Взаимоотношения леса и степи на Приволжской возвышенности (в порядке дискуссии)
")
Автор: Малышева Г.С.
Журнал: Фиторазнообразие Восточной Европы @phytodiveuro
Статья в выпуске: 2 т.12, 2018 года.
Бесплатный доступ
С использованием палеогеографического метода выявлены закономерности в распространении лесного и степного типов растительности в средней части Приволжской возвышенности в зависимости от ее морфоструктурных особенностей. Полученные результаты подтверждают представления об устойчивом равновесии и автохтонном развитии этих типов растительности в лесостепной зоне.
Лесостепь национальный парк "хвалынский" саратовская область приволжская возвышенность
Короткий адрес: https://sciup.org/148314622
IDR: 148314622 | УДК: 581.524.42 | DOI: 10.24411/2072-8816-2018-10019
Текст научной статьи Взаимоотношения леса и степи на Приволжской возвышенности (в порядке дискуссии)
Ботанико-географический статус лесостепи вызывает много вопросов. Большинство геоботаников считают ее зоной, некоторые рассматривают ее в качестве северной подзоны степной зоны, а другие называют переходной полосой между зоной широколиственных лесов и степной. Обзор данной проблемы рассмотрен В.А. Демьяновым (1996).
Важным моментом для уточнения понятия «лесостепь» является изучение пространственной структуры ее растительного покрова. В качестве объекта для решения этого вопроса выбрана территория национального парка «Хвалынский» (далее – парк), расположенного в средней части Приволжской возвышенности на границе двух ботаникогеографических провинций: Восточноевропейской лесостепной и Причерноморской [Понтической] степной (Исаченко, Лаврен-ко, 1980). Контакт провинций способствует проникновению не только отдельных видов растений, но и растительных сообществ. Анализ взаимоотношений между лесной и степной растительностью на территории парка предоставляет дополнительный материал к пониманию статуса лесостепи. На геоботанической карте (Карта растительности…, 1979) на территории парка выделено три контура: широколиственные леса, луговые степи и настоящие степи, которые на физикогеографической карте Саратовской области соответствуют таким ландшафтным структурам, как Хвалынские горы (широколиственные леса), и аккумулятивная долина реки Терешки, а также волжские террасы (различные типы степей, преимущественно луговые).
Таким образом, Приволжская возвышенность как крупный географический рубеж, протянувшийся в субмеридиональном направлении, влияет на изменение климатических условий с севера на юг и с запада на восток, перераспределяя баланс тепла и влаги и усиливая континентальность в восточном направлении. Благодаря этому в парке создаются своеобразные мезоклиматические условия: от умеренно-континентальных до резко континентальных (Макаров и др., 2006). Например, наветренный западный макросклон Хва-лынских гор и их водораздельный массив более увлажнены по сравнению с долиной р. Терешки и восточным макросклоном с
волжскими террасами, находящимися в ветровой тени. Соответственно, для восточного макросклона характерны повышенные температуры летнего сезона и преобладающие ветры восточных и юго-восточных направлений зимой, чего не наблюдается на западном макросклоне и водораздельном массиве, которые значительно прохладнее и лежат в зоне воздействия западных ветров. Благодаря указанным различиям индекс аридности меняется в пределах 1–1,41–1,67 (Бобровская, 2013).
Не только рельеф Приволжской возвышенности оказывает влияние на климат. Известно, что не менее значимую роль играет такой важный ботанико-географический рубеж, как долина р. Волга, выступающая в роли проводника лесной растительности далеко на юг. Также необходимо учитывать, что в волжской долине и на надпойменных террасах под влиянием континентально-тропического воздуха из Средней Азии и Ирана устанавливаются резко континентальные климатические условия, что приводит к распространению степей более южного варианта.
Возникает вопрос о динамике распределения лесного и степного типов растительности на Приволжской возвышенности. Для его решения важным является анализ структуры рельефа и растительного покрова в плане их исторического развития и в связи с геологическим прошлым. Геологическая история обуславливает существенные различия в строении смежных участков земной поверхности, форм рельефа, в литологическом составе, смене климатов, флор и растительности. В настоящее время имеются работы, освещающие этапы становления растительного покрова и динамику флоры в определенные периоды геологической истории на Юго-Востоке Русской равнины (Чигуряева и др., 1988; Завьялов и др., 2002; Благовещенский, 2003; Сенатор, Моров, 2016, 2017; Сенатор, 2017).
Настоящая публикация продолжает серию статей (Малышева, Малаховский, 2015; Малышева, 2016) о растительности национального парка «Хвалынский». Ранее на основе доминантно-детерминантного метода составлены классификации основных степных формаций (Stipeta pennatae и Stipeta capil-latae), рассмотрены их структурные особенности и разнообразие синтаксонов на уровне ассоциаций. По составу доминантов и фито-ценотипов установлена их принадлежность к южному, более ксерофитному варианту луговых степей. Выявлено, что настоящие степи имеют редкую встречаемость и не характерны для территории парка. Установлено преобладание кальцефитно-петрофитных вариантов степей в связи с карбонатными отложениями позднемеловой эпохи на Приволжской возвышенности. Показаны закономерности географического распространения фитоценозов, относящихся к ассоциациям степных формаций, обусловленные климатическими изменениями и формами рельефа.
Одна из задач настоящей статьи – выявление особенностей пространственной структуры фитоценозов лесной и степной растительности Приволжской возвышенности (Хвалынское Приволжье) с использованием палеогеографического метода. Номенклатура сосудистых растений приводится по сводке С.К. Черепанова (1995).
Этапы геологической истории Приволжской возвышенности
Приволжская возвышенность (Хвалын-ское Приволжье) в тектоническом отношении относится к восточному склону Сызран-ско-Терешкинского свода Воронежской ан-теклизы (Макаров и др., 2006; Национальный парк…, 2014). С восточной стороны свод ограничен крутым склоном глубокой грабенообразной долины р. Терешки и Приволжским прогибом, отделяющим его от Пугачев-ско-Чанаевского свода (Востряков, 1967). В составе Сызранско-Терешкинского свода находится Волго-Терешкинский горстообразный вал, ограниченный разломами вдоль рек Терешка и Волга. Хвалынское Приволжье – часть Приволжской возвышенности, в тектоническом отношении принадлежащая северо-западной окраине Жигулевско-Пугачевского свода. Это область относительно глубокого погружения кристаллического фундамента, перекрытого мощным чехлом палеозойских осадков, на которых лежат относительно молодые мезозойско-кайнозойские толщи, на поверхности представленные тонким разорванным плащом молодого песчано-глинистого делювия и вывет-релого щебенистого материала четвертичной системы. Общая мощность мезо-кайнозойских отложений не превышает 600 м и начало их формирования датируется байос-ским веком средней юры, когда произошла морская трансгрессия после длительного континентального перерыва. В раннемеловую эпоху продолжалось формирование морских осадочных отложений. Наиболее древними выходящими на поверхность являются отложения, соответствующие барремскому ярусу, состоящие из плотных темно-серых глин с прослоями алевритов.
Барремские глины перекрываются серыми ожелезненными глинами апта и затем почти черными глинами альбского яруса. Глинистые нижнемеловые толщи обнажаются редко. Они встречаются на приволжских склонах, на отметках ниже 150–100 м абсолютной высоты (Национальный парк…, 2014).
Отложения верхнего отдела меловой системы (100,5–66 млн. лет назад) относятся к морским и имеют для описываемой территории большее значение. Они представлены мел-мергелистыми толщами. Туронский ярус сложен серовато-белым песчанистым мергелем. Его сменяет сантонский, представленный грубым мергелем. Кампанская толща представлена глинистым мелом, маастрихтская – писчим мелом.
В меловой период, с точки зрения палеоботаников (Криштофович, 1930; Викулин, 1991; Жилин, 1991), в условиях влажного «оранжерейного» (т.е., теплого гумидного) климата господствовали субтропическая флора и растительность.
После небольшого континентального пе рерыва, привязанного к границе мел– палеоген, продолжилось формирование морских отложений. Они представлены нижним отделом – палеоценом. Выделяется сызранская свита, в настоящее время сопоставляе- мая с зеландским ярусом, для нее характерны кремниевые породы – опоки, переходящие в песчаные опоки и опоковидные песчаники (Макаров и др., 2006). Отложения, соответствующие лежащей выше по разрезу саратовской свите (сопоставляется с танетским ярусом) и представленные в лежащих севернее районах кварцевыми песками, содержащими прослои сливных песчаников, на описываемой территории не сохранились. Так завершилась относительно спокойная мезозойско-раннекайнозойская фаза морской аккумуляции, сформировавшая материал осадочных толщ Приволжской возвышенности.
В конце палеогена (эоцен-олигоцен) Восточно-Европейская платформа была охвачена тектонической перестройкой, связанной с глубокими структурными преобразованиями в альпийской геосинклинальной области, которые проявились в тенденции к нарастанию ее сводового поднятия. В миоценовую фазу происходило поднятие Воронежского выступа, а вдоль нынешней Волги формировалась полоса Саратовских дислокаций (Доскач, 1971). Дифференцированные движения земной коры определили формирование чередующихся возвышенных и низменных структур субмеридионального простирания (Дос-кач, 1971). Так поднялась Приволжская возвышенность, структурные особенности которой определены ее тектоникой и литологией.
К одной из зон наибольшей тектонической деформации относятся Хвалынские горы. Их субмеридиональная направленность определила асимметричность строения (западный и восточный макросклоны), а литологические особенности – ярусность и ступенчатость водораздельных поднятий. Вершины гор являются фрагментом верхней ступени выравнивания Приволжской возвышенности и сложены породами палеогенового возраста (пески, опоковидные песчаники). Главный водораздельный массив Хвалынских гор имеет высоты от 300 до 370 м, а наивысшие отметки достигают высоты 379 м.
На общем фоне поднятий на Юго-Востоке Русской равнины происходило отступание морей Понто-Арало-Каспийского бассейна.
Смена трансгрессивных и регрессивных фаз сопровождалась то расширением, то сужением площади морей. Сокращение водных пространств обусловило глобальное похолодание, что привело к началу распада субтропической (полтавской) флоры. Она еще длительное время (олигоцен) сохраняла свой облик, постепенно ведущий к развитию сава-ноидных типов, но в каждом регионе становление конкретных флор и фитоценозов имело свою специфику. Происходила смена субтропической флоры на умеренную (Криштофо-вич, 1934; Викулин, 1991; Камелин, 1995).
По мере похолодания формировались «саванны умеренной природы», на базе которых в дальнейшем могли вычленяться различные типы, например, сообщества, напоминающие травяные ярусы бывших саванн с отдельными более выносливыми древесными формами (из гелиофильных и опушенных). Эти комплексы испытывали автохтонную перестройку состава и структуры ценозов, в процессе которой происходило их существенное обновление за счет североголарктических элементов, что приводило к резкому всплеску видообразования. Появлялись новые виды, новые жизненные формы, хорошо адаптированные к изменяющимся условиям среды. Р.В. Камелин (1995) рассматривает возможность формирования безлесных пространств и становление степей на Русской равнине с конца миоцена, но не в комплексе с саванноидными типами, а уже с широколиственными лесами и с явно выраженными зональными особенностями. Этот взгляд на эволюцию растительности очень важен для понимания условий формирования и становления лесостепи. И.К. Пачоский (1910) на основе анализа флоры и ископаемых остатков растений допускает существование лесостепи в доледниковое время, при этом травяные и древесные сообщества – более или менее равноправные члены, занимающие определенные местообитания. Однако леса были «чем-то более древним – элементом лесостепи».
Ф.Н. Мильков (1952), вслед за И.К. Пачо-ским, признает древность лесостепи, полагая, что лесостепной ландшафт умеренного пояса представлял собой трансформированный в верхнем плиоцене саванный ландшафт. И.И. Спрыгин (1930) о средней части Приволжской возвышенности пишет как о «лесостепном районе, где фрагменты леса перемежаются со степью».
В результате можно предположить, что вслед за поднятием Приволжской возвышенности на границе палеогена и неогена произошло расширение лесостепного пояса Восточной Европы.
Для миоцена–плиоцена, с окончательной редукцией Понто-Арало-Каспийских бассейнов, исчезновением Тургайского пролива и последующей аридизацией климата, С.В. Викулин (1991) отмечает появление обширных безлесных пространств на территории Центральной России и Казахстана. Роль миграционных процессов в преобразовании флоры усиливается. Происходит перемещение не только видов, но и изменение растительности в целом. В.Л. Комаров (1921) отмечал, что именно с этого момента в Евразии сложились предпосылки к проявлению меридиональной зональности в составе флор и ценозов.
Миграционная флора Юго-Востока Русской равнины в этот период пополнялась и развивалась за счет гумидных и аридных центров (Коржинский, 1894; Лавренко, 1938). Так, Среднеевропейский и Южно-Уральский центры сыграли определенную роль в формировании флоры и растительности широколиственных лесов ( Quercus robur , Tilia cor-data , Corylus avellana и др.) и лугов европейской части России, вплоть до Урала. Из аридных центров более ощутимое влияние оказал восточносредиземноморский. Присутствие средиземноморских элементов проявляется на Юго-Востоке Русской равнины в составе растительности лесостепи, степи и особенно растительности меловых обнажений. Вероятно, нельзя исключать и роль более засушливых центров, таких, как Переднеазиатский и Арало-Каспийский. Так в Восточной Европе произошло широкое распространение теплоумеренной флоры и растительности, близкой к современной лесостепи.
Для рельефа Приволжской возвышенности в миоцен-неогеновый период наиболее характерными были эрозионноденудационные процессы. За счет денудации водоразделов в пределах которых ниже палеогенового уровня формировались ступени новой миоцен-неогеновой поверхности выравнивания (Доскач, 1971). Молодые подвижки платформы, как отмечает Е.В. Завьялов с соавт. (2002), приводили к глубокому врезанию древних рек. Отмирали старые ложбины стока, постепенно заполняясь осадками. Долины рек расширялись, создавая своеобразные долинные и межбалочные поверхности в виде полосы субмеридиональных равнин. Так, на Приволжской возвышенности формировалась густая и сильно разветвленная гидрографическая и балочная сеть, создавая морфоструктуры пестрого литологического состава.
Параллельно с денудационными процессами формировались трансгрессивные морские отложения (акчагыльское и апшеронское выравнивание) и связанные с ними абразионно-аккумулятивные террасы. Приволжская возвышенность в процессе всех преобразований к началу плейстоцена приобрела ступенчатый характер с четко обозначенными и гипсометрически неоднородными мор-фоструктурами. Ю.А. Мещеряков (1960) выделяет изогипсы 160–180 м, окаймляющие полосы водораздельных равнин, и изогипсы 200, 250, 300, 350 м, соответствующие поверхности выравнивания водораздельных поднятий Приволжской возвышенности. Подобные морфоструктурные особенности Приволжской возвышенности, по мнению А.Г. Доскач (1971), предопределили ее лесостепной характер, где древние морфострук-туры возвышенности относятся автором к морфоструктурам с господством лесных формаций, а морфоструктуры равнин, как более молодые, характеризуются доминированием степей. Уже в плиоцене становится очевидной связь типов растительности с определенными морфоструктурами.
В.В. Благовещенский (2003) считает, что в плиоцене на верхней ступени Приволжской возвышенности на песчаных и песчанокаменистых субстратах могли быть распространены сосновые боры, возможно, с примесью ели. В конце плиоцена в связи с миграцией флор на Приволжской возвышенности получили распространение широколиственные породы (дуб, липа, лещина), что в значительной степени способствовало становлению на средней ступени возвышенности сосново-широколиственных и чистых сосновых лесов. При разрушении верхней ступени, где обнажались верхнемеловые породы, могла возникнуть новая раса горных сосняков – «меловые» сосняки, которые встречаются и в настоящее время.
И.И. Спрыгин (1930) пишет, что «по отношению к лесной третичной области Приволжской возвышенности можно ставить вопрос и о значительной роли липы и березы, наравне с сосной и дубом». Что касается степных участков, то И.И. Спрыгин видит возможным их распространение по выровненным низинным междуречьям рек Суры и Волги, а со слов М.Н. Богданова, по ровным местам к югу от р. Сызрана – так называемая «Засызранская степь». Более развернутое описание растительного покрова Среднего Поволжья в плиоцене находим в работе С.А. Сенатора и В.П. Морова (2016).
Становится очевидным, что с момента распространения лесостепи на Приволжской возвышенности, лесной и степной тип растительности существовали рядом, занимая определенные формы рельефа (морфострукту-ры), перераспределяющие тепло и влагу на локальном уровне.
В плейстоцене Восточно-Европейская равнина испытала ряд оледенений. Приволжская возвышенность представляла собой перигляциальную область и геоморфологически развивалась во внеледниковых условиях. Основную роль в формировании рельефа играли эрозионно-денудационные процессы и аккумулятивное выравнивание депрессий. Значительное влияние на развитие этих процессов оказывали плейстоценовые изменения объема поверхностного стока, связанные с развитием и деградацией материковых оледенений.
Под воздействием флювиальных процессов окончательно сформировался рисунок гидросети. Морфоструктуры водораздельных поднятий оказались более устойчивыми к процессам денудации и эрозии, а межводораздельные равнинные пространства порой превращались в зоны активной аккумуляции (континентальной, иногда морской), формируя толщи рыхлых четвертичных отложений (Доскач, 1971).
Влияние на формирование морфоструктур возвышенности оказывали и небольшие тектонические движения платформы, совпадающие с этапами поднятий в ранне- и позднечетвертичное время (Мещеряков, 1960).
Находясь во внеледниковых условиях, Приволжская возвышенность не испытывала катастрофических изменений ландшафта. Климат эпохи оледенений в целом был суровым, но это не привело к полной гибели третичной доледниковой флоры и растительности (Пачоский, 1910; Мильков, 1952; Сенатор, Моров, 2017). Е.М. Лавренко (1938) первоначально считал Приволжскую возвышенность доледниковым рефугиумом лесостепного типа. В современных работах (Сенатор, Моров, 2016) Жигулевская возвышенность упоминается как рефугиум плиоценовой и ранне-плейстоценовой флоры. В.В. Благовещенский (2003) предполагал, что растительность на Приволжской возвышенности в течение всего плейстоцена сохранялась за счет преобладания хвойных видов. Растительность в эпоху оледенения испытывала автохтонную криоксерофильную перестройку. Обновление шло за счет криофильных, часто и более мезофильных элементов, попавших в условия более континентального климата при медленном смещении зон (Мильков, 1952; Камелин, 1995).
Эту точку зрения подтверждают результаты исследований самарских и саратовских ученых (Чигуряева и др., 1988; Сенатор, 2017; Сенатор, Моров, 2017), которые на основе анализа геоботанических, палеоботанических, палинологических и геологических данных с использованием параметрических показателей климата провели реконструкцию растительности и климата на Юго-Востоке Русской равнины в эоплейстоцене–голоцене. Не вдаваясь в подробности анализа споровопыльцевых комплексов из различных стратиграфических подразделений зональных типов растительности на территории Саратовской области (Чигуряева и др., 1988), остановимся на некоторых динамических аспектах климата, определяющих сочетание различных типов растительности. Палеогеографические события в апшероне–голоцене (продолжительностью около 2 млн. лет) представлены авторами в виде сводной таблицы. Ее данные не показали катастрофически холодных фаз климата, которые могли бы привести к гибели растительности. Тем не менее, климат и растительность менялись неоднократно и в разных масштабах. Например, за эоплейстоцен-плейстоцен палеоклиматические кривые показывают чередование пяти умереннохолодных и теплых фаз различной продолжительности. Им соответствовали 40 фаз, показывающих взаимоотношения лесной и степной растительности в лесостепи (степи рассматриваются вне их типологической принадлежности).
Умеренно-холодные и резко континентальные фазы (температура июля +12+14°, января –25–30°, гидроконтинентальность 2,5) имели место в периоды крупных трансгрессий и ледниковий и характеризовались в основном развитием перигляциальных и холодных (прохладных) степей при небольшом участии сосны (марево-полынно-сосновые комплексы) – так называемая холодная плейстоценовая лесостепь.
Отсутствие пыльцы злаков и других видов А.А. Чигуряева с соавт. (1988) связывают с перекрытием их пыльцой маревых и сложноцветных и большим участием пыльцы сосны. Это не позволяет достоверно реставрировать экологические условия развития перигляциальных степей. Однако по шкале гидроконтинентальности (ГК) можно предположить, что в перигляциальных степях доминировали степные ксерофиты вперемешку с арктоальпийскими формами (Лавренко, 1938).
Последнее похолодание, на границе плейстоцена и голоцена, было самым суровым (температура июля +10°, января –40°, ГК 2– 2,5). Оно сопровождалось двумя фазами: 1) перигляциальных холодных степей (полынно-маревый комплекс) и 2) тундровых и холодных степей (марево-полынный с участием березы, не исключено, что карликовой) – тундровая лесостепь.
А.К. Сытин (1997), анализируя отчеты по экспедиции П.С. Палласа в 1793 г. по Поволжью, пишет о его находке Betula nana в «холодных лесах» и болотах в окрестностях г. Пенза. Свидетельство Палласа можно рассматривать как аргумент в пользу признания реликтового происхождения Приволжских лесов и выделения Е.М. Лавренко (1938) «Приволжского центра консервирования межледниковых реликтов» (рефугиумы лесостепного типа).
Периоды потеплений, в отличие от похолоданий, были более продолжительными. Они наблюдались в межледниковья, когда климат был теплым (очень теплым) и влажным или полузасушливым (температура июля +20+22°, января –6°, ГК 1–2), создавая благоприятные условия для развития растительности. Прекращение ксеротермичности и последовавшее увлажнение привело к перестройке растительности. Холодная плейстоценовая и тундровая лесостепь претерпевает значительные изменения, с одной стороны, в сторону облесения, а с другой стороны – с заменой более мезофитными вариантами.
В выборе путей распространения леса оставляли открытыми низменные равнины, захватывая не целиком, а лишь частично расчлененные возвышенности и богатые грунтовой водой пески (Мильков, 1952). Это высказывание Ф.Н. Милькова очень важно для понимания роли геолого-геоморфологического фактора в распространении типов растительности в лесостепи и их связи с определенными морфоструктурными особенностями рельефа.
А.А. Чигуряева с соавт. (1988) для теплых периодов межледниковий отмечает широкое распространение растительности лесостеп- ного типа – отдельные участки сосновоберезовых лесов с участием ели и единичным присутствием термофильных пород – дуба, липы, вяза и др. Степные группировки характеризуются авторами как марево-полынные (маревые, злаки, кермеки, эфедра, скабиоза, полыни, гречишные и др.).
Можно предположить, что на Приволжской возвышенности (Саратовская область) в межледниковья господствовала хвойная лесостепь пестрого флористического состава; когда еще встречались растения моховых и лишайниковых тундр, болот, хвойных и лиственных лесов, разнотравных и сухих степей, солончаков и щебнистых склонов. Впоследствии, с отступлением ледника, позднеледниковая хвойная лесостепь продвинулась на север возвышенности (Мильков, 1952). В.В. Благовещенский (2003) также считает возможным распространение в плейстоцене на Приволжской возвышенности (Ульяновская область) сосновых лесов с участием ели.
В голоцене климат не отличался большими колебаниями, за исключением климатического оптимума (температура июля +20+22,5°, января –6–10°, ГК 1–1,4), который наблюдался 7,7–4,6 тыс. лет назад. Влажный и теплый климат оптимума, сменивший прохладный и полузасушливый (температура июля +16+18°, января –25°, ГК 1,4–2), способствовал быстрому распространению лесов с теплолюбивыми породами (дуб, липа, лещина и др.). Согласно Ф.Н. Милькову (1952), в период климатического оптимума, так называемый атлантический период, произошло максимальное облесение всего юга Русской равнины и началось распространение широколиственных лесов. При этом степи по-прежнему занимали огромные пространства, но, видимо, с некоторыми сменами. Е.М. Лавренко (1938) отмечает в их составе виды, характерные для более южных вариантов сухих ковылковых степей, как показатели распространения в ксеротермическую фазу послеледникового времени соответствующих типов степей в область, ныне занятую луговыми степями.
В начале субатлантического периода (3,2– 1,6 тыс. лет назад) произошло похолодание
(температура июля +16+18°, января –15°, ГК 1,4), которое приостановило «экспансию» лесов. Лесостепь с широколиственными лесами замещается лесостепью с березовососновыми лесами и единичной встречаемостью ели.
Во второй половине первого тысячелетия нашей эры произошла оптимизация климата. Палеоклиматическая кривая фиксирует «малый климатический оптимум» (температура июля +20+22°, января –10–13°, ГК 1–1,4), характеризующийся благоприятными сочетаниями тепла и влаги для развития лесостепи, с участием широколиственных лесов (березово-сосново-дубово-полынный комплекс), близкой к современной.
Представленная картина реконструкции, как отмечают сами авторы, не может претендовать на абсолютную достоверность и полноту, но позволяет сделать важные выводы. Основной вывод состоит в том, что в различные геологические отрезки апшерона– голоцена, несмотря на смену не менее 40 фаз растительности, авторами не выявлено доминирования исключительно лесного или степного типа растительности. Лесные и степные фитоценозы присутствовали на всех этапах развития растительного покрова, не сменяя друг друга, менялось лишь соотношение их комплексов и состав биоморф. Все это свидетельствует об устойчивом равновесии этих типов растительности в лесостепи Юго-Востока Русской равнины. На самобытность лесостепного ландшафта, в котором лесные и степные растительные фитоценозы четко маркируются и существуют как равноправные и независимые друг от друга, указывали многие геоботаники (Пачоский, 1910; Крашенинников, 1937; Мильков, 1952; Зозулин, 1958; Дохман, 1960; Лавренко, 1980 и др.). Однако существовали и продолжают существовать другие мнения о том, что степи в современной лесостепи существуют на месте бывших лесных территорий, или что лесостепь представляет собой различные стадии завоевания лесом степных территорий.
Из краткого обзора сведений о становлении и развитии лесостепи на Юго-Востоке
Русской равнины Приволжской возвышенности (Хвалынское Приволжье) следует, что распространение лесного и степного типов растительности четко определяется морфоструктурными особенностями рельефа возвышенности. Пространственное соотношение этих типов на протяжении всей геологической истории оставалось достаточно устойчивым. Следовательно, о лесостепи средней части Приволжской возвышенности можно говорить как о коренной растительности.
С овременная лесостепь
Особенности современного растительного покрова лесостепи Приволжской возвышенности рассмотрим на примере национального парка «Хвалынский». Они обусловлены спецификой геоморфологического и литологического строения возвышенности. С одной стороны, это возвышенные водоразделы (Хвалынские горы – 379 м), сложенные мезо-раннекайнозойскими отложениями (глины, мел, мергели, опоки, пески, песчаники), с другой – относительно молодые аккку-мулятивные равнины долины р. Терешки (160–180 м), сложенные суглинками и опес-чаненными глинами нижнего отдела меловой системы, перекрытыми четвертичными отложениями.
Хвалынские горы, простираясь вдоль Волги более чем на 70 км, перераспределяют баланс тепла и влаги, создавая систему мезоклиматов (Жуков, 1971): умеренноконтинентального – континентального – резко-континентального (восточный макросклон Хвалынских гор). Геоморфологические и литологические особенности территории обусловливают пестроту почвенного покрова. На поднятиях в местах обнажения и близкого залегания обломочных пород грубого механического состава с хорошей водопроницаемостью формируются почвы черноземного и лесного типов. На палеогеновых песках встречаются серые лесные супесчаные почвы, на выходах мела и опок формируются хрящеватые и щебенчатые черноземовидные с неполноразвитым профилем и карбонатные черноземы (Болдырев, 1997).
Подобные грубые субстраты Хвалынских гор и других останцовых поднятий в условиях умеренно-континентального климата создают благоприятные условия для распространения и развития лесной растительности. Ф.Н. Мильков (1952) справедливо заметил, что леса оставляют открытыми низменные, плоские равнины и захватывают лишь частично расчлененные возвышенности, что и наблюдается на территории национального парка.
Иные условия на пологих западных склонах водораздельных поднятий, переходящих в долину р. Терешка. Здесь широкое распространение получили делювиальные лессовые черноземами обыкновенными, которые характеризуются мелкоземистой структурой и трудной проницаемостью воды (Усов, 1948). По восточному макросклону Хвалынских гор и волжским террасам, где породы сильно размыты и покрыты глинистым делювием, встречаются черноземы обыкновенные, чаще маломощные и черноземы южные с домини рованием степного типа растительности. Та ким образом, в национальном парке наблюда ется закономерное чередование лесов и степей (рис.). Пространственное распределение каждого из этих типов растительности определяется морфоструктурными особенностями территории и достаточно устойчиво в глины и суглинки с нормально развитыми пространстве.
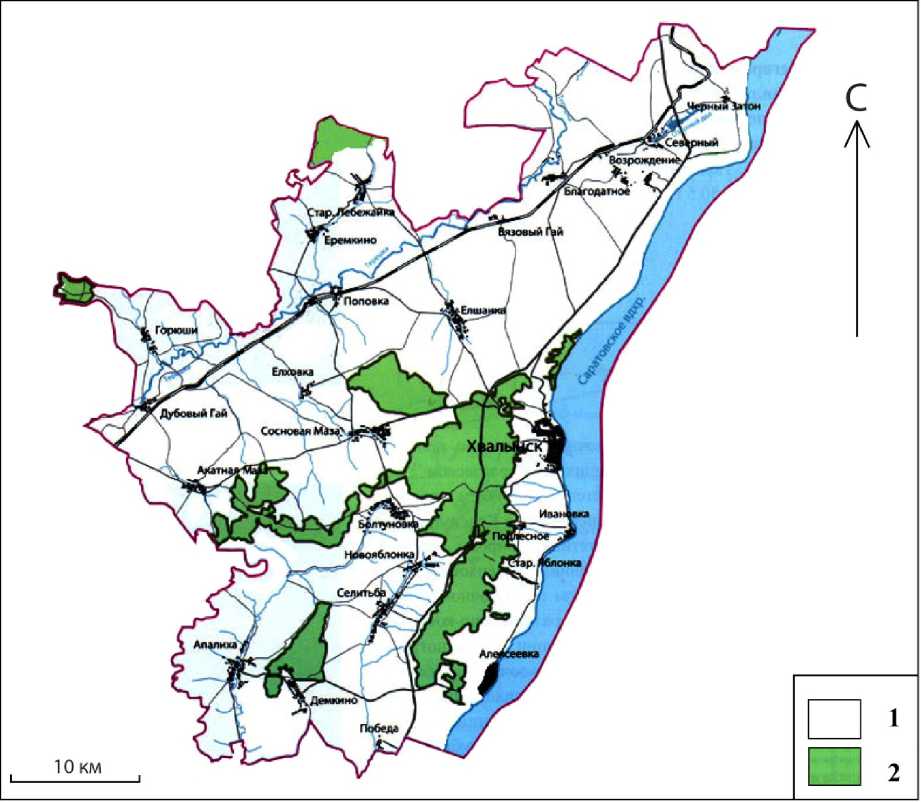
Рис. Рельеф и типы растительности Приволжской возвышенности: 1 – степи на равнинах; 2 – леса на возвышенностях
The topography and the vegetation type of the Volga Upland: 1 – steppes of plain; 2 – forests of upland
Лесная растительность парка изучена довольно подробно, в связи с чем ниже приведены масштабы ее площадей, спектр формаций и взаимоотношения со степной растительностью.
Общая площадь продуктивных лесов парка составляет 22931 га (Национальный парк…, 2014), что составляет около 16 % от площади Хвалынского района. В типологическом отношении это восточно-европейские широколиственные леса в переделах Восточноевропейской лесостепной провинции (Исаченко, 1980). Они отличаются от западных вариантов положением в более континентальных условиях, некоторой флористической бедностью за счет сокращения европейских неморальных видов. Основными лесообразующими породами являются Tilia cordata , занимающая 32,2 % земель, покрытых лесной растительностью, Quercus robur – 30,9 %, Pinus sylvestris – 26,1 %. На остальные породы ( Acer platanoides , Ulmus laevis , U. glabra , Populus tremula , Betulapubescens, ) приходится 10,8 % (Кабанов, 2014).
Известно, что в голоцене до начала хозяйственной деятельности в средней части Приволжской возвышенности господствовали сосновые и широколиственно-сосновые леса (Кузнецов, 1928; Спрыгин, 1930; Благовещенский, 1971; Чигуряева и др., 1988; Сенатор, 2017). Н.И. Кузнецов (1896) при этом выделяет особую роль дуба, а И.И. Спрыгин (1930) обращает внимание на участие липы наравне с дубом и сосной.
В настоящее время коренные сосняки встречаются небольшими участками среди массивов лиственных лесов в основном по вершинам водоразделов, на песках и по меловым склонам (Антонова, 1957; Тарасов, 1991; Серова, Березуцкий, 2008; Архипова, 2009; Кабанов, 2014 и др.).
Е.А. Архипова (2009), выделяя формацию сосны, диагностирует в ней 8 ассоциаций, из которых три представлены простыми сосняками (ландышевый, мертвопокровный и ор-ляковый), а 4 сложными, во втором ярусе которых участвуют Quercus robur, Tilia cordata, Acer platanoides. Достаточно структуриро- ванными среди них являются сосняки лазур-никовый и купеновый.
Фитоценотическое разнообразие сосняков описано С.В. Кабановым (2014), который отмечает устойчивость и разновозраст-ность ценопопуляций сосны, что свидетельствует о полном обороте возрастных поколений в сосняках. По мнению автора, отсутствие пирогенного воздействия за последнее десятилетие благоприятно сказывается на ходе их сукцессий.
Исследования по изучению взаимоотношения леса и степи показали, что сосна – единственная из древесных лесообразующих пород на территории парка, которая выходит в степь, образуя разновозрастные группировки (Малышева и др., 2011).
Определенный интерес в связи с ограниченной встречаемостью представляют сосновые леса на выходах мела – так называемые «меловые боры» с особой формой сосны (Дохман, Любимова, 1971). В них практически отсутствуют подрост и всходы, а в травяном покрове преобладают кальцефилы. Такие леса распространены по меловым обнажениям Хвалынских гор. Е.М. Лавренко (1938) называет их третичными реликтами и исчезающей формацией.
Основными лесообразующими породами лиственных лесов парка являются Quercus robur и Tilia cordata. Среди лесов преобладают дубравы. В.А. Болдырев (2005) называет приволжские дубравы на карбонатных почвах уникальным явлением Приволжской возвышенности. В.В. Благовещенский (1971) считает современные дубравы производными от сосново-широколиственных лесов. В настоящее время в парке преобладают низкоствольные дубовые леса, в большинстве случаев порослевые. По оценке С.В. Кабанова (2014), их древостой четвертой–пятой генерации. В пологе подроста обычно участвует липа мелколистная и клен платановидный. Часто в подобных дубравах к 50–60 гг. XXI в. за счет интенсивного вегетативного размножения липа может вытеснить дуб и возобновляться порослью. Так происходит смена эдификато-ров. Семенное возобновление в дубравах почти отсутствует, самосев единичен (Болдырев, 1991). Площади дубрав постоянно сокращаются в связи с периодически повторяющимися волнами массового усыхания. Граница дубрав со степной растительностью, в отличие от сосняков, достаточно четко обозначена. Нам не удалось встретить в степи ни дуба, ни видов растений, встречающихся в травянистом ярусе дубрав. Напротив, зафиксировано проникновение вглубь дубравы на расстояние 10–20 м степных видов (Bromopsis riparia, Festuca valesiaca, Stipa capillata, S. pennata, Filipendula vulgaris, Securigera varia, Medicago falcata и др.). Подобное проникновение степных видов в лес влияет на изменение экологической обстановки, что, естественно, приводит к остепнению леса.
Сложные дубравы на серых лесных почвах встречаются крайне редко. Они отличаются от производных флористическим богатством травостоя и сложной структурой древостоя. Флористическое и фитоценотическое разнообразие дубрав парка достаточно подробно охарактеризовано в работе Е.А. Архиповой (2009), которая в формации дуба выделяет 6 групп ассоциаций и 14 ассоциаций. Наиболее распространенной ассоциацией является дубрава ландышевая. Довольно часто в подобных ассоциациях отмечено участие в древостое Malus praecox . Вообще участие плодовых деревьев и кустарников в дубравах парка встречается достаточно часто.
Ранее отмечено, что одной из основных лесообразующих пород парка наряду с дубом является липа мелколистная, которая принимает участие в структуре всех типов леса (Макаров и др., 2006; Кабанов, 2014). Липовые леса, так же как и дубравы, являются производными сложных сосняков.
Широкое распространение липы С.В. Кабанов (2014) объясняет ее способностью к вегетативному размножению. Ценопопуля-ции липы, как правило, разновозрастны. Благодаря этому свойству липа на сегодняшний день усиливает позиции в парке, в отличие от дуба, что, вероятно, может привести к окончательному выпадению дуба.
В липняках кроме основного эдификатора, обычно присутствуют дуб и клен платанолистный. Наиболее распространенными типами сообществ в парке являются липовокленовые и кленово-липовые, в меньшей степени распространены дубово-липовокленовые (Антонова, 1957).
Е.А. Архипова (2009) провела синтаксо-номический анализ липняков парка и выделила 10 ассоциаций с разной долей участия дуба и клена. Среди них наибольший интерес представляет липняк волосистоосоковый, как наиболее представительный по участию липы в древостое (8Лп, 2Кл плат). Чаще встречаются липняки с участием клена и дуба и подроста этих пород, с подлеском из бересклета, жимолости, орешника. Травостой представлен лесными видами ( Convallaria majalis , Stel-laria holostea , Aegopodium podagraria и др.). Липняки приурочены в основном к теневым участкам западного макросклона.
Исследования взаимоотношения липняков со степной растительностью позволили выделить по их границе экотонную полосу, в которой наблюдается смена нескольких экологических рядов. Наличие экотонной полосы вокруг липняков, как более мезофитных типов леса, отличает их от ксерофитных сосняков и дубрав. Влияние леса прослеживается в пределах не менее 10–15 м.
Достаточно сложно в лиственных лесах парка выделить кленовую формацию, где эдификатором выступал бы Acer platanoides . Тем не менее, Е.А. Архипова (2009) выделяет эту формацию и диагностирует в ней ассоциации. Чаще всего встречаются леса с совместным доминированием или содоминиро-ванием в составе клена и липы. Древостои с кленом предпочитают световые склоны.
Осинники (Populus tremula) встречаются в различных местообитаниях, но при отсутствии избыточного увлажнения. Самые высокопродуктивные насаждения со вторым ярусом (липа, клен, вяз шершавый) и преимущественно кленовым подростом встречаются в плакорных условиях на супесчаных и суглинистых почвах (Кабанов, 2014). Осинники теневых склонов представляют более обед- ненный вариант. Большинство осинников при естественном развитии замещается кле-новниками и липняками. Е.А. Архиповой (2009) при исследовании осинников выявлено небольшое их фитоценотическое разнообразие (3 ассоциации).
Березовые леса в парке, как и осинники, встречаются редко (Архипова, 2009) и чаще формируют обособленные массивы. В березовой формации выделяется 7 ассоциаций. Сопутствующими породами в березняках обычно выступают липа и клен. Их возраст, как правило, не превышает 60–80 лет. Исследования взаимоотношения березняков со степной растительностью (Малышева и др., 2011) позволили выявить экотонную полосу, ширина которой зависит от типа леса. Например, в случае с кленово-березовым лесом орляковым его влияние проявлялось на расстоянии 25–30 м, в связи со сменой экотопов от влажного к свежему и суховатому. Смена экотопов выражалась в смене экологических рядов: 1 – орляковый с участием неморальных видов ( Asarum europaeum , Asperula odo-rata , Primula macrocalyx ); 2 – кустарниковый ( Cerasus fruticosa , из трав Anemone sylvestris , Lathyrus pisiformis , Bromus inermis , Melica nutans и др.); 3 – высокотравный ( Filipendula vulgaris , Dracocephalum ruyschiana , Geranium sylvaticum , Stachys sylvatica , Thalictrum minus и др.). В другом варианте сосново-березового леса с доминированием в травостое Laser tri-lobum и Polygonatum odoratum на дерново-карбонатной свежей почве выявлена экотон-ная полоса в радиусе 2–3 м с участием купены.
Незначительную площадь занимают ольшаники. Е.А. Архиповой (2009) при исследовании этих типов леса удалось выделить только одну ассоциацию – ольшаник таволговоосоковый ( Alnus glutinosa – Carex acutiformis + Filipendula ulmaria ).
При изучении лесной растительности парка не удалось выявить случаев наступления леса на степь. Это еще раз подчеркивает, что лес и степь исключают друг друга на одной и той же территории. Лес предпочитает воз- вышенные эродированные водоразделы, а степь – равнинные.
Степная растительность в национальном парке занимает, в основном, территорию его охранной зоны (рис.). Это безлесные равнинные водоразделы р. Терешки и Волги. Общая площадь степей превышает лесопокрытую в 5 раз и составляет 114000 га, в том числе пашни и залежи.
В типологическом отношении господствуют луговые степи, характеризующиеся определенным набором эдификаторных вариантов южного зонального подразделения лесостепи (Исаченко, Лавренко, 1980). Настоящие степи – разнотравно-ковыльные и разнотравно-ковылковые – имеют фрагментарное распространение. Состояние луговых степей в настоящее время, в связи со значительным снижением антропогенной нагрузки и прекращением выпаса, можно оценить как средней степени нарушенности, за исключением припоселковых земель. Учитывая высокую семенную продуктивность луговостепных видов растений, степи парка достаточно быстро восстанавливаются.
Флористический состав степей достаточно богат и разнообразен (Серова, Березуцкий, 2008), а видовая насыщенность сообществ составляет около 200 видов сосудистых растений (Малышева, Малаховский, 2015; Малышева, 2016). Эти виды имеют различные ареалы, происхождение, биоморфологиче-скую структуру и пестрый эколого-фитоценотический состав. Среди них сохранился ряд эндемичных и реликтовых видов – Koeleria sclerophylla , Centaurea carbonata , Astragalus zingeri , Globularia punctata , Thymus cimicinus , Asperula exasperata , Potentilla volga-rica (узколокальный эндемик Приволжской возвышенности), Onosma volgensis и др. и большая доля видов, вошедших в Красную книгу Саратовской области (2006) со статусом «исчезающие», «уязвимые» и «редкие». В этой связи луговые степи парка можно рассматривать как реликтовые, виды которых, особенно петрофитного комплекса, сохранились с миоцен-плиоцена (Лавренко, 1938).
Основными господствующими злаками степей (в широком смысле) являются плот-нодерновинные ковыли – Stipa pennata , являющийся к тому же основным доминирующим зональным мезоксерофильным видом луговых степей и Stipa capillata , встречающийся во всех типах степей (эвриксерофил), нередко доминирующий. В отличие от первых двух ковылей, изредка встречается эврик-серофил Stipa lessingiana. Кроме ковылей содоминирующим видом из злаков является длиннокорневищный Bromopsis riparia. Значительно меньшее участие в структуре сообществ степей принимают другие злаки. Из плотнодерновинных это эвриксерофилы Fes-tuca valesiaca, Koeleria cristata , из рыхлодер-новинных – Phleum pleoides (ксеромезофил), а из корневищных – Poa angustifolia , Bromopsis inermis , Calamagrostis epigeios , Elytrigia repens (эвримезофилы).
Разнотравье луговых степей отличается богатством и полидоминантностью, формируя сложные структуры многоярусных сообществ с ОПП – 80–100%, разного биоморфо-логического и эколого-фитоценотического состава.
Кустарники в луговых степях не так многочисленны ( Cerasus fruticosa , Chamaecytisus ruthenicus , Genista tinctoria , Amigdalus nana ), но некоторые из них играют определенную фитоценотическую роль, как Chamaecytisus ruthenicus и Genista tinctoria , выступая в качестве доминантов и содоминантов некоторых ассоциаций ( Stipeta pennatae–Genistosa tinc-toria–Pluriherbosa; Stipeta pennatae– Chamaecytisusosa– Pluriherbosa; Stipeta pen-natae–Stipetosa capillata–Pluriherbosa ).
Как правило, в сообществах преобладают травянистые многолетники лугово-степного эко-фитоценотипа (ксеромезофиты и мезоксерофиты), составляющие более 50% от видового состава. Основное флористическое ядро представляет доминантная группа и группа характерных видов (Stipa pennata, S. capillata, Bromopsis riparia, Poa angustifolia, Genista tinctoria, Medicago romanica, Salvia testuicola, S. nutans, Securigera varia, Origanum vulgare, Adonis vernalis, Anemone sylvestris и др.).
Лесолуговые мезофиты менее многочисленны ( Elytrigia repens , Calamagrostis epigeios , Silene nutans , Thalictrum simplex , Asparagus officinalis и др.), так же как и настоящие степные ксерофиты ( Stipa capillata , Festuca valesiaca , Koeleria cristata , Artemisia austriaca , Galatella villosa , Astragalus testicula-tus и др.). Однако на восточном макросклоне Хвалынских гор их состав увеличивается за счет эвксерофильного злака Stipa lessingiana и ксерофильных эфемероидов Poa bulbosa и Pastinaca clausii . Последний вид заслуживает особого внимания, как вид периодического доминирования исключительно в засушливые годы. Из полукустарничков следует отметить эвксерофилы (пустынно-степные и пустынные виды) Kochiaprostrata, Krascheninnikovia ceratoides, Artemisia santonica .
Роль однолетников ( Acinos arvensis , An-drosace elongata ) в луговых степях парка незначительна, хотя Acinos arvensis имеет 100 % встречаемость.
На карбонатных почвенных разностях встречаются кальцефильно-степные виды, нередко выступающие в роли доминантов и содоминантов. К ним относятся преимущественно полукустарнички Hedysarum grandiflo-rum , Linum ucranicum , Onosma volgensis , Thymus cimicinus и травянистый многолетник Globulariapunctata .
Состав луговых степей на супесчаных черноземах богат псаммофильно-степными видами ( Carex supina , Helichrysum arenarium , Potentilla arenaria , Gypsophila paniculata , Hy-lotelephium stepposum , Koeleria glauca , Ono-brychis arenaria , Artemisia marschalliana и др.). Преобладающее число видов луговых степей парка является длительновегетирующими растениями с максимумом цветения во второй половине июня и первой половине июля. В это же время в луговых степях парка наблюдается экологический оптимум их развития.
Флористическое богатство степей парка определяет их фитоценотическое разнообразие. В процессе классификации (Малышева,
Малаховский, 2015; Малышева, 2016) в степ- ном типе растительности выделено два подтипа: луговые и типичные степи и 4 формации: Stipetapennatae, Stipeta capillatae, Stipeta lessingianae и Mixherboreta.
В луговых степях выделены 33 ассоциации, относящиеся к 3 формациям: перистоко-выльников, тырсовников и разнотравным (Mixherboreta), а в настоящих степях – 3 ассоциации в 2 формациях: тырсовников и ко-вылковой. Экологические ареалы формаций приурочены к определенным местообитани- ям.
Несмотря на широкое распространение
Stipa pennata в степях парка, фитоценозы пе- ристоковыльной формации занимают небольшие площади и сосредоточены в непосредственной близости от лесных массивов.
Формация на основе сходства эдификатора, группы
доминантов
( Chamaecytisus
ruthenicus , Genista tinctoria , Bromopsis riparia , Poa angustifolia , Adonis vernalis , Anemone syl-vestris , Paeonia tenuifolia и др.) объединяет 12
зональных ассоциаций, 3 их петрофитных варианта ( Linum ucranicum , Hedysarum grandi-florum , Globularia punctata ) и 2 псаммофит-ных ( Carex supina , Atremisia marshalliana ). В ассоциациях этой формации наряду с группой характерных видов ( Bromopsis riparia , Poa angustifolia , Adonis vernalis , Salvia tesqui-cola и др.) высокая константность отмечена и у Stipa capillata .
Тырсовая формация, в отличие от перистоковыльной, имеет более широкое распространение. Обширность ее ареала и его широкая экологическая амплитуда объясняют фитоценотическое разнообразие тырсовни-ков. Данную формацию в парке представляют 7 зональных ассоциаций ( Bromopsis riparia , Silene exaltata , Origanum vulgare , Stipa pennata , Salvia nutans , Genista tinctoria , Fes-tuca valesiaca ), 6 их петрофитных вариантов ( Iris pumila , Globularia punctata , Hedysarum grandiflorum , Thymus cimicinus , Linum ucranicum , Medicago romanica ) и один псам-мофитный ( Carex supina ).
По видовому составу ассоциации данной формации проявляют значительное сходство с перистоковыльной, которое проявляется, в основном, по эдификаторной синузии, представляющей типичный вариант южных луговых степей. В результате наблюдаются близнецовые ассоциации в перистоковыльной и тырсовой формациях (Малышева, 2016).
В качестве соэдификатора и одновременно доминанта в ассоциациях тырсовой формации нередко участвует Stipa pennata , что приводит к абсолютному доминированию двух ковылей. Можно предположить, что происходит внедрение Stipa capillata в перистоковыльные степи. Эту гипотезу в свое время высказала Г.И. Дохман (1954) при изучении растительности Мугоджар.
Ассоциации типчаково-тырсовой группы формаций не типичны для степной растительности парка из-за их фрагментарной встречаемости. Они приурочены к вершинам увалов восточного макросклона и представлены разнотравно-типчаково-тырсовой ассоциацией ( Poa bulbosa , Artemisia austriaca , Thymus marschalianus , Astragalus testiculatus , Gypsophila altissima , Oxytropispilosa , Galatella villosa , Pimpinella tragium и др.).
Разнотравная формация имеет узкий ареал по границе с лесом на западном макросклоне Хвалынских гор. Ее характеризуют полидо-минантные сообщества, в которых группа доминирования представлена в основном лесолуговыми мезофитами и лугово-степными мезоксерофитами ( Origanum vulgare , Salvia tesquicola , Medicago falcata , Thalictrum simplex , Securigera varia , Galium verum , Vince-toxicum hirundinaria , Filipendula vulgaris и др.). Из злаков встречаются Stipa pennata, S. capillata , Bromopsis inermis , Calamagrostis epigeios , Elytrigia repens и др. Наиболее характерные ассоциации: злаково-дроковоразнотравная и злаково-душицево-разнотравная .
Сообщества ковылковой формации представлены фрагментарно на террасах восточного макросклона Хвалынских гор, где встречаются южные черноземы. Формация объединяет 2 ассоциации: разнотравно-типчаково-ковылковую и разнотравно-терескеново-ковылковую и ее петрофитный вариант – тырсово-ковылковую. Характерными видами являются типично степные, пустынно-степные и пустынные виды (Poa bulbosa, Galatella villosa, Achillea nobilis, Fes-tuca valesiaca, Koeleria cristata, Astragalus tes-ticulatus, Kochia prostrata, Krascheninnikovia ceratoides, Atraphaxis replicata, Artemisia san-tonica и др.) и усиливается роль кустарничков и полукустарничков. В засушливые годы в качестве факультативного доминанта выступает Pastinaca clausii. Большинство сообществ являются редкими и нуждаются в охране.
Процесс развития древней палеогеновой суши Приволжской возвышенности во многом объясняет современную картину пространственной структуры растительности лесостепи, где лес и степь как типы растительности достаточно устойчивы в пространстве и существуют независимо друг от друга. В Хвалынском Приволжье они представлены достаточно масштабно и фитоценотически разнообразно. Отдельные деревья могут встречаться в степи, но это еще не лес. Для их внедрения необходима существенная смена экологической обстановки.
Заключение
Взаимоотношения леса и степи на Приволжской возвышенности рассмотрены в соответствии с ее геологической историей, начиная с морских фаз мезозоя – раннего кайнозоя, сформировавших значительные толщи морских осадочных отложений. Относительно спокойный мезозой сменился тектонически беспокойным кайнозоем, когда в палеогене поднялась Приволжская возвышенность. Все это за многие миллионы лет происходило на фоне смены субтропических флор и растительности Восточно-Европейской равнины умеренными. Последние не пострадали катастрофически в период ледниковых оледенений плейстоцена, сохранив облик, близкий к современному.
Литологические и тектонические особенности территории привели к формированию структуры рельефа Приволжской возвышенности, в котором разновозрастные морфост- руктуры предопределили лесостепной характер ее растительности (лес – степь).
К древней палеогеновой морфоструктуре относятся водораздельные поднятия (Хва-лынские горы), где на поверхность выходят пески, песчаники и опоки палеоцена. Грубый механический состав этих горных пород с хорошей водопроницаемостью и сформированными лесными и черноземовидными – неполноразвитыми и скелетными почвами создает благоприятные условия для развития и распространения лесной растительности.
К молодой четвертичной морфоструктуре относится аккумулятивная равнина долины р. Терешки, переходящая в пологую нижнюю часть западного макросклона Хвалынских гор. Здесь на аллювии с мелкозернистой структурой и слабой водопроницаемостью развиты дерновые аллювиальные и обыкновенные черноземы со степной растительностью. Господствующим типом степей в парке являются луговые степи, а настоящие степи имеют фрагментарное распространение по вершинам увалов и террас восточного макросклона, где на сложном глинистом делювии сформированы с полно развитым профилем обыкновенные и южные черноземы.
Проведенные исследования свидетельствуют о том, что лесной и степной типы растительности на Приволжской возвышенности, испытавшие длительную автохтонную перестройку, имеют относительно устойчивое соотношение в пространстве, где лес и степь являются равноправными членами лесостепного ландшафта зоны лесостепи.
Автор выражает признательность и благодарность директору национального парка «Хвалынский» канд. с.-х. наук В.А. Савинову и с.н.с. Г.Ф. Сулеймановой за содействие и помощь при проведении экспедиционных работ на территории парка. Автор также признателен бывшему научному сотруднику Отдела геоботаники БИН РАН П.Д. Малаховскому за участие в полевых работах.
Работа выполнена в рамках государственного задания согласно тематическому плану
Список литературы Взаимоотношения леса и степи на Приволжской возвышенности (в порядке дискуссии)
- Антонова Л.А. Краткая характеристика лесов хвалынского лесхоза и основные закономерности их распределения. Учен. зап. СГПИ. Саратов, 1957, вып. 28, с. 225-247.
- Архипова Е.А. Фитоценотический состав и структура лесной растительности национального парка «Хвалынский». Автореф. дис. … канд. биол. наук. Саратов, 2009, 20 с.
- Благовещенский В.В. Лесная растительность центральной части Приволжской возвышенности. Автореф. дис. … д-ра биол. наук. Пермь, 1971, 35 с.
- Благовещенский В.В. Современная растительность Приволжской возвышенности как отражение ее прошлой истории. Растительный мир Среднего Поволжья: Сб. статей. Сер. «Природа Ульяновской области». Ульяновск: УлГТУ, 2003, с. 3-15.
- Бобровская Н.И. Климатические рубежи основных подзональных типов восточноевропейских степей. Наука в Центральной России, 2013, № 2, с. 6-12.
