Water stress at anthesis and storage temperature affected growth and germinability of rice (Oryza spp.)
")
Author: Dada Oyeyemi A., Okpe Jude A., Togun Adeniyi O.
Journal: Журнал стресс-физиологии и биохимии @jspb
Article in issue: 1 т.16, 2020.
Free access
A study conducted in 2017 and 2018 investigated components of yield (CY) from upland rice plants predisposed to water stress at anthesis and germinability indices (GI) of harvested seeds stored at different storage temperatures. Sixteen treatments comprising four rice cultivars: CG-14, NERICA2, NERICA4 and FARO-66 predisposed to four periods of water stress [5, 10, 15 days interval and 100% Field capacity (control)] at grain filling were laid out in a completely randomized design (CRD) with six replicates. Morphological and yield parameters and GI were evaluated following standard procedures. Data were analyzed using linear regression and ANOVA with means separated using Duncan’s multiple range test (p≤0.05). Occurrence of water stress at grain filling had deleterious effects on CY and GI of seeds of FARO-66 cultivar. NERICA4 and CG14 tolerated moisture stress better than NERICA2 and FARO-66. Sterility, unfilled grain and GI are strongly (R2=0.87) related to prolonged drought incidence. Occurrence of drought at generative growth stage in rice could cause total crop failure and poor seed germinability. Considering the unpredictability of rainfall season in rainfed farming system, irrigation plan should be essential cultural practice that must be factored into upland rice production to avoid crop failure.
Moisture stress, upland rice varieties, storage temperature, seed germinability
Short address: https://sciup.org/143171153
IDR: 143171153
Text of the scientific article Water stress at anthesis and storage temperature affected growth and germinability of rice (Oryza spp.)
Water availability to field crops constitutes critical role at ensuring food security among many abiotic variables. Limiting water conditions due to unpredictable climatic conditions is expected to reduce productivity of cultivable field in the next five decades (Adejare and Umebese 2008, Be et al. 2014, Mancosu et al. 2015). Water stress, resulting from water deficit, affects plant growth, metabolism and physiology through alteration of physical environment. Almost every plant processes are held back directly or indirectly by insufficient water supply. The physiological responses of plants to moisture deficit and their relative importance for crop productivity vary with species, soil type, nutrients and ecology (Adejare and Umebese 2008, Be et al. 2014, Mancosu et al. 2015). Equally, drought induced loss in crop yield may exceeds loses from all other abiotic causes (Steduto et al. 2012). With the occurrence of water deficits many of the physiological processes associated with growth are affected and under severe deficits, death of plants may result (Farooq et al. 2009). The effect of water stress may vary with variety, degree and duration of water stress and the growth stage of the rice crop (DOASL, 2006). Water stress during vegetative stage retards plant height, tiller number and leaf area development in rice (Samarah and Alqudah 2011). However, the effect during this stage varies with the severity of stress and age of the crop. Drought stress during reproductive stage reduced photosynthetic productivity in barley (Masoud et al. 2005), poor floral development in Arabidopsis (Su et al. 2013) and increased grain sterility by 83% resulting in significant yield loss in upland rice (Dada et al. 2018).
More than three billion of the world’s population relies on rice as an important staple food (Chauhan et al. 2017). In 2017, approximately 165 million hectares were harvested worldwide which makes it a valuable commodity that influences livelihood and economies of several billion people (Fuhrmann et al. 2018). Nigeria is facing serious economic water scarcity (Molden 2013) and this is impinging on upland rice production. The economic water scarcity condition is taking its toll on upland rice production whereby drastic yield reduction per unit area of about 25% had been reported under severe water deficit condition (Zhang et al. 2018). The shortage of water in many rainfed rice growing communities may account for the wide discrepancy between demand and production of the crop. Total yield loss due to severe drought may even occur in some cases. It has been hypothesized that drought has indirect effect on germination and vigor of seeds harvested from crops affected by drought. It is not certain if the accelerated loss in viability of rice after harvest is related to drought at the reproductive phase of its development. Samarah and Alqudah (2011) indicated that barley plant predisposed to drought at the post anthesis had poor seed germination and low seedling vigor. Another report by Yaklich (1984) showed that moisture stress had no significant influence on emergence of soybean seed when stressed at seed formation stage.
Rice grains loose viability shortly after harvest and this affects seed production and storage which ultimately affects food security. Subsistence rice growers which dominate rice production in many developing countries, retain seeds from their last harvest for the next planting season. Unfortunately, most of these stored seeds loose viability shortly after storage thus leading to poor emergence, vigour and eventual establishment (Ahmad et al. 2009). Farmers being conscious of this menace, sow more seeds per stand beyond the recommended seed rate to cover up for the likely poor viability. Loss of viability in seeds is linked mostly to environmental and biochemical factors (Chen and Arora 2011). Water limiting condition during anthesis is a major factor that impinges adverse influence on seed development (Mut et al. 2010, Sun et al. 2011). Researches had been carried out on how water deficit, affected growth dry matter and yield of rainfed rice (Lilley and Fukai 1994, Ye et al. 2012) incidentally; these were related to Asian and European environment which is quite different from tropical conditions. Besides, many of these studies focused on Oryza sativa varieties whereas; O. glaberimma and its relatives are commonly grown in many upland rice grown areas of the tropical subSaharan Africa were not considered Matsumoto et al. (2014). Equally, NERICA is a newly bred rice lines with few information on their agronomic characteristics in many parts of Africa countries.
There has been several information on yield response of upland NERICA cultigens to moisture stress in many countries of sub-Sahara regions (Akinbile et al. 2007, Matsumoto et al. 2014, ikuta et al. 2017). Despite the significance of NERICA varieties in advancing rainfed upland rice production in sub-Sahara African nations, information is inconsistent on its water use efficiency including moisture deficit tolerance level by NERICA varieties. Furthermore, reports on how drought affects viability of rice seed when water deficit occur during anthesis are still incoherent. This study investigated effect of limited moisture supply during grain filling period on growth, and germinability of Oryza species.
MATERIALS AND METHODS
Description of the experimental site
This research was carried out at the screen house of Department of Crop Protection and Environmental Biology, University of Ibadan, Nigeria (Long. 7˚33΄ N and Lat. 3˚56 ՛ E and 213 m asl.). The relative humidity ranged from 80% to 84% while the annual rainfall ranged from 1250 mm to 1500 mm and minimum and maximum temperatures of the screen house during grain filling were 23.2° C and 33.5° C , respectively.
Source of plant materials, sowing and trial management
Four rice cultivars comprising CG14, NERICA2, NERICA4 and FARO 66 were obtained from National Centre for Genetic Resources and Biotechnology, (NACGRAB), Ibadan, Nigeria for this study. CG14 is African upland rice ( O glaberrima ), NERICA2 and NERICA4 are both derived from interspecific crosses between Oryza sativa x O. glaberrima progenies (Matsumoto et al. 2014) while FARO 66 is a lowland flood tolerant variety used as control.
The soil used for the study was collected from a field previously cultivated with rice in the last two growing seasons of 2013-2014 at the University Teaching and Research Farm. Prior to application of soil amendment, soil samples were dried, crushed and sieved with a 2 mm mesh sieve ande analyzed for pH as described by Cieśliński et al (1998); total nitrogen using jeldahl procedure followed by distillation described by Hseu
(2004); organic carbon using the method described by (Walkley 1947); available phosphorus and exchangeable bases such as Na+, +, Mg2+, Ca2+ and trace element (mg/kg): Zn2+, Cu2+, Mn2+ and Fe2+ by atomic absorption spectrophotometer. Also, soil particle analysis (mg/kg) was determined with the methods described by Bationo et al (2011). Five liters plastic pots were filled each with 5 kg soil. Seeds were sown through direct drilling into a nursery bed for two weeks after which they were transplanted into the pots at one plant per pot. The soil in the pot was amended with cattle dung at the rate of 8 t/ha. Weeds were rouged on sight and no other pest management was applied because we did not observe ant pest infestation
Treatments, experimental design and layout
The experiment had sixteen treatments including, four rice cultivars (CG14, NERICA2, NERICA4 and FARO 66) and four periods of water stress induction [0 (100% FC), 5, 10 and 15 days] at grain filling stage (47101 Days after transplanting, DAP). The study was laid out in completely randomized design and replicated six times.
Determination of soil moisture at field capacity
Soil moisture at field capacity of was determined using the methods of Gemeda and Datt (2018).
The water holing capacity of the soil was then evaluated as:
WH wss wss
WDS (g) = MC + FP + DS - MC + FP………..….. [W 0 ] WSS (g) = MC + FP + SS - MC + FP…………….[W 1 ] WWt = W 1 – W 0 …….………………………..…….. [W 2 ]
Where: WHC = Water holding capacity, WWt = Weight of water contained in saturated soil, WSS = Weight of the saturated soil, WDS = Weight of dry soil, MC =Mass of cup, FP = filter paper, DS = dry soil.
Imposition of drought stress
Maximum and minimum daily temperatures during the study particularly at grain filling were measured using thermometer. Days to flowering were monitored and each plant was labeled with its flowering date, defined as the day on which pollen had been released in approximately two thirds of florets composing the panicle. Prior to grain filling when the water stress treatment commenced, the pots were watered to field capacity using 250 ml of water every other day until five days after pollination (47 - 101 Days After Transplanting [DAT]), after which the treatments were applied [intermittent drought and no-drought (control)]. The water stress treatment comprised plants subjected to successive drought by withholding irrigation for 5, 10 and 15 days. At the end of each drought cycle i.e when the soil water potential reached -0.06 MPa, plants were re-watered once to field capacity (250 ml/5 kg pot) before another cycle of drought treatment. Drought treatment continued until seeds were matured for harvesting. Control plants were watered adequately as required to field capacity until the water drained freely through the holes at the base of the pots. Therefore, the four cultivars were subjected to drought for 30 days, based on days to maturity of each cultivar. Moisture stressed started at 51 (NERICA 2 and 4), 67 (FARO 66) and 71 (CG14) DAT. Three plants from each treatment were harvested on different dates within the developmental period of the grain. Sampling was done at the end of a drought cycle, before re-watering in the case of stressed plants.
Data collection
Data collection commenced at 7 Weeks after Transplanting (WAT) during which the plants have attained anthesis. Data collected per plant were based on growth parameters such as number of leaves, plant height, leaf area, number of tillers and yield parameters such as number of spikelets, weight of grains (filled and unfilled), percentage sterility, 100 seeds weight.
Dried tip and leaf rolling
Leaf rolling and dried tip were assessed on a scoring basis described by Swapna and Shylaraj (2017). The scores range from 1-9 as shown in Table 1.
Table 1. Scoring system for drought resistance for rice crop
5 Tube like 100% of all leaves dried, plant 9 rolling (no (no unrolling) generally unrolling) approach permanent wilting point, some die.
Source: International Rice Research Institute, (IRRI 1999).
Storability and viability of water stressed upland rice seeds
Harvested seeds were dried to 12% moisture content and then stored under ambient (27.5˚C) and at 4oC for 14 days after which germination test was carried out on the stored seeds using Petri dish method. Seeds collected at different stages of development were placed on Whatman No.1. filter paper in Petri dishes containing 6 ml distilled water and incubated at 30˚C for 8 days. The following germination indices were determined:
. „ . л. Number of normal seedlings that germinated i. Germination % =---------------:—■— ----
Total number of seeds planted
, - . Number of normal seedlings that germinated
-
i. Germination % =-----------■ —■—■ ----
- Total number of seeds planted
x 100 (ISTA, 2008)
it Coefficient of Velocity of Germination (CVG) = ^- .x 100
it Coefficient of Velocity of Germination (CVG) = ^7 .x 100
(Scott et al. 1984)
Where: N is the number of seeds germinated each days and T is the number of days corresponding to N.
-
iii. Germination Index (G.l) = E (Nx)(DAP) iii.Germination Index (G.l) = X (Nx)(DAP) (Akande et al. 2012)
Where: N is the number of normal seedlings that emerge on day x after seeding and DAP is Days after planting
-
iv. Germination Rate Index (GRI) = — + — + — +■■■— v ' 1 2 3 x
-
iv. Germination Rate Index (GRI) = — + — + — +■••— 1 ' 1 2 3 x
(Olisa et al. 2010)
Where: G is germination percentage on each day after seed placement and x is the corresponding day of germination.
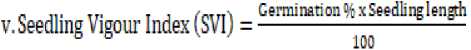
(Adebisi et al. 2008) 2.9. Data analysis
Data collected on germination test were transformed using logarithmic transformation. Analysis of variance (ANOVA) was then done using Statistical Analysis
System (Adebisi et al. 2008) (SAS). Means were separated using Duncan’s Multiple Range Test (DMRT) at P≤0.05. Relationship among some growth, yield and drought incidence was analyzed using linear regression.
RESULTS
Physico-chemical properties of the soil and cattle dung
The physical and chemical properties of the soil used for the experiment showed that the soil is slightly acidic with a pH of 6.2 and the textural class was loamy sand. The soil had low total nitrogen (14.2 mg/kg) while the available phosphorus (67.5 mg/kg) content of the soil was higher compared to that of the applied cattle dung (Table 2).
Influence of cultivar and imposed drought at anthesis on growth of upland rice
The effects of cultivars and water stress treatment on the height of upland rice plants are shown in Table 3. Results showed that the height of upland rice cultivars was not significantly (p≥0.05) influenced by water stress at the commencement of anthesis. Cultivar CG14 had tallest plants all through the sampling period. Cultivar CG14 water stressed at 10 days interval had significantly tallest plants (124.60 cm) compared to NERICA2 water stressed at 10 days interval and NERICA4 water stressed at 15 days interval both at 13 and 15 WAS.
Water deficit had significant influence on number of leaves produced at 13 and 15 WAT (Table 3). Cultivar FARO 66 had significantly higher number of leaves (15.75) at field capacity relative to NERICA4 watered at 10 days interval at 15 WAT.
Water stress cultivar had no significant effect on number of tillers formed by different rice cultivars. Cultivar FARO-66 under adequate irrigation recorded the highest tillers which was not significantly different from other treatments. The same trend was also observed all thought the sampling period.
The effect of water stress on leaf area of upland rice cultivars at grain filling is indicated in Table 3. The results showed that leaf area of upland rice was significantly affected by water stress. Cultivar CG14 under drought at 10 days interval recorded the highest leaf area (77.13 cm2) which was significantly different from those stressed at 15 days interval. Also, NERICA4 stressed at 15 days interval recorded the least leaf area (29.00 cm2).
Response of upland rice leaves to imposed drought at anthesis stage
Leaf rolling, an indicator of response of plant to drought was significant in cultivars subjected to longer period of moisture withdrawal. The highest leaf rolling score (0.95) was recorded in FARO-66 subjected to drought for 15 days compared to CG-14 cultivar which had least leaf rolling score (0.08) under adequate irrigation (Figure 1).
Moisture deficit significantly influenced the number of dried tips of the rice cultivars treated with different irrigation schedule or regime. Cultivars FARO-66 and NERICA4 had least dried tips (0.00) under field capacity condition (Figure 1). Although, the highest number of dried tips (0.92) was recorded in both CG-14 and NERICA2 subjected to drought cycle of 15 days interval but this was not statistically different from other cultivars under similar drought treatment (Figure 1).
Yield and yield components of upland rice cultivars as influenced by variety and water stress at anthesis
Yield and yield component response of the rice cultivars to water deficit at anthesis is indicated in Table 4. The results showed that occurrence of drought at anthesis had significant effect on yield and yield components of upland rice cultivars. There was statistical difference in the response of the different rice cultivars subjected to different duration of water stress at grain filling. The results revealed that FARO-66 adequately irrigated produced significantly higher number of spikelet (147.17) compared to NERICA2 predisposed to drought for 15 days at grain filling which had the least (44.98) spikelets.
Sterility of grains from different rice cultivars was significantly influenced by water deficit condition. The results revealed that NERICA2 subjected to drought at 15 days interval had higher percentage sterility (72.4%) relative to NERICA4 that received adequate irrigation which produced significantly least percent sterility (16.9%) under sufficient irrigation.
The total grain weight produced per plant was significantly influenced by drought. Different cultivars responded differently to varying duration of irrigation schedule. Generally, upland rice cultivars that were adequately irrigated produced highest grain yield while those predisposed to varying duration of irrigation regimes at grain filling had lower grain yield. Highest total grain yield (5.13 g) was produced by NERICA4 cultivar in adequately watered pots which was statistically comparable with FARO-66 treated with similar irrigation schedule. On the other hands least total grain (0.78 g) was obtained in NERICA2 line water stressed for 15 days this was not however significantly different from total grain yield produced by all the cultivars at this duration of water stress (Table 4).
Weight of filled grains of different upland rice cultivars was significantly affected by moisture stress at grain filling stage. NERICA4 had significantly highest filled grains weight (4.16 g) in pot supplied adequate water compared with other treatments. Conversely, NERICA2 subjected to drought at 15 days interval had least filled grains weight (0.29 g). FARO-66 had significantly highest weight of unfilled grain (0.95 g) in adequately watered pot. Least weight of unfilled grains (0.02 g) was recorded in NERICA2 variety that was adequately watered. Although, this weight was not significantly different from the same cultivar subjected to drought for 5 days interval treatment but was significantly different from the other treatments (Table 4). Similarly, cultivar CG14 and NERICA4 had statistically comparable weight of 100 seeds (2.83 and 2.79 g) under adequately irrigated situation which were higher than weight of 100 seeds (0.86 g) of NERICA2 cultivar under 15 days interval drought (Table 4).
Germinability of seeds of water stressed upland rice cultivars stored at two temperatures
The effect of storage temperatures and water stress at grain filling had significant (p≤0.05) effect on germination percentage of different rice cultivars (Table 5). At the third day after sowing, seeds of cultivar CG14 subjected to 15 days drought interval and stored at ambient temperature had significantly highest germination percentage (67.1%) while NERICA2 predisposed to drought for five days had the least
(2.5%) germination percentage. On the other hands, at four days after sowing, there was no significant difference in the germination percentage of seeds harvested from FARO-66 and CG14 cultivars predisposed to interval of 10 and 15 days drought and stored at 4oC or ambient temperatures, respectively. Least percentage germination (16.6%) was observed in seed obtained from NERICA2 subjected to 5 days drought and stored at 4oC (Table 6). Germination percentage of seeds of NERICA4 plants that was adequately irrigated and stored at ambient temperature had significantly highest germination percentage (83.4%) at five days after sowing whereas, seeds form NERICA2 plants water stressed for 10 days and stored at 4° C had the least germination percentage (25.8%). At the sixth day after sowing, seeds of FARO 66 water stressed for 15 days interval and stored at ambient temperature had significantly highest (83.4%) germination percentage while seeds from NERICA2 plant exposed to water deficit for 15 days and stored under ambient temperature had the least germination percentage. At the seventh day after sowing, seeds from different upland rice cultivars responded differently to varying period of water stress and stored either at 4° C or ambient temperature. However, seeds of FARO 66 had highest seed germination percentage (97.5%) across all the well watered, water stressed for 10 and 15 days and stored at 4° C or ambient temperature both at seventh and eight days after sowing. Least (39.1 and 36.2%) percentage germination were recorded in NERICA2, exposed to drought for 5 days and adequately watered and stored at ambient temperature and 4° C , respectively both at seventh and eighth day after sowing.
Effect of storage temperatures on germination indices of rice varieties predisposed to different drought cycles
The effect of storage temperatures, cultivars and water stress at grain filling on rice had significant (p≤0.05) effect on seed germinability. Germination index of seeds of different rice cultivars subjected to drought and stored at different temperature differed statistically (Table 6). At the end of the germination study, seeds harvested from cultivar CG 14 subjected to drought on 15 days interval and stored at ambient temperature had significantly highest germination index (35682) but this was not significantly different from FARO-66 and NERICA4 at field capacity under ambient temperature or FARO-66 water stressed for 15 days. Seeds from
NERICA2, subjected to drought for 15 days and stored at ambient temperature had significantly least (7406) germination index.
Table 2: Physical and chemical properties of soil and cattle dung used for the trial
|
Samples |
Soil Clay |
particle size (mg/kg) Silt Sand |
WHC (%) |
pH |
Avail. P (mg/kg) |
Organic Carbon (mg/kg) |
Total N (mg/kg) |
Exchangeable bases |
Trace element (mg/kg) |
||||||
|
Ca |
(Cmol/kg) |
K |
|||||||||||||
|
Mg |
Na |
Mn |
Fe |
Cu |
Zn |
||||||||||
|
Soil |
200 |
1340 8460 |
19.2 |
6.2 |
67.5 |
132.5 |
14.2 |
5.3 |
1.7 |
0.2 |
0.2 |
20.7 |
96.3 |
0.7 |
3.4 |
|
CDM |
na |
na na |
na |
6.4 |
16.7 |
243.1 |
196 |
39.5 |
12.4 |
16.7 |
71 |
287.5 |
68.6 |
11.3 |
25.5 |
CDM = Cattle dung, WHC = Water holding capacity, na = applicable
Table 3: Growth performance of different upland rice cultivars under imposed drought at anthesis
|
Cultivars |
Drought interval (days) |
Plant height (cm) |
Number of leaves |
Number of tillers |
Leaf area (cm2) |
||||||
|
7* |
13 |
15 |
7 |
13 |
15 |
7 13 15 |
7 |
13 |
15 |
||
|
FC |
92.87a |
104.10ab c |
106.15a b |
7.00a |
7.00ab |
7.00ab |
1.83a 2.17a 2.17a |
66.10ab |
74.35abc |
60.14abc |
|
|
5 |
88.17a |
104.97ab |
97.73ab c |
8.17a |
9.20ab |
9.75ab |
2.33a 2.33a 2.17a |
57.80abc |
74.98abc |
59.16abc |
|
|
CG14 |
10 |
97.28a |
122.01a |
124.60a |
10.33a |
11.33a b |
10.50a b |
2.67a 3.50a 3.17a |
76.77a |
88.11a |
77.13a |
|
15 |
75.06a b |
104.40ab c |
107.40a b |
5.73a |
7.20ab |
7.00ab |
1.11a 2.00a 2.00a |
46.38bcde |
83.93ab |
73.10ab |
|
|
FC |
52.02a b |
60.20bcd |
59.93bc |
5.08a |
6.33ab |
6.00ab |
1.29a 1.33a 1.33a |
29.77fed |
43.67def |
37.37cde |
|
|
5 |
81.95a b |
81.56abc d |
83.58ab c |
9.53a |
11.40a b |
10.80a b |
2.6a 2.80a 2.60a |
51.21bcd |
56.73cdef |
47.30cde |
|
|
NERICA 2 |
10 |
52.25a b |
52.63d |
53.42c |
6.33a |
6.00ab |
5.50ab |
1.83a 1.67a 1.17a |
26.71fe |
35.57ef |
30.29de |
|
15 |
71.64a b |
69.47abc d |
72.3abc |
7.55a |
7.25ab |
7.00ab |
1.91a 2.00a 2.00a |
44.35bcde f |
62.96abcd |
55.00abc |
|
|
FC |
87.03a b |
85.76abc d |
86.00ab c |
7.89a |
9.00ab |
8.40ab |
2.39a 2.80a 2.60a |
47.73bced 62.03abcde |
53.42bdc |
||
|
5 |
61.38a b |
65.6bcd |
66.18bc |
6.60a |
6.80ab |
6.60ab |
1.59a 1.80a 1.80a |
38.50cdef |
45.31def |
38.93cde |
|
|
NERICA 4 |
10 |
69.76a b |
78.06abc d |
79.36ab c |
4.93a |
5.40ab |
5.00b |
1.51a 1.81a 1.80a |
36.75cdef |
49.55cdef |
43.24cde |
|
15 |
45.62b |
57.52cd |
58.58bc |
4.73a |
5.20ab |
5.20ab |
0.91a 1.20a 1.20a |
22.87f |
32.91f |
29.00e |
|
|
FC |
72.56a b |
94.53abc d |
98.33ab c |
12.33a |
15.75a |
13.75a |
3.71a 4.25a 4.00a |
52.25bcd |
68.14abcd |
56.34abc |
|
|
5 |
67.31a b |
72.22abc d |
75.37ab c |
9.83a |
8.67ab |
8.00ab |
3.00a 2.83a 2.83a |
35.03cdef |
44.48def |
38.38cde |
|
|
FARO 66 |
10 |
75.23a b |
85.52abc d |
90.32ab c |
12.17a |
11.17a b |
9.67ab |
3.5a 3.17a 3.00a |
49.95bcd |
46.90def |
39.29cde |
|
15 |
77.72a b |
82.26abc d |
84.48ab c |
10.82a |
11.20a b |
10.40a b |
3.52a 3.80a 3.60a |
53.67bc |
58.79bcdef |
50.27bcd e |
|
Values are means of six plants, Means with the same letter on the same column are not significantly different at P > 0.05 using DMR., FC = Field capacity.* = Weeks after transplanting.
Table 4: Yield and yield components of upland rice as influenced by cultivar and water stress
|
Cultivars |
Drought interval (days) |
Number of spikelet |
Paddy sterility (%) |
Weight (g)/plant |
|||
|
Total grains |
Filled grain |
Unfilled grains |
100 seeds |
||||
|
FC |
124.17ab |
18.56de |
4.63ab |
3.56ab |
0.30ab |
2.83a |
|
|
CG-14 |
5 |
120.17abc |
24.64cde |
3.67abcd |
2.44bcd |
0.76ab |
2.58abc |
|
10 |
93.33bcde |
37.40bcde |
2.45def |
1.81cdef |
0.43ab |
2.23abcde |
|
|
15 |
77.17cde |
58.69ab |
1.17f |
0.83ef |
0.27ab |
1.30de |
|
|
FC |
94.88abcde |
17.51de |
2.50cdef |
2.29bcde |
0.02b |
2.35abcde |
|
|
5 |
87.46bcde |
25.54cde |
1.79ef |
1.50cdef |
0.05b |
1.95abcde |
|
|
NERICA-2 |
10 |
90.17bcde |
42.56bcd |
1.85ef |
1.43def |
0.38ab |
1.92abcde |
|
15 |
44.98e |
72.44a |
0.78f |
0.29f |
0.2ab |
0.86e |
|
|
FC |
134.67ab |
16.92e |
5.13a |
4.16a |
0.38ab |
2.79a |
|
|
5 |
130.91ab |
24.54de |
4.51abc |
3.43ab |
0.35ab |
2.37abcd |
|
|
NERICA-4 |
10 |
97.77abcde |
35.79bcde |
1.99ef |
1.48cdef |
0.44ab |
1.57bcde |
|
15 |
72.17de |
47.57abcd |
1.40f |
1.04ef |
0.37ab |
1.37de |
|
|
FC |
147.17a |
31.89cde |
4.98a |
3.88ab |
0.95a |
2.70ab |
|
|
FARO-66 |
5 |
109.50abcd |
24.18de |
3.23bcde |
2.53bc |
0.27ab |
2.64ab |
|
10 |
94.33bcde |
47.03abcd |
1.79ef |
1.28def |
0.65ab |
1.39de |
|
|
15 |
88.79bcde |
51.16abc |
1.63ef |
0.95ef |
0.83ab |
1.67cde |
|
|
Means with the same letter on the same column are not significantly different using DMRT at P<0.05, FC = |
Field capacity |
||||||
Table 5: Effect of storage temperatures on germination percentage of rice cultivars predisposed to different drought regime
|
Drought interval (days) |
Storage temperature (oC) |
Cultivars |
Germination percentage (Days after sowing) |
|||||
|
3 |
4 |
5 |
6 |
7 |
8 |
|||
|
CG14 |
36.2bcde |
42.1bcde |
47.9bcdefg |
50.8bcde |
53.8cde |
64.2bcd |
||
|
4 |
NERICA2 |
33.2bcde |
39.2bcde |
45.0bcdefg |
50.9bcde |
41.9de |
36.2e |
|
|
NERICA4 |
45.0bc |
47.9abcd |
53.8bcdef |
64.2abcd |
90.5ab |
84.5abc |
||
|
Field capacity |
FARO 66 |
45.0bc |
53.8ab |
70.1ab |
71.6abc |
97.5a |
97.5a |
|
|
CG14 |
39.2bcd |
47.9abcd |
53.8bcdef |
56.8abcde |
70.1abcd |
84.5abc |
||
|
Ambient |
NERICA2 |
29.9cdef |
29.9cdef |
36.2efg |
39.1ed |
42.1de |
45.0de |
|
|
(27.5) |
NERICA4 |
45.0bc |
45.0bcde |
83.4a |
71.6abc |
97.5a |
97.5a |
|
|
FARO 66 |
45.0bc |
45.0bcde |
67.1abc |
77.2ab |
84.5ab |
97.5a |
||
|
CG14 |
39.2bcd |
45.0bcde |
53.8bcdef |
56.8abcde |
84.5ab |
84.5abc |
||
|
4 |
NERICA2 |
2.5h |
16.6f |
28.8fg |
38.7ed |
41.9de |
47.9de |
|
|
NERICA4 |
41.9bc |
45.0bcde |
50.90cdefg |
50.8bcde |
77.5abc |
97.5a |
||
|
5 days |
FARO 66 |
39.1bcd |
45.0bcde |
56.8abcde |
64.2abcd |
77.2abc |
84.5abc |
|
|
CG14 |
47.9b |
45.0bcde |
50.8bcdefg |
56.8abcde |
64.2bcde |
84.5abc |
||
|
Ambient |
NERICA2 |
22.5efg |
25.8fe |
29.9efg |
35.8ed |
39.1e |
47.9de |
|
|
(27.5) |
NERICA4 |
45.0bc |
42.1bcde |
50.8bcdefg |
53.8abcde |
70.1abcd |
84.5abc |
|
|
FARO 66 |
36.2bcde |
45.0bcde |
67.1abc |
77.2ab |
90.5ab |
97.5a |
||
|
CG14 |
18.4gf |
29.9cdef |
45.0bcdefg |
47.9bcde |
64.2bcde |
64.2bcd |
||
|
4 |
NERICA2 |
22.5efg |
25.8fe |
25.8g |
38.7ed |
41.9de |
41.9de |
|
|
NERICA4 |
45.0bc |
50.8abc |
70.1ab |
77.5ab |
97.5a |
97.5a |
||
|
10 days |
FARO 66 |
48.0b |
67.1a |
67.1abc |
77.5ab |
97.5a |
97.5a |
|
|
CG14 |
39.2bcd |
42.1bcde |
45.0bcdefg |
50.8bcde |
67.1bcde |
67.1bcd |
||
|
Ambient |
NERICA2 |
10.5gh |
32.9bcdef |
39.1defg |
45.0ced |
53.8cde |
67.1bcd |
|
|
(27.5) |
NERICA4 |
26.6efd |
36.2bcdef |
42.1cdefg |
48.0bcde |
53.8cde |
61.2cde |
|
|
FARO 66 |
45.0bc |
50.8abc |
47.9bcdefg |
77.1ab |
90.5ab |
84.5abc |
||
|
CG14 |
42.1bc |
45.0bcde |
47.9bcdefg |
50.8bcde |
83.4ab |
97.5a |
||
|
4 |
NERICA2 |
33.2bcde |
36.2bcdef |
39.2defg |
45.0cde |
47.9de |
47.9de |
|
|
NERICA4 |
36. 2bcde |
39.2bcde |
42.1cdefg |
47.9bcde |
53.8cde |
77.5abc |
||
|
15 days |
FARO 66 |
47.9b |
50.9abc |
64.2abcd |
74.1abc |
84.5ab |
97.0a |
|
|
CG14 |
67.1a |
67.1a |
70.1ab |
77.5ab |
97.5a |
97.5a |
||
|
Ambient |
NERICA2 |
22.5efg |
26.6def |
29.9efg |
29.9e |
42.1de |
42.1de |
|
|
(27.5) |
NERICA4 |
39.2bcd |
42.1bcde |
42.1cdefg |
53.8abcde |
90.5ab |
90.45ab |
|
|
FARO 66 |
42.1bc |
47.9abcd |
47.9bcdefg |
83.4a |
97.5a |
97.5a |
||
Means with the same letter on the same column are not significantly different using DMRT at P<0.05.
Table 6: Effect of storage temperatures on germination indices of upland rice cultivars
|
Table 7: Regression analysis of influence components of rice cultivars |
of drought treatment at grain filling on |
growth and yield |
|
Predictors |
ᵦ |
t |
|
Leaf roll score |
0.457 |
6.815 |
|
Dry tip score |
0.332 |
4.449 |
|
Number of spikelet |
0.015* |
0.159 |
|
Percentage sterility |
0.087 |
1.199 |
|
Grain weight (g) |
0.049 |
0.269 |
|
Weight of filled grains |
-0.189 |
-1.071 |
|
Weight of unfilled grains |
-0.038* |
-0.687 |
|
Weight of 100 seeds |
-0.036* |
-0.499 |
|
Plant height at physiological maturity |
-0.027* |
-0.355 |
|
Number of tillers |
0.015* |
0.317 |
|
Leaf area at physiological maturity |
0.032* |
0.454 |
|
F |
82.22 |
|
|
R2 |
0.869 |
|
|
Adjusted R |
0.852 |
*significant at p<0.05 level
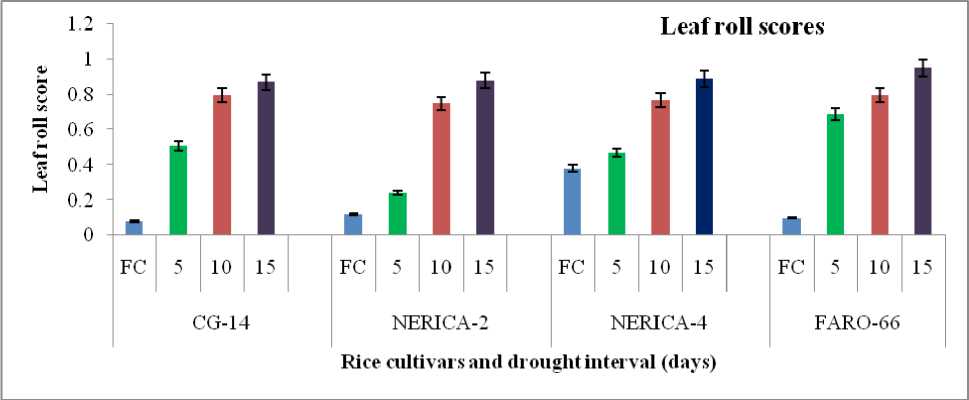

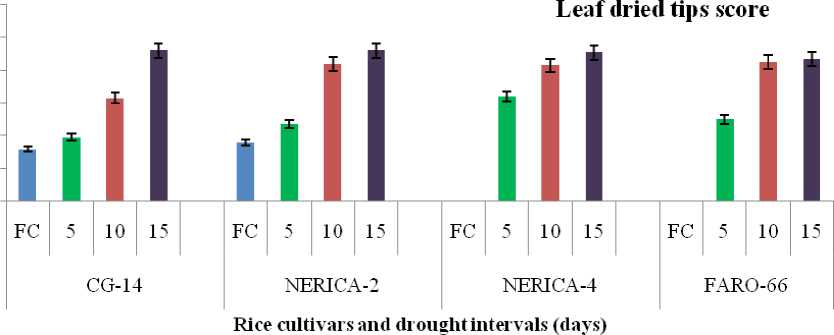
Figure 1: Effect of water stress on leaf morphology of four rice cultivars at grain filling.
Means of three plants with standard error bars at P<0.05, FC = Field capacity.
The results also showed that coefficient of velocity of germination was highest (0.37) in seeds from cultivar NERICA4 under field capacity and stored at ambient temperature while the least (0.15) coefficient of velocity of germination was recorded in cultivar FARO-66 subjected to drought for 5 days and stored at ambient temperature. Seeds of CG14 obtained from adequately watered plants and stored at 4oC had the highest germination rate index (3.26 %/day) which was significantly different from all other treatments while cultivar NERICA 2 stressed for 15 days and stored at 4ᴼC had the least germination rate index (0.71 %/day). Seedling vigor index was significantly highest (0.50) in seeds of NERICA4 cultivar from that was subjected to drought at of intervals 10 days and stored at 4oC while seeds from NERICA2 that was adequately irrigated and stored at 4oC had significantly the least (0.10) seedling vigor index.
Yield and yield components are significant (R2 = 0 87) predictors of severity of drought. Number of spikelet, weight of unfilled grains and 100 seeds are strongly related to the severity of drought on rice cultivars (Table 7).
DISCUSSION
All the four rice cultivars responded differently to different regimes of irrigation at the grain filling stage. The crop performed poorly with respect to growth, yield and seed germinability under severe drought condition irrespective of the cultivar. This was observed in the leaf morphological responses of these cultivars to irrigation scheduled at 15 days interval. This explains the importance of water in crop growth at the reproductive stage. It has been shown that at anthesis, many moisture mediated metabolic processes occur at this stage. Withdrawing moisture or occurrence of moisture deficit in form of drought at this stage may affect grain yield and its components negatively. Sikuku et al. (2010) have provided related explanation on the effects of water deficit on days to maturity and yield of three NERICA rainfed rice varieties. Cultivar CG14 showed better performance with respect to height and leaf area. This is per chance linked to the inherent tallness trait of its progenitor, Oryza glaberrima variety and ability to cope with many tropical biotic constraints which other rice varieties cannot tolerate. This is in accordance with conclusion of Megersa et al. (2016) that O. glaberrima thrives better in many severe environments difficult for O. sativa to survival. This is perhaps due to evolution and adaptation mechanisms that are tolerance of abiotic stresses especially drought.
Likewise, leaf area of the CG14 cultivar was observed to follow similar trend as the height. This is probably due to the fact that significant tallness of the cultivar predisposed it to intercept rays of sunlight, thus resulting in synthesis of more photoassimilate better than the other cultivars. This observation is in accordance with earlier reports by Ranawake et al. (2013) and Dada et al. (2014). The variation in number of leaves, number of tillers formed and leaf area is an indication that different genotypes had different environmental requirements for growth and development.
The significant improvement recorded in the number of tillers and leaves per plant of FARO-66 under optimum water supply can be ascribed to its genetic makeup as it relates to its hydrophilic nature. Ability of the cultivar to utilize maximally the available nutrients as well as partitioning of its photosynthate for tiller production more than the other cultivars could also explained the cultivar’s intolerant response to moisture stress. Similar thought have been reported by Akinwale et al. (2012) and im et al. (2015).
Water stress at grain filling drastically affected the shoots developed by all the rice cultivars negatively. The decline in growth may be connected to retardation in morphogenesis caused by moisture deficit at anthesis. Development of normal generative organs in cereal crops is directly related to proficient vegetative development which promotes adequate photosynthate accumulation and efficient assimilate partitioning into grain. This has profound influence on the amount and quality of the grain formed. Many researchers have given similar explanation on cereal crops. (Huijser and Schmid 2011, Hossain et al. 2012, Sié et al. 2012). The growth retardation observed in all the cultivars after imposition of water stress could be attributed to inhibition of cell elongation process due to interruption of water flow from the xylem to the surrounding elongating cells. Growth is accomplished through cell division, enlargement and differentiation as well as genetic, physiological, ecological and morphological events and their complex interactions. This agrees with previous reports by Hossain (2001) and Sangtarash (2010) that drought stress impairs cell division, elongation and expansion causing dwarf plant, reduced leaf area, reduced tiller formation and crop growth. Our observation disagrees with that of Liwani et al. (2019) where they reported that limiting water condition had no significant influence on growth parameters of wheat. Number of spikelet varied with cultivars and duration of drought intervals. Variation due to cultivars was primarily dependent on the number of tiller which was in turn affected by drought stress. The variety and water stress interaction effects on rice grain weight was significant at the various duration of drought cycle with the yield decreasing with increasing duration of the drought cycle. This suggests that formation of yield and its components is directly linked to moisture availability in sufficient quantity to be able to drive metabolic process in the plant. The variation in genetic makeup, amount of nutrient uptake and efficient utilization potential and photosynthates/dry matter partitioning may explain the observed differences in the cultivars responses. All the cultivars produced higher grain weight per plant when the plants were adequately watered. The deleterious effect of moisture stress on generative organs of rice cultivars followed the order: adequate moisture<5≤10<15 days drought intervals during grain filling depending on the cultivar. Reduced grain mass under drought especially prolonged water deficit might be related to inhibition of photosynthesis and impairment of assimilates translocation towards grain filling due to soil moisture stress. Protracted soil water deficit during active grain filling hinders or alters smooth partitioning of assimilates into sink which often lead to abortion and formation of poorly developed, sterile and unfilled grains. Clearly, this infers that drought has multidimensional effects on crop performance with devastating effect on cellular, molecular, biochemical and physiological attributes of crops Our observation is in line with that of Hossain et al. (2012) and Ambrosone et al. (2012), Salehi-Lisar and Bakhshayeshan-Agdam (2016).
The influence of drought and rice cultivars on the quantity of filled and unfilled grains per panicle was obviously severe. The reduced filled grains per panicle under insufficient soil moisture levels might be related to inhibition of assimilate translocation to the grains due to moisture deficit. These results are in consonance with the report of Hossain et al. (2012) and Liwani et al. (2019). The linear response of rice to severity of moisture stress with respect to weight of unfilled grains amplified the damaging effect of drought in reducing the quality and potential yield of rice plant. The exponentially higher quantity of unfilled grains per panicle under severe soil moisture deficit might be linked to inactive pollen particle; poorly developed pollen tube or alteration in assimilates production and its distribution to sink (grains). Similar reports have been presented by Farooq et al. (2009) eyvan (2010), and Osakabe et al. (2014). When crops are predisposed to limited water availability especially at anthesis and post anthesis stages, seed germination and germination indices are adversely affected. It appeared that exposure of rice paddy to longer period of drought enhanced seed germination at slower rate at early germination stage when compared to shorter period of water deficit. The better response observed at longer water deficit period may be due to initiation of synthesis of abscisic acid by the drought situation possibly because water limiting conditions triggers ABA synthesis in grains (Barnabás et al. 2008, Hossain et al. 2012, Salehi-Lisar and Bakhshayeshan-Agdam 2016).
The higher and better percentage germination observed in FARO 66 and NERICA4 compared to other cultivars as well as longer seedling of CG14 could be ascribed to differences in the inherent genetic nature of these cultivars. The variation in percentage germination and seedling length is an indication that different genotype has different inherent characteristics and genetic makeup, which could have relationship with their water use efficiency and adaptability or tolerance of water deficit condition. This has been reported by (Sié et al. 2012) and im im et al. (2015). There is a possibility that and NERICA4 tolerate water stress better than the other cultivars. Hence there is the need for explanatory genotypic and biochemical investigation on the enzymatic reactions that alters ABA concentration within seed and embryo during dormancy-to-germination switch especially in crops predisposed to post anthesis drought (Gilbert 2003, ermode 2005, Salehi-Lisar and Bakhshayeshan-Agdam 2016).
The research has shown that, storage temperature had no significant influence on germination indices and seedling length of water stressed upland rice seeds dried to 12% moisture content and stored for 14 days. This contradicts the reports of Wang et al. (2015){Wang, 2015 #109} that during storage, seed deterioration is temperature dependent in rice. However, the observation in this trial is in line with the findings of Wang et al. (2018) that even with high seed moisture content, the deterioration of rice seeds can be delayed for a few weeks at 30oC. Seed viability is influenced by the complex interrelationship among seed moisture content, storage temperature and storage period. Wang et al. (2015) indicated that the effect of storage temperature on seed deterioration increased as temperature increases. However, such positive correlation was not observed in this study, this is probably because the 14 days storage period was too short to reach the threshold point of seed deterioration. Besides, the 12% moisture content of the seeds was adequate not to trigger series of physiological metabolism that could reduce or alter viability and germinability of the rice seeds.
Germination indices of different rice cultivars were adversely affected by and moisture stress. This is in agreement with Walters et al. (2005) who reported that germination characteristics are affected by cultivar and moisture stress. In this study, cultivar CG14 and NERICA 4 demonstrated superior germination indices regardless of duration of moisture stress. The high rate and uniformity of seed germination irrespective of the duration of moisture stress can be ascribed to the lowered ABA level during seed imbibitions, which allow embryos to germinate and develop into seedlings. Similar observation had been reported by Reyes and Chua (2007) and Feng et al. (2014).
CONCLUSION AND RECOMMENDATION
Moisture deficit at grain filling affected growth, quality and quantity of grains produced by rice cultivars. The storage temperature had no effect on germinability of rice seeds that had been affected by moisture stress in field. Germinability of rice seeds is directly related to severity of drought at reproductive phase and not related to the storage temperature. Cultivars CG14 and NERICA4 tolerated moisture stress up to 10 days interval during grain filling better than NERICA2 and FARO66. To attain the maximum production potential of rice in many rice growing regions of the world, attention must be focused on mitigating the harmful effects of drought on staples particularly with the incidence of climate change. Observation from this study revealed that different rice cultivars responded differently to different levels of moisture deficit at grain filling. The timing, duration and severity of drought determine to a great extent how plants respond to water deficit. It should be noted, however that the reduction in performance of rice plant based on drought is a complex issue encompassing a number of factors rather than any single factor and thus further research is needed to fully elucidate these factors and the possible interaction among them.Moisture deficit at grain filling affected growth, quality and quantity of grains produced by rice cultivars. The storage temperature had no effect on germinability of rice seeds that had been affected by moisture stress in field. Germinability of rice seeds is directly related to severity of drought at reproductive phase and not related to the storage temperature. Cultivars CG14 and NERICA4 tolerated moisture stress up to 10 days interval during grain filling better than NERICA2 and FARO66. To attain the maximum production potential of rice in many rice growing regions of the world, attention must be focused on mitigating the harmful effects of drought on staples particularly with the incidence of climate change. Observation from this study revealed that different rice cultivars responded differently to different levels of moisture deficit at grain filling. The timing, duration and severity of drought determine to a great extent how plants respond to water deficit. It should be noted, however that the reduction in performance of rice plant based on drought is a complex issue encompassing a number of factors rather than any single factor and thus further research is needed to fully elucidate these factors and the possible interaction among them.
ACKNOWLEDGEMENTS
The authors would like to acknowledge National Centre for Genetic Resources and Biotechnology, (NACGRAB), Ibadan, Nigeria for providing the seeds used for the trial. North West university, Mafikeng Campus is acknowledged for support given to the lead author during the postdoctoral fellowship.
COMPETING INTERESTS
The authors declare no potential conflict of interest.
References Water stress at anthesis and storage temperature affected growth and germinability of rice (Oryza spp.)
- Adebisi, M., Ola, J., Akintobi, D. and Daniel, I. (2008) 'Storage life of sesame (Sesamum indicum L.) seeds under humid tropical conditions', Seed Science and Technology, 36(2), 379-387
- Adejare, F. and Umebese, C. (2008) 'Water stress induces cultivar dependent changes in stomatal complex, yield and osmotic adjustments in Glycine max L', Int. J. Agric. Res, 3(4), 287-295
- Ahmad, S., Ahmad, R., Ashraf, M. Y., Ashraf, M. and Waraich, E. A. (2009) 'Sunflower (Helianthus annuus L.) response to drought stress at germination and seedling growth stages', Pak. J. Bot, 41(2), 647-654
- Akande, S., Olakojo, S., Ajayi, S., Owolade, O., Adetumbi, J., Adeniyan, O. and Ogunbodede, B. (2012) 'Planting date affects cowpea seed yield and quality at southern Guinea Savanna, Nigeria', Seed Technology, 51-60
- Akinbile, C., Sangodoyin, A., Akintayo, I., Nwilene, F. and Futakuchi, K. (2007) Growth and yield responses of upland rice (NERICA 2) under different water regimes in Ibadan, Nigeria, translated by Asian Association for Agricultural Engineering
- Akinwale, M., Akinyele, B., Odiyi, A., Nwilene, F., Gregorio, G. and Oyetunji, O. (2012) Phenotypic screening of Nigerian rainfed lowland mega rice varieties for submergence tolerance, translated by 4-6
- Ambrosone, A., Costa, A., Leone, A. and Grillo, S. (2012) 'Beyond transcription: RNA-binding proteins as emerging regulators of plant response to environmental constraints', Plant Science, 182, 12-18
- Barnabás, B., Jäger, K. and Fehér, A. (2008) 'The effect of drought and heat stress on reproductive processes in cereals', Plant, cell & environment, 31(1), 11-38
- Bationo, A., Kimetu, J., Vanlauwe, B., Bagayoko, M., Koala, S. and Mokwunye, A. (2011) 'Comparative analysis of the current and potential role of legumes in integrated soil fertility management in West and Central Africa' in Fighting Poverty in Sub-Saharan Africa: The Multiple Roles of Legumes in Integrated Soil Fertility Management, Springer, 117-150
- Be, T. O., Arnell, N., Benito, G., Cogley, J., Jiang, T., Mwakalila, S., Fischer, T., Gerten, D., Hock, R. and Kanae, S. (2014) 'Freshwater resources' in. Chauhan, B. S., Jabran, K. and Mahajan, G. (2017) Rice production worldwide, Springer
- Chen, K. and Arora, R. (2011) 'Dynamics of the antioxidant system during seed osmopriming, post-priming germination, and seedling establishment in Spinach (Spinacia oleracea)', Plant Science, 180(2), 212-220
- Cieśliński, G., Van Rees, K., Szmigielska, A., Krishnamurti, G. and Huang, P. (1998) 'Low-molecular-weight organic acids in rhizosphere soils of durum wheat and their effect on cadmium bioaccumulation', Plant and soil, 203(1), 109-117
- Dada, O., Togun, A., Adediran, J. and Nwilene, F. (2014) 'Effect of Compost on Agro-Botanical Components Responsible for Rice (Oryza sativa) Grain Yield in Southwestern Nigeria', Journal of Agriculture and Sustainability, 6(1)
- Dada, O. A., Lawal, O. E., Kutu, F. R. and Olaniyan, A. B. (2018) 'Osmotic stress mitigation in upland NERICA field using compost augmentation and silicon enrichment', Research on Crops, 19(2)
- Farooq, M., Wahid, A., Kobayashi, N., Fujita, D. and Basra, S. (2009) 'Plant drought stress: effects, mechanisms and management' in Sustainable agriculture, Springer, 153-188
- Feng, C. Z., Chen, Y., Wang, C., Kong, Y. H., Wu, W. H. and Chen, Y. F. (2014) 'Arabidopsis RAV 1 transcription factor, phosphorylated by S n RK 2 kinases, regulates the expression of ABI 3, ABI 4, and ABI 5 during seed germination and early seedling development', The Plant Journal, 80(4), 654-668
- Fuhrmann, I., He, Y., Lehndorff, E., Brüggemann, N., Amelung, W., Wassmann, R. and Siemens, J. (2018) 'Nitrogen fertilizer fate after introducing maize and upland-rice into continuous paddy rice cropping systems', Agriculture, Ecosystems & Environment, 258, 162-171
- Gemeda, W. S. and Datt, S. P. (2018) 'Characterization of soils under major land uses in Chencha District, South Ethiopia', Journal of Soil Science and Environmental Management, 9(8), 127-137
- Gilbert, S. F. (2003) 'The morphogenesis of evolutionary developmental biology', International Journal of Developmental Biology, 47(7-8), 467
- Hossain, A., da Silva, J. A. T., Lozovskaya, M. V. and Zvolinsky, V. P. (2012) 'High temperature combined with drought affect rainfed spring wheat and barley in South-Eastern Russia: I. Phenology and growth', Saudi journal of biological sciences, 19(4), 473-487
- Hossain, M. (2001) Growth and yield performance of some boro rice cultivars under different soil moisture regimes, unpublished thesis MS Thesis, Dept. Crop Bot. Bangladesh Agric. Univ., Mymensingh
- Hseu, Z.-Y. (2004) 'Evaluating heavy metal contents in nine composts using four digestion methods', Bioresource technology, 95(1), 53-59
- Huijser, P. and Schmid, M. (2011) 'The control of developmental phase transitions in plants', development, 138(19), 4117-4129
- Kermode, A. R. (2005) 'Role of abscisic acid in seed dormancy', Journal of Plant Growth Regulation, 24(4), 319-344
- Keyvan, S. (2010) 'The effects of drought stress on yield, relative water content, proline, soluble carbohydrates and chlorophyll of bread wheat cultivars', J. Anim. Plant Sci, 8(3), 1051-1060
- Kikuta, M., Makihara, D., Arita, N., Miyazaki, A. and Yamamoto, Y. (2017) 'Growth and yield responses of upland NERICAs to variable water management under field conditions', Plant Production Science, 20(1), 36-46
- Kim, J.-M., Sasaki, T., Ueda, M., Sako, K. and Seki, M. (2015) 'Chromatin changes in response to drought, salinity, heat, and cold stresses in plants', Frontiers in plant science, 6, 114
- Lilley, J. and Fukai, S. (1994) 'Effect of timing and severity of water deficit on four diverse rice cultivars III. Phenological development, crop growth and grain yield', Field Crops Research, 37(3), 225-234
- Liwani, U., Magwaza, L. S., Odindo, A. O. and Sithole, N. J. (2019) 'Growth, morphological and yield responses of irrigated wheat (Triticum aestivum L.) genotypes to water stress', Acta Agriculturae Scandinavica, Section B-Soil & Plant Science, 1-8
- Mancosu, N., Snyder, R., Kyriakakis, G. and Spano, D. (2015) 'Water scarcity and future challenges for food production', Water, 7(3), 975-992
- Masoud, S., Saoub, H., Migdadi, H. and Al-Nashash, A. (2005) 'Evaluation of Jordanian barley (Hordeum vulgare L.) landraces collected from diverse environments', Dirasat Agric. Sci, 32, 163-170
- Matsumoto, S., Tsuboi, T., Asea, G., Maruyama, A., Kikuchi, M. and Takagaki, M. (2014) 'Water response of upland rice varieties adopted in sub-Saharan Africa: A water application experiment', Rice Research: Open Access
- Megersa, A., Seo, J., Chin, J. H., Kim, B. and Koh, H.-J. (2016) 'Characterization of Selected Rice Varieties Adapted in Africa', Plant Breeding and Biotechnology, 4(3), 297-305
- Molden, D. (2013) Water for food water for life: a comprehensive assessment of water management in agriculture, Routledge
- Mut, Z., Akay, H. and Aydin, N. (2010) 'Effects of seed size and drought stress on germination and seedling growth of some oat genotypes (Avena sativa L.)', African Journal of Agricultural Research, 5(10), 1101-1107
- Olisa, B., Ajayi, S. and Akande, S. (2010) 'Physiological quality of seeds of promising African yam bean (Sphenostylis stenocarpa (Hochst. ex A. Rich) Harms) and pigeon pea (Cajanus cajan L. Mill sp.) landraces', Res. J. Seed Sci, 3(2), 93-101
- Osakabe, Y., Osakabe, K., Shinozaki, K. and Tran, L.-S. P. (2014) 'Response of plants to water stress', Frontiers in plant science, 5, 86
- Ranawake, A., Amarasinghe, U. and Senanayake, S. (2013) 'Determination of some agronomic characters in different age groups of traditional rice cultivars in Sri Lanka under fertilized and non-fertilized conditions', Journal of Agricultural Biotechnology and Sustainable Development, 5(5), 79-83
- Reyes, J. L. and Chua, N. H. (2007) 'ABA induction of miR159 controls transcript levels of two MYB factors during Arabidopsis seed germination', The Plant Journal, 49(4), 592-606
- Salehi-Lisar, S. Y. and Bakhshayeshan-Agdam, H. (2016) 'Drought stress in plants: causes, consequences, and tolerance' in Drought Stress Tolerance in plants, Vol 1, Springer, 1-16
- Samarah, N. and Alqudah, A. (2011) 'Effects of late-terminal drought stress on seed germination and vigor of barley (Hordeum vulgare L.)', Archives of Agronomy and Soil Science, 57(1), 27-32
- Sangtarash, M. (2010) 'Responses of different wheat genotypes to drought stress applied at different growth stages', Pakistan Journal of Biological Sciences, 13(3), 114
- Scott, S., Jones, R. and Williams, W. (1984) 'Review of Data Analysis Methods for Seed Germination 1', Crop science, 24(6), 1192-1199
- Sié, M., Sanni, K., Futakuchi, K., Manneh, B., Mandé, S., Vodouhe, R., Dogbe, S., Drame, K., Ogunbayo, A. and Ndjiondjop, M.-N. (2012) 'Towards a rational use of African rice (Oryza glaberrima Steud.) for breeding in Sub-Saharan Africa'
- Sikuku, P., Netondo, G., Musyimi, D. and Onyango, J. (2010) 'Effects of water deficit on days to maturity and yield of three NERICA rainfed rice varieties', ARPN Journal of Agricultural and Biological Science, 5(3), 1-9
- Steduto, P., Hsiao, T. C., Fereres, E. and Raes, D. (2012) Crop yield response to water, fao Rome
- Su, Z., Ma, X., Guo, H., Sukiran, N. L., Guo, B., Assmann, S. M. and Ma, H. (2013) 'Flower development under drought stress: morphological and transcriptomic analyses reveal acute responses and long-term acclimation in Arabidopsis', The Plant Cell, 25(10), 3785-3807
- Sun, Y. D., Du, X. H., Zhang, W. J., Sun, L. and Li, R. (2011) Seed germination and physiological characteristics of Amaranthus mangostanus L. under drought stress, translated by Trans Tech Publ, 1071-1074
- Swapna, S. and Shylaraj, K. S. (2017) 'Screening for osmotic stress responses in Rice varieties under drought condition', Rice Science, 24(5), 253-263
- Walkley, A. (1947) 'A critical examination of a rapid method for determining organic carbon in soils-effect of variations in digestion conditions and of inorganic soil constituents', Soil science, 63(4), 251-264
- Walters, C., Wheeler, L. M. and Grotenhuis, J. M. (2005) 'Longevity of seeds stored in a genebank: species characteristics', Seed Science Research, 15(1), 1-20
- Wang, W., He, A., Peng, S., Huang, J., Cui, K. and Nie, L. (2018) 'The effect of storage condition and duration on the deterioration of primed rice seeds', Frontiers in plant science, 9, 172
- Wang, Y., Yang, L., Chen, X., Ye, T., Zhong, B., Liu, R., Wu, Y. and Chan, Z. (2015) 'Major latex protein-like protein 43 (MLP43) functions as a positive regulator during abscisic acid responses and confers drought tolerance in Arabidopsis thaliana', Journal of experimental botany, 67(1), 421-434
- Yaklich, R. (1984) 'Moisture stress and soybean seed quality', Journal of Seed Technology, 60-67
- Ye, N., Jia, L. and Zhang, J. (2012) 'ABA signal in rice under stress conditions', rice, 5(1), 1
- Zhang, J., Zhang, S., Cheng, M., Jiang, H., Zhang, X., Peng, C., Lu, X., Zhang, M. and Jin, J. (2018) 'Effect of Drought on Agronomic Traits of Rice and Wheat: A Meta-Analysis', International journal of environmental research and public health, 15(5)
