Закономерность изменения электрического сопротивления миокарда как косвенное подтверждение явления поочередного сокращения кластеров кардиомиоцитов

Автор: Шевченко Ю.Л., Лычкова А.Э., Ульбашев Д.С.
Журнал: Вестник Национального медико-хирургического центра им. Н.И. Пирогова @vestnik-pirogov-center
Рубрика: Редакционные статьи
Статья в выпуске: 3 т.17, 2022 года.
Бесплатный доступ
Несмотря на современные знания о физиологии сердца, остаются до конца не изученными процессы регуляции сократительной функции миокарда в условиях покоя, физической нагрузки и стресса. Сложное воздействие симпатической и парасимпатической нервной системы на миокард, подключение различных многочисленных интра- и экстракардиальных регуляторных механизмов способствует возможности поочередного сокращения отдельных ассоциаций кардиомиоцитов, что обеспечивает энергетически более выгодную работу сердца. Изучение регуляции сердечной деятельности может быть связано с использованием методов, основанных на измерении импеданса ткани, которые сейчас широко используются в различных медико-биологических исследованиях. Цель: определить колебания изменений электрического сопротивления различных зон миокарда в состоянии функционального покоя и при стимуляции парасимпатического и симпатического отделов нервной системы. Материалы и методы. Кроликам породы шиншилла (n = 24) проводилась препаровка правого и левого блуждающих нервов, звездчатого ганглия. В миокард передней стенки левого желудочка субэпикардиально устанавливались электроды для регистрации импеданса миокарда. Инвазивно измерялось артериальное давление в сонной артерии на протяжении всего эксперимента. Повышенная нагрузка на миокард создавалась раздражением правого звездчатого ганглия импульсами тока, напряжением 3-7 мВ, 2 мс; правого блуждающего нерва - 1,5-5 мВ, 2 мс. Результаты. Выявлено статистически значимое различие трех кластеров миокарда с колебаниями сопротивления высокой амплитуды (8,0-11,0), средней (3,0-8,0) и низкой (менее 3,0), (р function show_abstract() { $('#abstract1').hide(); $('#abstract2').show(); $('#abstract_expand').hide(); }
Физиология сердца, кластеры миокарда, поочередность сокращения, симпатическая и парасимпатическая нервная система, кардиомиоциты
Короткий адрес: https://sciup.org/140296576
IDR: 140296576 | DOI: 10.25881/20728255_2022_17_3_11
The regularity of changes in the electrical resistance of the myocardium as an indirect confirmation of the phenomenon of alternate contraction of cardiomyocyte clusters
Despite modern knowledge about the physiology of the heart, the processes of regulation of the contractile function of the myocardium in conditions of rest, physical activity and stress remain completely unexplored. The complex effect of the sympathetic and parasympathetic nervous system on the myocardium, the connection of various numerous intra- and extracardial regulatory mechanisms contributes to the possibility of alternately reducing individual associations of cardiomyocytes, which provides energetically more beneficial heart function. The study of the regulation of cardiac activity may be associated with the use of methods based on the measurement of tissue impedance, which are now widely used in various biomedical research. Objective: to determine fluctuations in the electrical resistance of various zones of the myocardium in a state of functional rest and during stimulation of the parasympathetic and sympathetic parts of the nervous system. Materials and methods. Chinchilla rabbits (n = 24) underwent dissection of the right and left vagus nerves, stellate ganglion. Electrodes were installed subepicardially in the myocardium of the anterior wall of the left ventricle to register the myocardial impedance. The arterial pressure in the carotid artery was measured invasively throughout the experiment. The increased load on the myocardium was created by irritation of the right stellate ganglion with current pulses of 3-7 mV, 2 ms; of the right vagus nerve - 1.5-5 mV, 2 ms. Results. A statistically significant difference was revealed between three myocardial clusters with resistance fluctuations of high amplitude (8.0-11.0), medium (3.0-8.0) and low (less than 3.0), (p function show_eabstract() { $('#eabstract1').hide(); $('#eabstract2').show(); $('#eabstract_expand').hide(); }
Текст научной статьи Закономерность изменения электрического сопротивления миокарда как косвенное подтверждение явления поочередного сокращения кластеров кардиомиоцитов
Процессы регуляции сократительной функции сердца в условиях покоя, нагрузки, при раздражении симпатического (адренергического) и парасимпатического (холинергического) отделов вегетативной нервной системы исследовались на протяжении долгих лет, однако множество вопросов, связанных со сложностью взаимодействия их влияния остаются и в наше время [1].
Регуляция деятельности сердца в связи с изменяющимися потребностями организма осуществляется интракардиальными, экстракардиальными и гуморальными механизмами [2; 3]. Внутрисердечные нервные сплетения формируются за счет афферентных и эфферентных симпатических и парасимпатических элементов [4].
К экстракардиальным механизмам относят холинергические, адренергические, серотонинергические, пури-нергические и нитрергические нервные и гуморальные

Шевченко Ю.Л., Лычкова А.Э., Ульбашев Д.С.
ЗАКОНОМЕРНОСТЬ ИЗМЕНЕНИЯ ЭЛЕКТРИЧЕСКОГО СОПРОТИВЛЕНИЯ МИОКАРДА КАК КОСВЕННОЕ ПОДТВЕРЖДЕНИЕ ЯВЛЕНИЯ ПООЧЕРЕДНОГО СОКРАЩЕНИЯ КЛАСТЕРОВ КАРДИОМИОЦИТОВ факторы регуляции [5–10]. Известно, что блуждающий нерв содержит весьма значительное количество чувствительных афферентных нервных волокон, практически 80% [1]. Аксоны постганглионарных нейронов парасимпатического пути идут к узлам проводящей системы сердца, мышечным пучкам предсердий, в меньшей степени к желудочкам. Блуждающие нервы обеспечивают иннервацию кровеносных и лимфатических сосудов [11–13]. Правый блуждающий нерв преимущественно действует на синоатриальный узел, вызывая замедление или остановку сердечных сокращений, а левый — на атриовентрикулярный, определяя проведение возбуждения в другие отделы сердца [14].
Симпатическое влияние на сердце вызывает положительный ино-, хроно-, дромо- и батмотропный ответы. Правые и левые симпатические нервы не равнозначны по влиянию: правый в большей степени увеличивает частоту сердечных сокращений, а левый — изменяет длительность предсердно-желудочкового интервала [15]. Улучшается синхронизация сокращений отдельных миокардиальных волокон, увеличивается скорость проведения возбуждения в предсердиях, атриовентрикулярном узле и желудочках. При раздражении звездчатого узла происходит резкое увеличение скорости изменения давления в обоих желудочках во время систолы и диастолы. Таким образом, положительное инотропное действие симпатических нервов на сердце является не только результатом увеличения силы сокращения каждого волокна, но и следствием большей синхронизации их деятельности [16].
О взаимоотношениях симпатических и парасимпатических нервов при их влиянии на сократительную функцию сердца имеется несколько точек зрения: антагонизм, синергизм, и взаимокомпенсация. Периферические рефлексы, осуществляемые с помощью экстраорганных ганглиев, протекают при обязательном участии холинергических и адренергических структур [17]. Первые являются пусковыми, вторые — модулирующими [18]. Предполагается, что во взрослом организме при нахождении в покое, парасимпатические нервные влияния в сердце преобладают над симпатическими [19; 20]. Уменьшение содержания ацетилхолина или снижение функциональной активности парасимпатической нервной системы приводит к развитию патологии и может быть причиной возникновения внезапной смерти. Симпатическая нервная система в этой ситуации выступает как синергист парасимпатической — это предотвращает развитие острой декомпенсации сердечной деятельности [21].
Постганглионарные симпатические и парасимпатические нервные окончания в сердце часто располагаются близко друг к другу, что является субстратом для их сложных взаимодействий [22; 23]. Адреналин блокирует проведение возбуждения в ганглиях так же, как обнаруженный в них норадреналин. Его источником являются либо хромаффинные клетки (SIF-клетки), либо другие вставочные нейроны, а, возможно, и преганглионарные волокна. Кроме того, катехоламины могут выделяться под влиянием экзо- генного и эндогенного ацетилхолина, освобождающегося в самом ганглии во время его возбуждения [24].
По мнению некоторых исследователей, парасимпатическая иннервация желудочков выражена слабо и оказывает свое влияние косвенно за счет торможения симпатических эффектов: ингибируется высвобождение норадреналина из окончаний постганглионарных симпатических волокон под действием ацетилхолина. Происходят взаимодействия на уровне самих эффекторных клеток, обусловленные циклическими нуклеотидами (цАМФ и цГМФ) через интракардиальную нервную систему [25–27] С участием α-адренорецепторных образований нервных терминалей и миокардиоцитов вызывается гиперполяризация мембран нейронов симпатических ганглиев [28]. Определенную роль в механизме усиления вагусного торможения деятельности сердца при раздражении звездчатого ганглия играют и другие биологически активные вещества (серотонин, АТФ и ее производные) [29].
Изучение регуляции сердечной деятельности может быть связано с использованием методов, основанных на измерении импеданса ткани, которые сейчас широко используются в различных медико-биологических исследованиях. Изучение зависимости изменения электрических показателей миокарда, в том числе его сопротивления, от множества интра- и экстракардиальных воздействий, может помочь понять различные особенности сокращения сердечной мышцы.
В настоящее время нет однозначных ответов на вопросы о необходимости этого сложного взаимодействия симпатической и парасимпатической нервной систем в регуляции сердечной деятельности. Вероятнее всего, возможность такого разнонаправленного воздействия на миокард, подключение многочисленных интра- и экстракардиальных регуляторных механизмов при адаптации к физической нагрузке и стрессу способствует сложной форме сокращений ассоциаций кардиомиоцитов с целью сохранения энергии и продолжительности работы сердца.
Материалы и методы
Эксперименты in vivo на 24 кроликах обоих полов породы «шиншилла» массой 3100±550 г проведены в соответствии с ГОСТ 10993.6–2009 и с требованиями «Правил лабораторной практики в Российской Федерации» (Приказ Министерства здравоохранения и социального развития Российской Федерации №708 н от 23.08.2010). Все манипуляции с животными проведены согласно принципам гуманного отношения к животным в соответствии с Международными рекомендациями, а также с соблюдением биоэтических норм и требований Международного комитета по науке.
Интраоперационно в щадящих условиях с использованием искусственной вентиляции легких фиксированному на спине животному проводилась препаровка правого и левого блуждающих нервов, они перерезались на уровне щитовидного хряща гортани, препарировался звездчатый ганглий и правая сонная артерия. Выпол-
Шевченко Ю.Л., Лычкова А.Э., Ульбашев Д.С.
ЗАКОНОМЕРНОСТЬ ИЗМЕНЕНИЯ ЭЛЕКТРИЧЕСКОГО СОПРОТИВЛЕНИЯ МИОКАРДА КАК КОСВЕННОЕ ПОДТВЕРЖДЕНИЕ ЯВЛЕНИЯ ПООЧЕРЕДНОГО СОКРАЩЕНИЯ КЛАСТЕРОВ КАРДИОМИОЦИТОВ нялся доступ к сердцу во II–III межреберье. В миокард передней стенки левого желудочка с помощью круглой кишечной иглы диаметром 0,1 мм субэпикардиально вводились электроды для регистрации импеданса миокарда, прежде всего, в переднюю стенку левого желудочка. Разрезы в грудной клетке ушивались, электроды подключались к реографу. В сонную артерию вводилась канюля, заполненная гепаринизированным физиологическим раствором.
Для более полного анализа регистрировались быстрые и медленные колебания импеданса миокарда при изменении уровня сократительной активности на реографе РГ 4-01 с установленным переключателем типа ТГ-1-2 (Рис. 1).
Каждый электрод представлял собой провод ПЭЛ-12 на опорной площадке, освобожденный от изоляции 1,5–2 мм (отступя на 2 мм от места прикрепления). Для исключения искажения импедансограммы при трении опорных площадок друг о друга каждый электрод был фиксирован на расстоянии, превышающем диаметр опорной площадки (2–2,5 мм). Площадь активной поверхности таких электродов составила 0,5–0,7 мм2. Проводились расчеты в трех отведениях по три пары электродов, расположенных рядом на расстоянии 5 мм друг от друга (Рис. 2).
Для оценки изменения сопротивления пучков кардиомиоцитов на кривой импедансограммы измерялась максимальная амплитуда волны; параллельно во время эксперимента оценивалась частота сердечных сокращений, величина систолического, диастолического и пульсового давления. Повышенная нагрузка на миокард создавалась раздражением правого звездчатого ганглия импульсами тока напряжением 3–7 мВ, 2 мс; правого блуждающего нерва — 1,5–5 мВ, 2 мс.
Для статистического анализа использовалась программа Statistica 12. Показатели описательной статистики включали определение следующих величин: число наблюдений (n), среднее значение (М), стандартное отклонение (SD). Учитывая малую выборку, неправильное распределение, использовали непараметрические U-критерий Манна-Уитни и критерий Вилкоксона (для парных выборок). Различия считались статистически значимыми при p<0,05.
Результаты
Первым этапом проводилась непрерывная запись изменений сопротивления участков миокарда в покое. Оказалось, что с каждым сердечным циклом у кроликов происходили колебания импеданса миокарда в исследуемых зонах, демонстрирующие определенную чередующуюся последовательность изменения рефрактерного периода. Выявлено статистически значимое различие трех кластеров с колебаниями сопротивления высокой амплитуды (8,0–11,0 Ом), средней (3,0–8,0 Ом) и низкой (волна γ: менее 3,0 Ом), (статистически значимые различия согласно критерию Манна-Уитни при р<0,05).
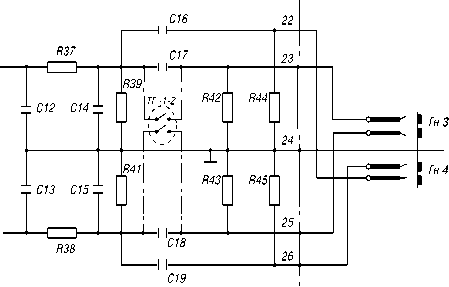
Рис. 1. Схема реографа для регистрации импеданса миокарда.
Рис. 2. Схема проведения эксперимента (подключение электродов к сердцу).
На втором этапе проводилась серия исследований с записью колебаний импеданса отдельных зон кардиомиоцитов.
В первом отведении за цикл сокращения сердца наибольший показатель сопротивления отмечался в участке миокарда между первой парой электродов. В соседней зоне, между второй парой электродов наблюдалось снижение показателей до средних значений, а в третьей зоне этого отведения выявлялся минимальный импеданс, то есть происходило чередование изменений импеданс от высоких показателей до низких.
Во втором отведении прослеживались аналогичные процессы, только в другой последовательности: промежуточный показатель сопротивления определялся между первой парой электродов, между второй парой — самые низкие значения, между третьей — максимальные.
В третьем отведении сохранялось последовательное изменение электрической активности участков
Шевченко Ю.Л., Лычкова А.Э., Ульбашев Д.С.
ЗАКОНОМЕРНОСТЬ ИЗМЕНЕНИЯ ЭЛЕКТРИЧЕСКОГО СОПРОТИВЛЕНИЯ МИОКАРДА КАК КОСВЕННОЕ ПОДТВЕРЖДЕНИЕ ЯВЛЕНИЯ ПООЧЕРЕДНОГО СОКРАЩЕНИЯ КЛАСТЕРОВ КАРДИОМИОЦИТОВ
Табл. 1. Импеданс участков миокарда левого желудочка (первое исследование)
|
Электроды |
Сопротивление, Ом |
|
Первое отведение |
|
|
1-я пара |
10,2±1,1 |
|
2-я пара |
5,7±0,7 |
|
3-я пара |
0,4±0.05 |
|
Второе отведение |
|
|
1-я пара |
6,3±0,8 |
|
2-я пара |
0,3±0,05 |
|
3-я пара |
9,8±0,7 |
|
Третье отведение |
|
|
1-я пара |
0,5±0,03 |
|
2-я пара |
10,5±1,2 |
|
3-я пара |
6,2±0,5 |
Табл. 2. Импеданс участков миокарда левого желудочка (второе исследование)
|
Электроды |
Сопротивление, Ом |
|
Первое отведение |
|
|
1-я пара |
0,4±0,01 |
|
2-я пара |
8,7±0,5 |
|
3-я пара |
10,2±0,3 |
|
Второе отведение |
|
|
1-я пара |
9,3±0,6 |
|
2-я пара |
1,0±0,05 |
|
3-я пара |
7,4±0,6 |
|
Третье отведение |
|
|
1-я пара |
7,2±0,6 |
|
2-я пара |
8,4±0,3 |
|
3-я пара |
0,5±0,02 |


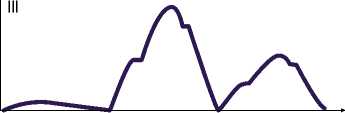
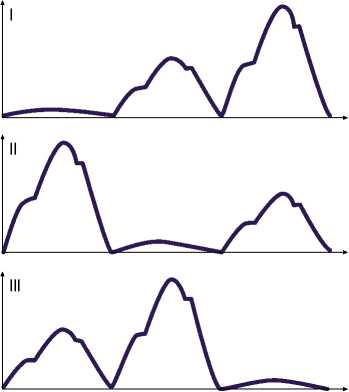
Рис. 4. Схематическое изображение колебаний сопротивления на трех электродах в отведениях (I–III) при втором исследовании.
Рис. 3. Схематическое изображение колебаний сопротивления на трех электродах в отведениях (I–III) при первом исследовании.
миокарда, регистрировался показатель импеданса в следующей последовательности: между первой парой электродов — наименьший, между второй — максимальный, между третьей — промежуточный (табл. 1) (Рис. 3).
Проводилось второе исследование. В первом отведении наименьшее сопротивление отмечалось между первой парой электродов, средние значения — в области второй пары, максимальные — между третьей. Во втором отведении, между второй парой электродов зарегистрировано минимальное значение сопротивления при сохранении импеданса миокарда на высоком уровне между первой и промежуточном — между третьей парой электродов. В третьем отведении минимальные значения сопротивления выявлено на третьем участке миокарда, а максимальные — на втором (табл. 2) (Рис. 4).
При третьем измерении импеданса миокарда кролика в покое чередование зон с высоким сопротивлением, средним и низким сохранялось (табл. 3) (Рис. 5).
Табл. 3. Импеданс участков миокарда левого желудочка (третье исследование)
|
Электроды |
Сопротивление, Ом |
|
Первое отведение |
|
|
1-я пара |
6,7±0,9 |
|
2-я пара |
0,6±0,05 |
|
3-я пара |
10,5±0,4 |
Второе отведение
|
1-я пара |
9,3±0,6 |
|
2-я пара |
5,7±0,5 |
|
3-я пара |
0,7±0,04 |
Третье отведение
|
1-я пара |
0,3±0,06 |
|
2-я пара |
10,6±0,3 |
|
3-я пара |
7,5±0,4 |
Третьим этапом проводилось создание специальных условий повышенной нагрузки. При стимуляции правого звездчатого ганглия наблюдалось значимое увеличение сопротивления на всех исследуемых участках миокарда (табл. 4).

Шевченко Ю.Л., Лычкова А.Э., Ульбашев Д.С.
ЗАКОНОМЕРНОСТЬ ИЗМЕНЕНИЯ ЭЛЕКТРИЧЕСКОГО СОПРОТИВЛЕНИЯ МИОКАРДА КАК КОСВЕННОЕ ПОДТВЕРЖДЕНИЕ ЯВЛЕНИЯ ПООЧЕРЕДНОГО СОКРАЩЕНИЯ КЛАСТЕРОВ КАРДИОМИОЦИТОВ
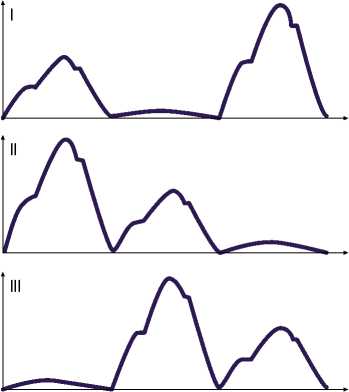
Табл. 4. Импеданс участков миокарда левого желудочка при стимуляции правого звездчатого ганглия
|
Электроды |
Сопротивление, Ом |
|
Первое отведение |
|
|
1-я пара |
10,7±0,8 |
|
2-я пара |
11,6±0,6 |
|
3-я пара |
10,2±0,3 |
|
Второе отведение |
|
|
1-я пара |
11,3±0,8 |
|
2-я пара |
11,8±0,6 |
|
3-я пара |
10,3±0,4 |
|
Третье отведение |
|
|
1-я пара |
10,4±0,6 |
|
2-я пара |
10,6±0,3 |
|
3-я пара |
10,7±0,6 |
Рис. 5. Схематическое изображение колебаний сопротивления на трех электродах в отведениях (I–III) при третьем исследовании.
При стимуляции блуждающего нерва наблюдался отрицательный хронотропный эффект, незначительно уменьшалось артериальное давление (табл. 5), однако несимметрично происходило увеличение амплитуды волн импедансограммы в I и II отведениях, меньшее по сравнению с предыдущим исследованием. Выявлено уменьшение сопротивления в III отведении (табл. 6).
Обсуждение
Изменение импеданса (сопротивления) на разных участках миокарда зависит от чередования рефрактерных периодов пучков кардиомиоцитов [30; 31]. Было проведено несколько этапов исследования, все колебания показателей импеданса миокарда разделены на три вида: с высокими значениями, средними и низкими. На основе проведенного эксперимента установлено, что в миокарде существуют зоны с последовательно чередующимися периодами невозбудимости, то есть происходит сменная работа ассоциаций кардиомиоцитов. Вероятно, есть различные режимы сокращения этих кластеров. С течением эксперимента происходила миграция гипо- и акинетич-ных зон между электродами, но всегда присутствовали кластеры с максимальной активностью, что говорит о возможности наличия именно трех чередующихся зон сокращения.
Во время нагрузки на миокард в виде стимуляции симпатической нервной системы, — в эксперименте отмечалось увеличение значения импеданса между регистрирующими электродами во всех трех кластерах. То есть терялся принцип поочередности сокращения. Вегетативной нервной системе присуще облегчение передачи возбуждения в нервно-мышечных синапсах за счет увеличения освобождения медиатора на каждый импульс при повторных редких раздражениях. Так, при раздражении нерва током низкой частоты (2–16 Гц) или при естественной импульсации — содержание норадре-
Табл. 5. Изменение артериального давления и ЧСС при стимуляции блуждающего нерва
|
Параметры |
В покое |
При стимуляции |
|
Частота сердечных сокращений (в 1 мин) |
150±11,3 |
134±10,3 |
|
Систолическое артериальное давление (мм рт.ст.) |
72±5,5 |
70,5±5,1 |
|
Диастолическое артериальное давление (мм рт.ст.) |
51±6,0 |
47±5,4 |
Табл. 6. Импеданс участков миокарда левого желудочка при стимуляции блуждающего нерва
Раздражение правого блуждающего нерва имитировало повышенную статическую нагрузку на миокард (аналогичную таковой у спортсменов-штангистов). Благодаря деятельности холинэстеразы, ацетилхолин, освобожденный из парасимпатических терминалей, имел ограниченный доступ к мускариновым рецепторам, связанным с калиевыми каналами. В сердце ацетилхолин взаимодействует с МЗ-холинорецепторами и активирует посредством гуаниннуклеотидзависимых

Шевченко Ю.Л., Лычкова А.Э., Ульбашев Д.С.
ЗАКОНОМЕРНОСТЬ ИЗМЕНЕНИЯ ЭЛЕКТРИЧЕСКОГО СОПРОТИВЛЕНИЯ МИОКАРДА КАК КОСВЕННОЕ ПОДТВЕРЖДЕНИЕ ЯВЛЕНИЯ ПООЧЕРЕДНОГО СОКРАЩЕНИЯ КЛАСТЕРОВ КАРДИОМИОЦИТОВ
G-белков быстрые управляемые калиевые каналы. Это приводит к увеличению проницаемости для ионов К+ и уменьшению проницаемости мембраны для Са2+ посредством угнетения системы аденилатциклазы — цАМФ. Усиленный ток К+ из клетки и ограничение входа Са2+ в клетку вызывало ее гиперполяризацию [1]. Следствием этого являются замедление деполяризации пейсмекерных клеток и снижение времени проведения возбуждения в атриовентрикулярном узле, что сопровождалось уреже-нием сокращений сердца.
В эксперименте при раздражении правого блуждающего нерва произошла утрата сменного режима работы пучков кардиомиоцитов. Выявилось парадоксальное увеличение сократимости зон миокарда, вероятно, это было связано с выделением малых доз ацетилхолина (10–20–22 ммоль/л) в этих участках, которые, как известно, оказывают необычное стимулирующее влияние на деятельность сердца, тогда как большие дозы медиатора (10–5–6 ммоль/л) вызывают обычный тормозной эффект. Медиатор парасимпатической системы влиял на интрамуральные нейроны миокарда, синаптически связанные с α- и β-адренорецепторами миокарда [7]. При стимуляции блуждающего нерва симметричное увеличение импеданса происходило не во всех отведениях. Определена зона с уменьшением сопротивления, вероятно, это связано с тем, что менялась нормальная регуляция сократительной деятельности пучков кардиомиоцитов, и некоторые близкорасположенные зоны функционально объединялись.
Разнородность рецепторных структур, на которые воздействовал ацетилхолин, косвенно демонстрирует возможность поочередной регуляции сокращения кластеров кардиомиоцитов. При этом блуждающий нерв, влияя на состояние вставочных дисков, изменял силу сокращений миокарда, включая в функцию различное количество мышечных пучков. Возможно, что функция вставочных дисков регулируется и внутрисердечной нервной системой, которая обеспечивает синхронизацию поочередного сокращения ассоциаций кардиомиоцитов [5]. Подобное длительное стрессовое воздействие может приводить к развитию «электрофизиологического станнинга», в дальнейшем к гибернации и декомпенсации сердечной деятельности [32; 33].
Заключение
В соседних зонах миокарда происходит чередование рефрактерных периодов, изменяется сопротивление от минимальных показателей до максимальных. На основе полученных экспериментальных данных можно предположить, что одномоментно в миокарде существует три группы кардиомиоцитов с различной сократительной активностью, последовательно сменяющие в сокращении друг друга, этот принцип поочередной работы функционирует в условиях нормы. При появлении продолжительного стрессового воздействия привычный режим сменяется «включением» большей зоны миокарда, проис- ходит объединение «рабочих» кластеров кардиомиоцитов, что в дальнейшем может приводить к электрическому истощению, «оглушению», гибернации и развитию декомпенсации сердечной деятельности.
Авторы заявляют об отсутствии конфликта интересов (The authors declare no conflict of interest).
ЗАКОНОМЕРНОСТЬ ИЗМЕНЕНИЯ ЭЛЕКТРИЧЕСКОГО СОПРОТИВЛЕНИЯ МИОКАРДА КАК КОСВЕННОЕ ПОДТВЕРЖДЕНИЕ ЯВЛЕНИЯ ПООЧЕРЕДНОГО СОКРАЩЕНИЯ КЛАСТЕРОВ КАРДИОМИОЦИТОВ
Список литературы Закономерность изменения электрического сопротивления миокарда как косвенное подтверждение явления поочередного сокращения кластеров кардиомиоцитов
- Смирнов В.М. Физиология человека. — М.: Медицина, 2002. [Smirnov VM. Fiziologiya cheloveka. M.: Medicina, 2002. (In Russ).]
- Косицкий Г.И., Червова И.А. Сердце как саморегулирующаяся система. (Интрамуральная нервная система и ее роль в регуляции функции сердца). — М.: Медицина, 1968. [Kosickij GI, Chervova IA. Serdce kak samoreguliruyushchayasya sistema. (Intramural'naya nervnaya sistema i ee rol' v regulyacii funkcii serdca). — M.: Medicina, 1968. (In Russ).]
- Зефиров Т.Л., Зиятдинова Н.И., Хисамиева Л.И., Зефиров А.Л. Сравнительный анализ влияния блокады al- и а2- адренорецепто-ров на сердечную деятельность крыс в постнатальном онтогенезе // Бюллетень экспериментальной биологии и медицины. — 2011. — Т.151. — №6. — С.607-610. [Zefirov TL, Ziyatdinova NI, Hisamieva LI, Zefirov AL. Sravnitel'nyj analiz vliyaniya blokady al- i a2- adrenorec-eptorov na serdechnuyu deyatel'nost' krys v postnatal'nom ontogeneze. Byulleten' eksperimental'noj biologii i mediciny. 2011; 151(6): 607-610. (In Russ).]
- Швалев В.Н., Сосунов А.А., Гуски Г. Морфологические основы иннервации сердца. — М.: Наука, 1992. [Shvalev VN, Sosunov AA, Guski G. Morfologicheskie osnovy innervacii serdca. M.: Nauka, 1992. (In Russ).]
- Косицкий Г.И. Афферентные системы сердца. — М.: Наука, 1975. [Kosickij GI. Afferentnye sistemy serdca. M.: Nauka, 1975. (In Russ)].
- Лычкова А.Э. Влияние серотонинергической и холинергической нервных систем и их медиаторов на деятельность сердца и моторную активность гладкомышечных органов малого таза // Бюлл. экспер. биол. и мед. — 2004. — T.138. — №10. — C.368-372. [Lychkova AE. Vliyanie serotoninergicheskoj i holinergicheskoj nervnyh sistem i ih mediatorov na deyatel'nost' serdca i motornuyu aktivnost' gladkomyshechnyh organov malogo taza. Byull. eksper. boil. i med. 2004; 138(10): 368-372. (In Russ).]
- Лычкова А.Э. Серотонинергическая нервная система: градиенты нервных влияний в норме и патологии // Экспер. клин. гастроэнтерол. — 2003. — Т.6. — C.114-120. [Lychkova AE. Serotoninergicheskaya ner-vnaya sistema: gradienty nervnyh vliyanij v norme i patologii. Eksper. klin. gastroenterol. 2003; 6: 114-120. (In Russ).]
- Лычкова А.Э., Смирнов В.М. Механизмы синергизма симпатической и парасимпатической нервной системы в регуляции деятельности сердца и желудка // Вестник РАМН. — 2002. — №4. — C.16-20. [Lychkova AE, Smirnov VM. Mekhanizmy sinergizma simpaticheskoj i parasimpatich-eskoj nervnoj sistemy v regulyacii deyatel'nosti serdca i zheludka. Vestnik RAMN. 2002; (4): 16-20. (In Russ).]
- Шевченко Ю.Л., Бобров Л.Л., Обрезан А.Г. Диастолическая функция левого желудочка. — М.: ГЭОТАР-Медиа, 2002. [Shevchenko YuL, Bo-brov LL, Obrezan AG. Diastolicheskaya funkciya levogo zheludochka. M.: GEOTAR-Media, 2002. (In Russ).]
- Шевченко Ю.Л. Медико-биологические и физиологические основы клеточных технологий в сердечно-сосудистой хирургии. — Санкт-Петербург: Наука, 2006. [Shevchenko YuL. Mediko-biologicheskie i fiziolo-gicheskie osnovy kletochnyh tekhnologij v serdechno-sosudistoj hirurgii. Sankt-Peterburg: Nauka, 2006. (In Russ).]
- Klein HU, Ferrari GM. Vagus nerve stimulation: A new approach to reduce heart failure. Cardiol J. 2010; 17(6): 638-44.
- Zandstra TE, Notenboom RGE, Wink J, et al. Asymmetry and Heterogeneity: Part and Parcel in Cardiac Autonomic Innervation and Function. Front Physiol. 2021; 12: 665298. doi: 10.3389/fphys.2021.665298.
- Page PL, Dandal N, Cardinal R, Nadeau R. Comparison of the infution of acetylcholine into the artry of the sinoatrial node with the elactric stimulation of cardiac parasympathetic nerves. Ann. Chir. 1995; 49(8): 719-727.
- Yamakawa K, So EL, Rajendran PS, et al. Electrophysiological effects of right and left vagal nerve stimulation on the ventricular myocardium. Am J Physiol Heart Circ Physiol. 2014; 307(5): H722-31. doi: 10.1152/ajpheart.00279.2014.
- Coote JH, Chauhan RA. The sympathetic innervation of the heart: Important new insights. Auton Neurosci. 2016; 199: 17-23. doi: 10.1016/ j.autneu.2016.08.014.
- Motiejunaite J, Amar L, Vidal-Petiot E. Adrenergic receptors and cardiovascular effects of catecholamines. Ann Endocrinol (Paris). 2021; 82(3-4): 193-197. doi: 10.1016/j.ando.2020.03.012.
- Goldstein RS, Kalcheim C. Normal segmentation and size of the primary sympathetic ganglia depend upon the alternation of rostrocaudal properties of the somites. Development. 1991; 112(1): 327-34. doi: 10.1242/ dev. 112.1.327.
- Smith PG, Warn JD, Steinle JJ, et al. Modulation of parasympathetic neuron phenotype and function by sympathetic innervation. Auton Neurosci. 2002; 96(1): 33-42. doi: 10.1016/s1566-0702(01)00371-x.
- Ng GA, Brack KE, Coote JH. Effects of direct sympathetic and vagus nerve stimulation on the physiology of the whole heart--a novel model of isolated Langendorff perfused rabbit heart with intact dual autonomic innervation. Exp Physiol. 2001; 86(3): 319-29. doi: 10.1113/eph8602146.
- Masuda Y. Role of the parasympathetic nervous system and interaction with the sympathetic nervous system in the early phase of hypertension. J Cardiovasc Pharmacol. 2000; 36 Suppl 2: S61-4. doi: 10.1097/ 00005344-200000006-00013.
- Oberhauser V, Schwertfeger E, Rutz T, et al. Acetylcholine release in human heart atrium: influence of muscarinic autoreceptors, diabetes, and age. Circulation. 2001; 103(12): 1638-43. doi: 10.1161/01.cir.103.12.1638.
- Khan AA, Lip GYH, Shantsila A. Heart rate variability in atrial fibrillation: The balance between sympathetic and parasympathetic nervous system. Eur J Clin Invest. 2019; 49(11): e13174. doi: 10.1111/eci.13174.
- Smith-White MA, Wallace D, Potter EK. Sympathetic-parasympathetic interactions at the heart in the anaesthetised rat. J Auton Nerv Syst. 1999; 75(2-3): 171-5. doi: 10.1016/s0165-1838(98)00169-6.
- Kawai Y, Senba E. Correlation between dendrodendritic synapses of adrenergic type and synaptically evoked hyperpolarization in the sympathetic ganglion of adult rats. Neuroscience. 1995; 68(3): 925-35. doi: 10.10-16/0306-4522(95)00202-t.
- Benarroch EE. Physiology and Pathophysiology of the Autonomic Nervous System. Continuum (Minneap Minn). 2020; 26(1): 12-24. doi: 10.1212/ CON.0000000000000817.
- Schwartz PJ, De Ferrari GM. Sympathetic-parasympathetic interaction in health and disease: abnormalities and relevance in heart failure. Heart Fail Rev. 2011; 16(2): 101-7. doi: 10.1007/s10741-010-9179-1.
- Камардина О.Л., Басс И.Н. Анатомия и физиология: Учеб. пособие для иностр. учащихся. Ч. 1. Ордена Ленина гос. ун-т им. Ленинского комсомола. Подгот. фак. для иностр. граждан. — Воронеж, 1971. [Kamardina OL, Bass IN. Anatomiya i fiziologiya: [Ucheb. posobie dlya inostr. uchashchihsya] CH. 1. Ordena Lenina gos. un-t im. Leninskogo komsomola. Podgot. fak. dlya inostr. grazhdan. Voronezh, 1971. (In Russ.)]
- Neto FR, Sperelakis N. Analysis of the hyperpolarizing effect of catecho-lamines on canine cardiac Purkinje fibres. Br J Pharmacol. 1989; 96(3): 591-8. doi: 10.1111/j.1476-5381.1989.tb11857.x.
- Hatip-Al-Khatib I, Bolukba^i-Hatip F. Modulation of the negative inotropic effect of haloperidol by drugs with positive inotropic effects in isolated rabbit heart. Pharmacology. 2002 Sep;66(1):19-25. doi: 10.1159/000063249.
- Беляков К.Ф. Изменение ширины приишемической и отдаленной зон миокарда // Кардиология. — 1980. — Т.4. — С.96-98 [Belyakov KF. Iz-menenie shiriny priishemicheskoj i otdalennoj zon miokarda. Kardiologiya. 1980; 4: 96-98 (In Russ).]
- Беляков К.Ф. Сократительная функция отдельных участком миокарда и сердца в целом при острой коронарной недостаточности: Дис. ... канд. мед. наук. — Москва; 1973 [Belyakov KF. Sokratitel'naya funkciya otdel'nyh uchastkom miokarda i serdca v celom pri ostroj koronarnoj ned-ostatochnosti: Dis. ... kand. med. nauk. Moskva; 1973. (In Russ).]
- Burgdorf C, Richardt D, Kurz T, et al. Adenosine inhibits norepinephrine release in the postischemic rat heart: the mechanism of neuronal stunning. Cardiovasc Res. 2001; 49(4): 713-20.
- Canty JMJr, Fallavollita JA. Chronic hibernation and chronic stunning: a continuum. J Nucl Cardiol. 2000; 7(5): 509-27.
