показателями. Количество данных интегральных показателей равно количеству исходных признаков. Вместе с тем наиболее важная информация определяется набором только первых из них (на практике обычно первых двух показателей). Процедура оценки этих вкладов в формирование интегрального показателя выполняется методом главных компонент (ГК). В системе Matlab реализация алгоритма выполняется с использованием процедуры princomp (principal component), входным параметром которой является матрица выборочного распределения, а на выходе – значения главных компонент и весовые нагрузки для исходных переменных.
Здесь первая главная компонента ГК1 обеспечивает 46,6 % исходной дисперсии многомерной выборки (ее доля среди собственных значений матрицы корреляции выборочных данных), а вторая компонента ГК2 – 41 %. Эта ситуация отражает тот факт, что первые две главные компоненты показывают с высокой степенью достоверности характер связей выборочных данных.
При хронологическом описании физиологических процессов нами использовался термин "цирканный (цирканнуальный, окологодичный) ритм", поскольку биологические ритмы, как правило, не равны геофизическим ( Браун, 1977; Шилов, 2001 ).
В отличие большинства схем эндогенных часов концепция хронона основывается на представлениях молекулярной биологии ( Браун, 1977 ). ДНК программирует синтез РНК, а РНК – синтез белка. Для транскрипции всего комплекса необходимо около 24 часов, после чего начинается новый цикл. Скорость генетической транскрипции регулируется целым комплексом клеточных факторов. Настройка эндогенных часов частично осуществляется экзогенными часами (суточная ритмика освещенности) и внешними сезонными факторами (например, температура). Внешние факторы действуют на молекулярные опосредованно, через клеточные факторы. На клеточные факторы также влияют физиологические процессы, интенсивность которых отчасти зависит от внешних условий. Биоритм организма складывается под воздействием комплекса причин и может меняться с возрастом. На наш взгляд, это единственная известная нам схема эндогенных часов, которая позволяет наглядно продемонстрировать взаимодействие эндогенных и экзогенных факторов.
Результаты и обсуждение
Стадийность онтогенеза южного одноперого терпуга определяется интенсивностью физиологических процессов и сменой их приоритетности с возрастом. Первые два года преобладает накопление вещества, а далее – циклическое чередование ассимиляции и диссимиляции. С увеличением возраста снижается интенсивность физиологических процессов и в целом обмена веществ ( Вдовин и др., 2018) .
С возрастом также четче проявляется цикличность изменения значений физиологических процессов. Это проявляется и в динамике таких интегральных показателей, как главные компоненты ГК1 и ГК2 (рис. 1). Более четко периодичность проявляется в динамике значений ГК2. Хронологические отрезки между минимальными значениями, соответствующие окологодичному (цирканному) биоритму, составляют 11–12 месяцев. Изменчивость размеров подобных хронологических отрезков для ГК1 составляет 10–14 месяцев. В целом окологодичный жизненный цикл терпуга длится от 9–11 до 13 месяцев, увеличиваясь с возрастом ( Вдовин и др., 1993 ).
Периодичность динамики ГК2 подобна периодичности активности питания, которая выражается через массу пищевого комка. Коэффициент корреляции между этой компонентой и удельной скоростью динамики массы пищевого комка составляет 0,895. Следующим по значимости показателем для ГК2 является динамика депозитного жира. Корреляция между удельной скоростью данного показателя и ГК2 равна 0,548. Питание является физиологическим процессом, т. е. эндогенным фактором, и в первую очередь определяется потребностью организма. Последнее напрямую подтверждается тем, что с возрастом уменьшается относительное потребление пищи ( Вдовин и др., 2018 ). Такой внешний фактор, как кормовая база, в российских водах Японского моря рыбами используется только частично. При этом терпуг не имеет явных конкурентов ( Пущина, 2005; Пущина и др., 2014 ). Таким образом, кормовая база не может оказывать на терпуга не только ультимативного, но и сколько-нибудь значительного влияния. Корреляция с показателем температуры для ГК2 составляет всего 0,225.
Периодичность и ход динамики ГК1 практически идентичны таковым для половых продуктов. Корреляция между ГК1 и удельной скоростью динамики массы гонад составляет 0,88. Связь между ГК1 и VR , ° С, невелика – коэффициент корреляции равняется 0,429. Значимость коррелятивной связи на третьем-пятом годах (от 2+ до 4+) заметно выше 0,522. Таким образом, некое упорядочение хода динамики физиологических процессов (периодизация и согласованность) происходит после снижения темпов соматического роста на третьем году жизни ( Вдовин и др., 2018 ).
Сходство хода циклических (цирканных) ритмов становится очевидным для ГК с четвертого года (рис. 1). Возрастная изменчивость цирканных биоритмов у терпуга отмечалась и ранее (Вдовин и др., 1993; 2015а). Динамика же температурного показателя имеет четкую периодизацию, равную астрономическому году. С возрастом меняется только размах колебаний температурного показателя (рис. 1). Взаимосвязь интенсивности физиологических процессов с температурным фактором также имеет выраженную возрастную изменчивость (таблица; рис. 2). При рассмотрении всего возрастного ряда зависимость физиологических показателей и главных компонент от температурного фактора не прослеживается: все значения коэффициента корреляции ниже 0,5 (таблица). В возрастном ряду от 2+ до 6+ (впервые созревающие и половозрелые особи) корреляция выше 0,5 отмечается для трех физиологических процессов: линейного (FL), белкового (Pr) и соматического (Ws) роста. У отдельных возрастных групп отмечаются значения коэффициентов корреляции Y(fo) и ГК1 немногим выше 0,5.
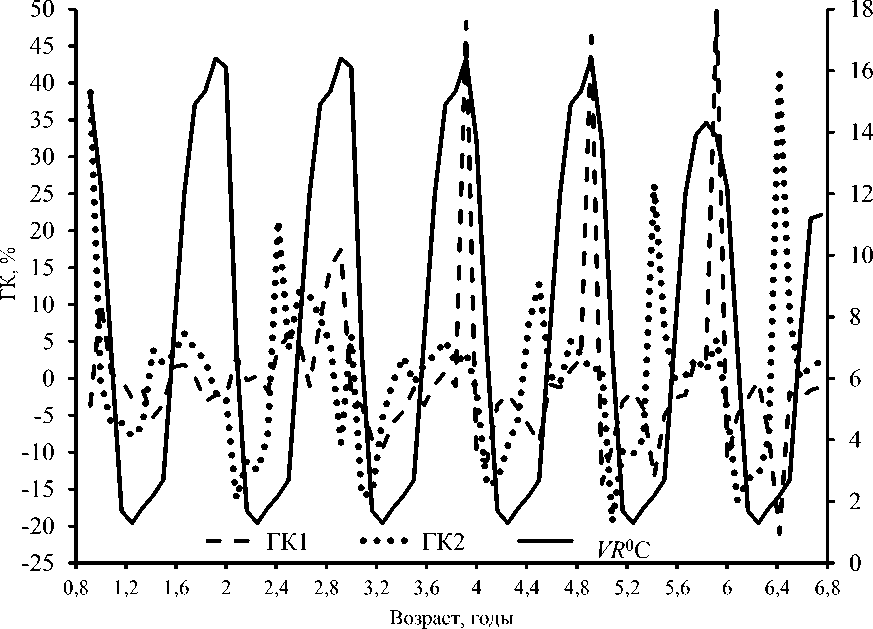
Рис. 1. Возрастное распределение значений главных компонент ГК1 и ГК2 и размаха вариации температуры VR в водах Приморья у Pleurogrammus azonus Fig. 1. Age distribution of the values of the principal components (PC1 – first, PC2 – second) and the range of temperature variation (VR) in the habitat (in the waters of the Primorsky Krai) of the Arabesque greenling Pleurogrammus azonus
Таблица. Возрастная изменчивость коррелятивных связей размаха вариации температуры VR у Pleurogrammus azonus с удельной скоростью физиологических процессов и главными компонентами в водах Приморья Table. Age-related variability of correlative relationships of the scope of variation of the temperature (VR) in the habitat (in the waters of the Primorsky Krai) of the Arabesque greenling Pleurogrammus azonus with the specific rate of physiological processes and the principal components
|
Возраст T, годы |
Y ( FL ) |
Y ( Pr ) |
Y ( Ws ) |
Y ( Q ) |
Y ( Fa ) |
Yf o) |
ГК1 |
ГК2 |
|
0+ |
– 0,583 |
– 0,453 |
– 0,456 |
– |
– |
–0,423 |
– |
– |
|
1+ |
0,283 |
0,151 |
0,278 |
0,379 |
– 0,040 |
0,407 |
0,182 |
0,338 |
|
2+ |
0,816 |
0,835 |
0,743 |
0,528 |
– 0,237 |
0,117 |
0,513 |
0,214 |
|
3+ |
0,714 |
0,626 |
0,567 |
0,548 |
– 0,120 |
–0,015 |
0,542 |
0,466 |
|
4+ |
0,815 |
0,637 |
0,631 |
0,473 |
– 0,174 |
–0,050 |
0,509 |
0,290 |
|
5+ |
0,611 |
0,684 |
0,554 |
0,436 |
– 0,079 |
–0,199 |
0,463 |
0,072 |
|
6+ |
0,501 |
0,640 |
0,578 |
0,399 |
0,091 |
–0,163 |
0,363 |
–0,036 |
|
7+ |
0,529 |
0,479 |
0,286 |
0,428 |
– 0,104 |
–0,069 |
– |
– |
|
От 0+ до 7+ |
0,479 |
0,421 |
0,420 |
0,376 |
– 0,100 |
–0,036 |
0,431 |
0,233 |
|
От 2+ до 6+ |
0,700 |
0,595 |
0,616 |
0,480 |
– 0,112 |
–0,058 |
0,483 |
0,212 |
Примечание. Положительные значения корреляции величиной больше 0,5 выделены жирным шрифтом.
Самые слабые связи с температурой для всего возрастного ряда (близкие к нулю) отмечены для интенсивности питания и жиронакопления. Отметим, что два этих процесса тесно взаимосвязаны ( Вдовин и др., 2018 ). Интенсивность питания и темпы жиронакопления характеризуются низкими коррелятивными связями со скоростями других физиологических процессов. Высокая корреляция удельной скорости массы пищевого комка ( R = 0,895) отмечается только с ГК2. Можно предположить, что интенсивность питания не определяется каким-либо физиологическим процессом, связанным с такими факторами среды, как температура и кормовая база, а зависит от общих потребностей организма. О малозначащей роли кормовой базы уже сказано выше. О влиянии температуры на скорость пищеварения и интенсивность питания издано множество публикаций. Мы полагаем уместным сослаться на работу ( Пономарев, 1993 ): благодаря температурной компенсации пищеварительной функции, рыбы обладают способностью к эффективному гидролизу пищи в любое время года. Разумеется, речь не идет о значениях температур, выходящих за пределы толерантности. Однако терпуг и не выходит за пределы толерантности, а обычно обитает в пределах температурного оптимума, активно перемещаясь в градиентном поле температуры ( Вдовин, 1998 ). По данным Симомура и Фукутаки, полученным в 1957 г. [цитируется по работе ( Горбунова, 1962 )], уже личинки терпуга обладают высокой избирательной способностью к условиям среды – они активно избегают районов с температурой выше 7 °C и соленостью ниже 17 ‰.
Жир и пища, как внутренний и внешний источники энергии, вероятно, необходимы для воплощения эргономической стратегии посредством адаптивной самоорганизации и не могут следовать какому-либо одному процессу. Приоритетной составляющей в эргономической стратегии является соотношение скорости и эффективности обмена веществ ( Бретт и др., 1983; Шмидт-Ниельсен, 1987 ). Известно, что при снижении скорости обмена веществ повышается его эффективность. Данная ситуация ранее описывалась для терпуга ( Вдовин и др., 2018 ). Определяющими для биологического состояния организма являются процессы соматического и генеративного роста. Интенсивность именно этих процессов определяет стадийность онтогенеза. Тем не менее динамика соматического и генеративного роста не играет определяющей роли в регуляции интенсивности физиологических процессов, а, скорее всего, задается генетической программой. Структура различий биологического состояния на разных этапах и периодах онтогенеза может определяться фенотипической настройкой ( Вдовин и др., 2018 ).
Действительно, одним из критериев возрастной периодизации является оценка взаимодействия организма с условиями среды ( Антонова, 2006 ). Температура – один из определяющих внешних факторов, влияющих на обмен веществ ( Дгебуадзе, 2001 и др.). Широко распространено мнение, что у пойкилотермных животных повышение температуры ускоряет биохимические реакции и повышает интенсивность физиологических процессов ( Шмидт-Ниельсен, 1982 и др . ). Однако многочисленные эксперименты показывают, что большее стимулирующее действие на обмен веществ рыб оказывают смена температурного режима и особенно осцилляция температуры ( Бретт, 1983; Константинов и др., 1989, 1996 ). В работе ( Проссер, 1977 ) приводится интересный пример в опытах с серебряным карасем Carassius auratus gibelio : избыточный подъем интенсивности обмена наблюдается при быстром изменении температуры в том и другом направлении.
Важнейшая роль температуры, по мнению некоторых исследователей, видится в синхронизации биоритмов ( Браун, 1977 ). При этом синхронизация более эффективно проявляется не у отдельного индивидуума, а у группы особей ( Слоним, 1971 ). В частности, в экспериментах с грызунами при понижении температуры обмен веществ в группе особей понижался сильнее, чем у отдельных индивидуумов. Согласованность биоритмов в группе особей определяется взаимным влиянием друг на друга. Примеры, имеющиеся для южного одноперого терпуга, весьма показательны. У особей одного возраста, но разного биологического состояния, физиологические и миграционные процессы проходят сходным образом ( Вдовин, 1998 ). После перехода к придонно-пелагическому образу жизни распределение молоди и половозрелых рыб принципиально не различается, о чем свидетельствует тот факт, что в нерестовый период неполовозрелые рыбы и производители, особенно одного возраста, совершают сходные миграции ( Вдовин и др., 1994 ).
Динамика массы половых продуктов у особей, участвующих и не участвующих в нересте, также подобны ( Вдовин и др., 1995 ). Сигнальным фактором для начала нереста служит понижение температуры на нерестилищах. Абсолютная величина температуры особой роли не играет, поскольку ее значения на разных нерестилищах отличаются на 20–40 % ( Горбунова, 1962; Ковтун, 1979; Гомелюк, 1987; Китакама и др., 1967 ). Нерест начинают самые крупные особи. По мере уменьшения размеров и возраста сдвигаются сроки нереста (становятся более поздними), что обусловлено территориальным поведением терпуга. Крупные производители занимают глубокие и обширные нерестовые участки ( Гомелюк, 1987 ). В период весенней нагульной миграции половозрелые особи подходят к берегу раньше молоди ( Вдовин, 1998 ).
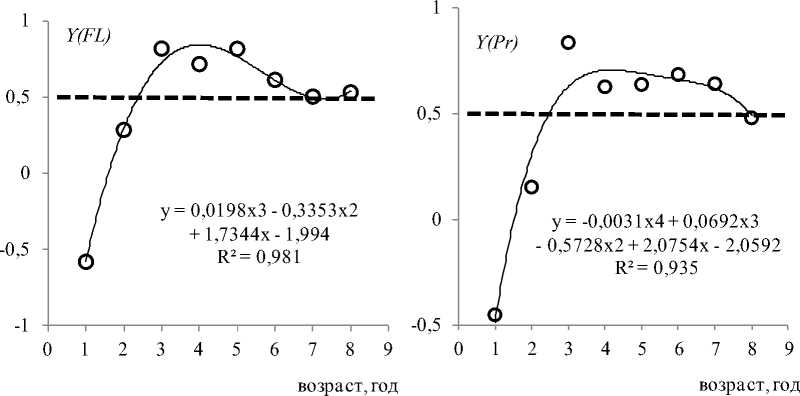
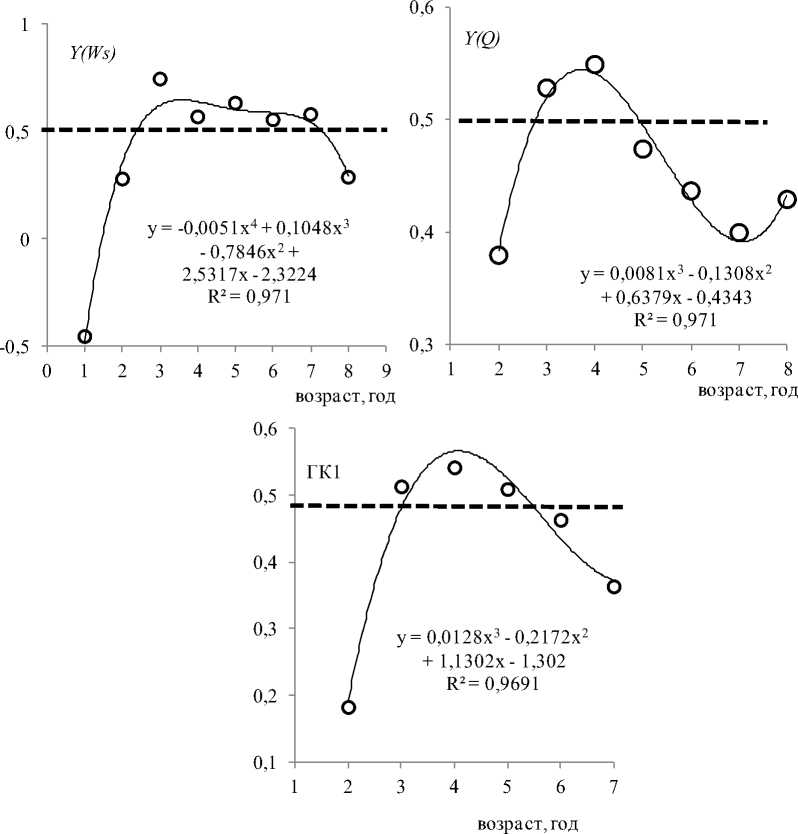
Рис. 2. Возрастная изменчивость коррелятивных связей размаха вариации температуры VR с удельной скоростью физиологических процессов и ГК1 у Pleurogrammus azonus в водах Приморья. По оси ординат – значения коэффициентов корреляции. Пунктирная линия разделяет значения R по 0,5 Fig. 2. Age-related variability of correlative relationships of the scope of variation of the temperature ( VR ) in the habitat (in the waters of the Primorsky Krai) of the Arabesque greenling Pleurogrammus azonus with the specific rate of physiological processes and PC1. There are the values of the correlation coefficients on the ordinate axis. The dotted line divides the R values by 0.5
Отметим, что именно у половозрелых особей связь интенсивности отдельных физиологических процессов и ГК1 с температурным фактором проявляется более отчетливо, чем у неполовозрелых (рис. 2). Ход возрастной динамики связи температурного фактора с перечисленными показателями схож. В течение двух первых лет (молодь) корреляция ниже 0,5, при этом она заметно возрастает на втором году жизни. С третьего года жизни корреляция температуры с рассматриваемыми показателями в течение 2–6 лет выше 0,5. Самые высокие значения корреляционных связей осцилляции температуры отмечаются с Ƴ ( FL ) и Ƴ ( Pr ) (рис. 2) . Отметим, что линейный рост является весьма условным показателем и отражает и другие процессы, в частности процесс белкового роста ( Вдовин и др., 1993 ). Белковый, как и линейный, рост у терпуга носит поступательный характер – без потерь белковой массы с возрастом. Удельная скорость белкового роста тесно связана с удельными скоростями линейного и весового роста. Корреляция между этими показателями больше 0,9. Коэффициенты корреляции между остальными показателями не превышают 0,5. Основным показателем роста следует считать удельную скорость белкового роста. Содержание белка и размеры костей являются абсолютными показателями роста ( Бретт и др., 1983; Антонова, 2006 ).
Тем не менее не следует забывать, что рост является комплексным процессом и тесно связан с развитием ( Мина и др., 1976; Зотин и др., 1993 ). Поэтому мы полагаем, что эти три показателя (соматическая масса тела, содержание белка и линейные размеры тела) характеризуют один процесс – соматический рост.
В течение первых двух лет определенной связи соматического роста с температурным фактором не наблюдается (рис. 2). Этот хронологический отрезок ранее назван нами ювенальным этапом ( Вдовин и др., 2018 ). На данном этапе преобладает активное накопление органического вещества с малозаметными кратковременными потерями. В первый год жизни (ювенильный период ювенального этапа) при самых высоких темпах соматического роста у всех трех его показателей отмечаются обратные связи с осцилляцией температуры (таблица; рис. 2). Происходит это потому, что при тренде снижения темпов соматического роста температурный тренд имеет противоположную направленность. На первом году жизни темпы роста у рыб всегда самые высокие ( Campana, 2001 ). Точнее, они должны быть самыми высокими всегда, за исключением ультимативных условий среды.
На втором году жизни (пубертатный период ювенального этапа) при некотором снижении темпов соматического роста и активизации генеративного роста и жиронакопления корреляция показателей соматического роста с жиронакоплением становится положительной (таблица; рис. 2), но ее значения меньше 0,5. Постепенная перестройка организма к этапу половозрелости не нуждается в стимуляции обмена веществ температурой. Роль температурного фактора, возможно, начинает проявляться в отдельные моменты, в частности с апреля по июнь, когда при весеннем подъеме температуры активизируется генеративный рост.
На следующем этапе онтогенеза – половозрелости, длящегося с третьего по восьмой год жизни ( Вдовин и др., 2018 ), связь показателей соматического роста с размахом вариации температуры весьма стабильна (рис. 2). Полиномиальные тренды показывают наличие устойчивой тенденции: связь показателей роста с температурным фактором постепенно снижается к концу жизни. Указанная тенденция прослеживается и для показателя генеративного роста Ƴ (Q), и для ГК1 (рис. 2). В данном случае уместнее говорить не о ходе отдельных процессов, а о зависимости от температуры интенсивности обмена веществ, которую эти процессы отражают. Полагаем, что данная зависимость определяется степенью согласованности физиологических процессов. Особенно обращает на себя внимание подобие хода генеративного роста у половозрелых и впервые созревающих рыб. Несмотря на качественные различия гаметогенеза в каждой возрастной группе, динамика хода генеративного роста имеет сходный характер ( Вдовин и др., 1995 ). Механизм приведения в соответствие процессов генеративного роста с другими физиологическими процессами позволяет одновозрастным рыбам относительно одновременно участвовать в нересте ( Вдовин, 1998 ). Корреляция между процессами соматического и генеративного роста с температурным фактором позволяет предположить его значительную регулирующую роль в синхронизации этих процессов.
Как было отмечено выше, влияние температуры на рыб проявляется эффективней для группы видов, чем для отдельного индивидуума ( Вдовин, 1998; Вдовин и др., 1994;1995; Слоним, 1971 ). В данном случае имеются особи, которые первыми реагируют на изменения температурного фактора, а их поведение стимулирует к подобным действиям других рыб.
По мере увеличения возраста корреляция размаха вариации температуры с показателями соматического и генеративного роста уменьшается. Последнее связано с возрастанием самоорганизации для реализации эргономической стратегии (Вдовин и др., 2018). Во-первых, с возрастом увеличивается конкурентоспособность в борьбе за благоприятные условия. Крупные самцы первыми занимают лучшие нерестовые участки и не пускают туда молодых (Гомелюк, 1987). Избирательная способность к условиям среды также увеличивается, в частности снижается температурный диапазон (рис. 1). Снижение температурного диапазона у терпуга с возрастом происходит направленно. Иначе говоря, потребность в стимуляции обменных процессов температурой снижается. Рыбы крупнее 36 см перестают встречаться в пелагиали. Такого размера терпуг в массе достигает к концу пятого года жизни. Во-вторых, снижение температурного диапазона должно способствовать снижению интенсивности общего обмена, что и происходит у терпуга с возрастом. Таким образом, воплощается адаптивная самоорганизация эргономической стратегии (Вдовин и др., 2018). О том, что интенсивность обмена и питания тесно связана с температурой, говорится во многих источниках (Кафанов и др., 1983; Фандж и др., 1983 и др.). По нашим данным, интенсивность питания не коррелирует с температурой (таблица). По мнению В. И. Пономарева (1993), активность пищеварения и не должна быть тесно связана с температурой среды из-за температурной компенсации пищеварительной функции. Однако подобная связь в условиях экспериментов, длящихся непродолжительное время, может проявляться. В нашем случае максимальная интенсивность питания непродолжительное время (июль -сентябрь) также наблюдается на фоне самой высокой осцилляции температуры. Полагаем, что интенсивность питания и жиронакопления (физиологических процессов, слабо коррелирующих с температурой) не зависит от внешних условий, а определяется потребностями организма. По данным, указанным в работе (Суханов, 1979), пойкилотермный организм в течение всего года постоянно находится в условиях среды, близких к оптимальным для роста. Снижение с возрастом температурного диапазона и обмена веществ также является благоприятным фактором для терпуга.
Снижение обмена веществ с возрастом - явление универсальное. Важнее тот факт, что по мере взросления уменьшение интенсивности обмена сопровождается повышением его эффективности ( Бретт и др., 1983; Шмидт-Ниельсен, 1987) . Снижение обмена веществ способствует увеличению продолжительности жизни ( Радзинская и др., 1987; Зотин и др., 1993) . В статьях ( Суханов, 1979; Кафанови др., 1983 ) на основе опубликованной информации и собственных расчетов выводится постулат о зависимости продолжительности жизни от температурного режима, что связано с замедлением обмена веществ. По мнению В. В. Суханова ( 1979 ), "по мере роста и увеличения возраста организмы с оптимальным поведением должны мигрировать в среду с более низкими температурами". Данный постулат убедительно подтверждается нашими данными по терпугу. Разумеется, продолжительность жизни зависит не только от обмена веществ. Предельная продолжительность жизни у самок на год больше, чем у самцов: вероятно, процессы обновления организма у самок эффективнее, чем у самцов ( Зотин и др., 1993 ). Тем не менее стадийность онтогенеза у обоих полов почти идентична, а сезонная динамика интенсивности физиологических процессов почти не отличается ( Вдовин и др., 2018 ).
Биологическая ритмика прежде всего определяется наследственной программой ( Мина и др., 1976; Браун, 1977 и др.). Точкой отсчета окологодового (цирканного) ритма следует считать нерест, сроки которого отличаются у разных популяций. Темпы роста и полового созревания у южного одноперого терпуга, обитающего в водах России и Японии, весьма схожи ( Вдовин, 1998; Вдовин и др., 2018; Irie, 1986; Takashima et al., 2016) . Из существующих схем биологических (эндогенных) часов, основывающихся на молекулярных представлениях, наиболее убедительной и простой, на наш взгляд, является концепция хронона ( Браун, 1977) : "Предполагают, что существуют очень длинные фрагменты ДНК, на протяжении которых скорость генетической транскрипции регулируется целым комплексом клеточных факторов...". На клеточные факторы, в свою очередь, могут оказывать влияние внешние факторы.
Для согласования физиологических процессов, а в целом и для регуляции обмена веществ, температура окружающей среды имеет большое значение. Однако терпуг способен контролировать этот фактор благодаря активному перемещению в градиентном поле. Самый широкий температурный диапазон наблюдается в июле - августе, в преднерестовый период, когда молодь и половозрелые особи совершают миграции, направленные к берегу (и из придонных слоев воды в пелагиаль) ( Вдовин, 2004 ) (рис. 1). Резкая смена температурного режима позволяет активизировать обмен веществ, а также соматический и генеративный рост ( Вдовин и др., 2018 ).
О взаимосвязи разных уровней экзогенных и эндогенных ритмов имеется множество публикаций. Схему согласования биоритмов можно описать следующим образом:
-
1. На молекулярном уровне (хронон) скорость генетической транскрипции (соответственно, синтез пептидных цепей) регулируется комплексом клеточных факторов (ферментами и гормонами, а также комплексом метаболитов) ( Браун, 1977) . Гормональная и ферментативная активность во многом зависит от условий среды и главным образом от температурного режима ( Проссер, 1977) .
-
2. На уровне органов и тканей интенсивность и приоритетность физиологических процессов меняется в зависимости от биологического состояния организма. В течение первых двух лет преобладает накопление вещества, в дальнейшем - чередование фаз ассимиляции и диссимиляции ( Вдовин и др., 2018 ). Широкий температурный диапазон активизирует обмен веществ, а узкий - снижает его интенсивность. Данная зависимость имеет не только сезонную, но и возрастную динамику. В теплый период года расширяется температурный диапазон обитания и активизируются обменные процессы. С возрастом снижается обмен веществ и сужается температурный диапазон (рис. 1).
-
3. На уровне организма температурный режим также играет роль сигнального. В частности, он служит сигналом начала нереста для самых зрелых особей. Наличие индивидуальной биологической ритмики и индивидуальных вариаций биологического состояния позволяет предположить разную ответную реакцию особей одного возраста на внешние раздражители ( Браун, 1977 ).
-
4. Особи одного возраста, но разного биологического состояния, характеризуются подобием хода физиологических процессов и совершают сходные миграции. Так, у созревающих особей и половозрелых рыб отмечается подобие хода динамики генеративного роста ( Вдовин и др., 1995 ). Особи способны оказывать влияние друг на друга, вследствие чего повышается групповая избирательность к условиям среды. Первыми в район нереста следуют самцы, затем самки. Эту же миграцию без захода на нерестилища повторяют созревающие, но не готовые к нересту особи ( Вдовин и др., 1990; 1994 ). Таким образом, синхронизация биоритмов более эффективно проявляется не у отдельного индивидуума, а у группы особей ( Слоним, 1971 ).
-
5. Для демонстрации согласованности нами предлагается упрощенная схема, в основу которой положена концепция хронона ( Браун, 1977 ) (рис. 3). Предполагается, что настройка биологических часов (хронон) определяется клеточными факторами. На клеточные факторы, в свою очередь, могут влиять физиологические процессы, а точнее – их интенсивность. Физиологические процессы и клеточные факторы взаимосвязаны. На клеточные факторы, физиологические процессы и биоритмы организма оказывают влияние внешние условия. Непосредственное влияние внешних факторов на биоритмы может проявляться как сигнал к смене биологического сезона. Возрастной биоритм у терпуга определяется способностью выбора оптимальных условий среды.
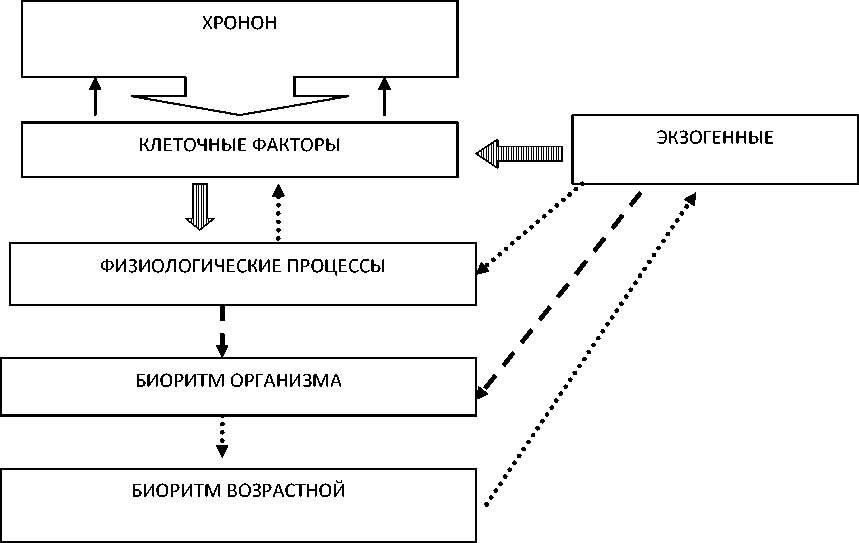
Рис. 3. Схема согласования биоритмов южного одноперого терпуга Pleurogrammus azonus Fig. 3. Scheme of coordination of the Arabesque greenling Pleurogrammus azonus biorhythms
Заключение
В синхронизации биоритмов южного одноперого терпуга решающую роль играют эндогенные ритмы (внутренние биологические часы). Данное предположение подтверждается аргументами, свидетельствующими о разной продолжительности экзогенных и эндогенных ритмов:
-
– динамика температурного показателя в среде обитания терпуга имеет выраженную периодизацию, равную одному астрономическому году. Продолжительность цирканных ритмов в динамике физиологических процессов и главных компонент меняется с возрастом. Изменчивость цирканного ритма физиологических процессов терпуга по ГК1 составляет 10–14 месяцев, по ГК2 – 11–12 месяцев. В целом окологодичный жизненный цикл терпуга длится от 9–11 до 13 месяцев, увеличиваясь с возрастом;
-
– возрастной изменчивости подвержена связь показателей соматического и генеративного роста с температурным фактором. Тенденция этой изменчивости проявляется одинаково для всех показателей роста. В течение первых двух лет, когда в росте терпуга не проявляется явная периодичность и преобладает процесс накопления органического вещества, корреляция с температурным фактором незначительна. В начале периода половозрелости наблюдаются значимые связи, затем зависимость показателей роста от температуры снижается, поскольку возрастает избирательная способность терпуга к выбору температурного диапазона;
-
– избирательная способность к условиям среды в большей степени определяется не индивидуальным, а групповым поведением терпуга.
Конфликт интересов
