Закономерности динамики еловых насаждений в старолесьях Подмосковья

Автор: Абатуров Александр Васильевич, Браславская Татьяна Юрьевна, Королева Светлана Юрьевна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Лесные ресурсы
Статья в выпуске: 1-3 т.16, 2014 года.
Бесплатный доступ
В старовозрастных еловых насаждениях Подмосковья уже более 30 лет проводятся наблюдения динамики лесообразующих видов на постоянных пробных площадях. Выявлено, что насаждения обычно представлены одним поколением ели, и динамика его размерной структуры соответствует теоретическим представлениям о результатах конкурентных отношений в древостое. Однако из одновозрастных насаждений не формируются разновозрастные, поскольку возобновление идет неудовлетворительно. Поэтому в насаждении неизбежна смена лесообразователя после естественного распада господствующего в древостое поколения ели.
Ель европейская, размерная структура, возрастная структура, ценопопуляция, возобновление
Короткий адрес: https://sciup.org/148202851
IDR: 148202851 | УДК: 581.524.34
Regularities of fir-tree plantings dynamics in old-age spruce stands of Moscow region
In old-age fir-tree plantings of Moscow region more than 30 years observations of forest forming types dynamics on constant trial squares are made. It is revealed that plantings are usually presented by one generation of a fir-tree, and dynamics of its dimensional structure corresponds to theoretical ideas of results of the competitive relations in a forest stand. However of even-aged plantings aren't formed uneven-age as renewal goes unsatisfactorily. Therefore in planting change of forest forming after natural disintegration of generation of a fir-tree dominating in a forest stand is inevitable.
Текст научной статьи Закономерности динамики еловых насаждений в старолесьях Подмосковья
одна и та же нумерация кварталов, что облегчает возможность использовать архивные сведения по истории лесопользования [3]. К моменту начала исследований здесь было больше спелых и перестойных насаждений с господством или участием ели, поэтому им было уделено больше внимания.
Для проведения исследований подбирали участки размером не менее 50х50 м со спелыми и перестойными деревьями и малым участием сухостойных и суховершинных деревьев. В выбранных участках закладывали постоянные пробные площади (ППП) для многолетних исследований. Программа, содержание и первые результаты этих исследований ранее публиковались [1, 4, 12]. В ходе их непременным условием было сохранение и восстановление нумерации деревьев. В значительной мере благодаря этому были получены новые данные о лесоводствен-ных свойствах основных лесообразующих пород. Сегодня, через 30-40 лет после начала этой работы, исследуемые насаждения распадаются, быстро или медленнее, под влиянием возраста, ветровала и бурелома, вредителей и болезней. Предметом настоящего обсуждения является динамика состояния еловых насаждений на 4-х ППП, материалы по которым дополнены результатами обследований 2009-2011 гг. (проведенных с участием аспиранта Института лесоведения РАН А.П. Кулешова). Лесотипологические и таксационные характеристики обсуждаемых насаждений в момент первого обследования (при 100-летнем возрасте деревьев) приведены в табл. 1. Там же даны сведения о местонахождении ППП в ближнем Подмосковье, что позволяет оценить степень воздействия на них атмосферного загрязнения со стороны Москвы. Все обсуждаемые насаждения возникли и формировались под влиянием разнообразной хозяйственной деятельности человека, что известно из лесохозяйственных документов [4].
Таблица 1. Характеристики пробных площадей при первом обследовании
|
№ ППП* |
Площадь Состав пород Местонахождение Тип леса Год обследования |
Порода |
Возраст, лет |
Число стволов, шт. на 1 га |
Средние |
Запас древесины, м3 на 1 га |
|||||
|
живые |
сухие |
всего |
высота, м |
диаметр, см |
живые |
сухие |
всего |
||||
|
1 |
0,3 га 10Е Лосино-Погонный лесопарк, кв.19, клетка 3 (3,5 км к С от Москвы) ельник кислично-зеленчуковый 1975 |
Е |
90÷120 |
298 |
298 |
28 |
38 |
446 |
446 |
||
|
Лп |
10 |
10 |
27 |
22 |
|||||||
|
Ос |
3 |
3 |
18 |
4 |
4 |
||||||
|
Всего |
311 |
3 |
314 |
444 |
444 |
||||||
|
2 |
0,25 га 10Е+Лп, ед.Д ЛосиноПогонный лесопарк, кв.19, клетка 5 (2,5 км к С от Москвы) ельник с липой зеленчу-ково-волосистоосоковый 1975 |
Е |
100÷120 |
368 |
12 |
380 |
27 |
29 |
343 |
6 |
349 |
|
Лп |
100÷120 |
608 |
4 |
612 |
20 |
15 |
47 |
47 |
|||
|
Лп |
**252 |
**252 |
17 |
||||||||
|
Д |
32 |
32 |
4 |
||||||||
|
Кл.о. |
28 |
28 |
4 |
||||||||
|
Ос |
8 |
8 |
18 |
||||||||
|
Всего |
1044 |
16 |
1060 |
390 |
|||||||
|
3 |
0,25 га 10Е Лосино-Погонный лесопарк, кв.19, клетка 4 (3,5 км к С от Москвы) ельник кислично-зеленчуковый 1975 |
Е |
90÷100 |
296 |
8 |
304 |
26 |
36 |
410 |
410 |
|
|
Лп |
128 |
128 |
20 |
14 |
10 |
10 |
|||||
|
Лп |
**13 |
**13 |
18 |
||||||||
|
Д |
52 |
8 |
60 |
21 |
|||||||
|
Д |
**8 |
**8 |
**16 |
24 |
|||||||
|
Кл.о. |
192 |
4 |
196 |
8 |
|||||||
|
Всего |
668 |
20 |
688 |
420 |
420 |
||||||
|
4 |
2,25 га 8Е 2Д +Ос+Б, ед. С,Лп,Кл.о. Красногорский лесопарк, кв.16 (10 км к СЗ от Москвы) ельник с дубом зеленчу-ково-кислично-волосистоосоковый 1980 |
Е |
50+90 +110÷150 |
559 |
6 |
565 |
24 |
25 |
340 |
340 |
|
|
Д |
90÷110 |
105 |
46 |
151 |
18 |
26 |
60 |
30 |
90 |
||
|
Всего |
664 |
52 |
716 |
400 |
30 |
430 |
|||||
Примечания: * В публикации [4] принята иная нумерация ППП (с. 65-67, табл. 4): № 1 не имеет специального номера (п. 7 в табл. 4), № 2 указана как № 4, № 3 – как № 5, № 4 – как № 6. ** Число деревьев диаметром более 11 см
На рис. 1 для каждого из 4-х еловых насаждений показано (в виде сглаженных огибающих кривых), как изменилось за время наблюдений распределение живых деревьев ели по ступеням диаметра ствола (2 см). Анализ распределения размерного признака (в данном случае – диаметра ствола на высоте 1,3 м) позволяет выделять совокупности деревьев, внутренне статистически однородные и обособленные друг от друга (если их несколько). Такие совокупности предлагалось называть элементами леса [14]. В элементе леса распределение числа деревьев по какому-либо размерному признаку, равно как и по календарному возрасту, имеет мономодаль-ный характер [8, 14]. Достаточно четкие рамки календарного возраста (узкие относительно общей продолжительности жизни дерева) позволяют называть совокупность деревьев одного и того же вида, относящихся к одному элементу леса, популяционно-демографическим термином «когорта» [6, 7].
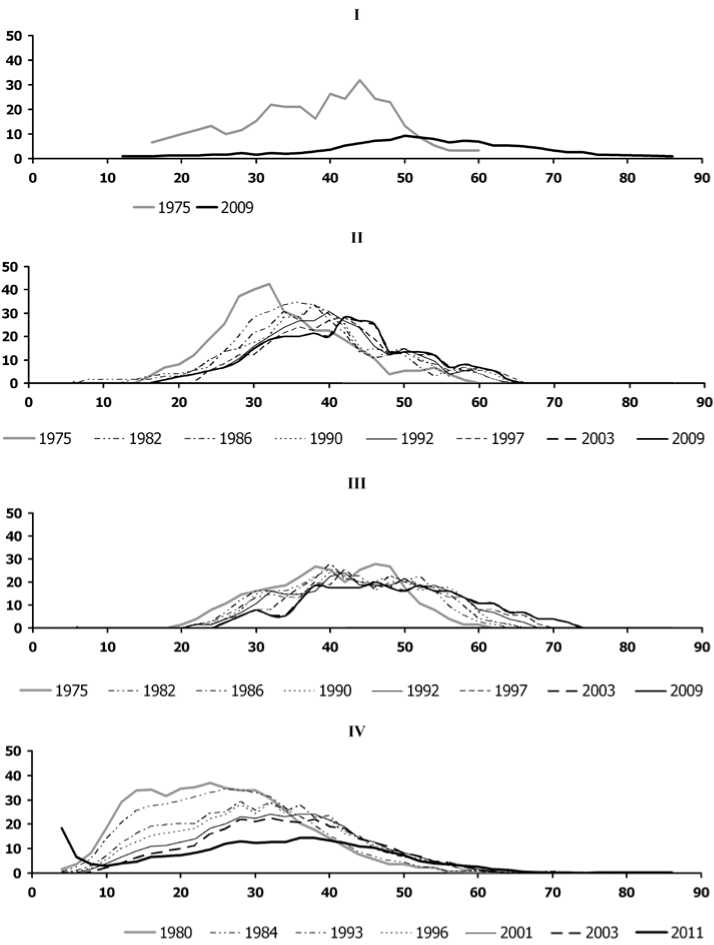
Рис. 1. Распределение деревьев ели по диаметру ствола в разные годы наблюдений:
I – ППП № 1, II – ППП № 2, III – ППП № 3, IV – ППП № 4. По оси абсцисс – ступени диаметра ствола (2см) на высоте 1,3м, по оси ординат – число деревьев (шт./га). Огибающие кривые для разных годов показаны линиями разного типа
С.А. Дыренков [8] показал, что в ранний период жизни элемента леса распределение различных размерных признаков деревьев характеризуется сильной левой асимметрией, но впоследствии становится все более и более симметричным, приближаясь к нормальному. Достижение нормального типа распределения можно рассматривать как показатель наибольшего развития древостоя, ослабления в нем конкуренции и проявления всей возможной для него наследственной нормы реакции на условия произрастания. Кроме увеличения симметричности с течением времени, динамика распределения деревьев по размеру проявляется в виде сдвига вправо (в сторону больших значений) как общего диапазона, так и положения максимума, а также в виде расширения диапазона и уменьшения амплитуды максимума. Такие закономерности С.А. Дыренков и другие исследователи, не имея возможности провести многолетние прямые наблюдения, обычно выводили косвенно – на основе сравнения параметров у древостоев разного возраста, произрастающих в разных участках единого лесного массива.
На каждой из рассматриваемых нами 4-х ППП деревья ели в момент первого обследования можно отнести к одному (и единственному) элементу леса, поскольку их распределение в диапазоне диаметра ствола было непрерывным и почти везде мономодальным. Выявив это, можно заключить, что в каждом насаждении формирование ценопопуляции (ЦП) ели включало всего лишь один этап – посадку или же естественное заселение елью участка, освободившегося после сельскохозяйственного использования или пройденного пожаром.
Возраст деревьев ели был определен в период закладки ППП по кернам, взятым с восточной стороны ствола на высоте 30-50 см от поверхности почвы; в большинстве случаев он составил 90-130 лет (табл. 1). При этом на всех ППП в момент первого обследования максимумы распределений ели по диаметру были растянуты в достаточно широком диапазоне, охватывавшем не менее 10 ступеней по 2 см. Такой большой разброс диаметров у близких по возрасту деревьев характерен для всех основных лесообразующих пород нашего региона. Специально это явление никем не изучалось, а между тем оно способно ввести исследователя в заблуждение относительно числа когорт в древесной ЦП. В этой ситуации важными критериями принадлежности всех деревьев к одному элементу леса являются именно непрерывность и мономодальность их распределения по размеру.
На ППП № 4 в момент первого обследования (рис. 1, IV, 1980 г.) максимум распределения деревьев по диаметру ствола был растянут в особенно широком диапазоне (10-42 см). Детальное изучение возрастной структуры этой ЦП ели на основе выборки модельных деревьев (250 экз.) позволило заключить, что здесь первоначально созданные культуры (посадки) ели были дополнены в середине ХХ века, когда произошло частичное изреживание сформировавшегося древостоя [4]. Дополнительно посаженные деревья ели достигли к моменту закладки ППП 5065-летнего возраста; они были выявлены в ступенях диаметра от 6 до 30 см. Однако в ходе дальнейших наблюдений выяснилось, что именно эти экземпляры выпали из насаждений в первую очередь.
Продолжая анализ состава ЦП ели нужно подчеркнуть, что в каждой из них еще за несколько десятилетий до начала исследований должны были появиться семеносящие деревья, судя по преобладающему в древостое возрасту в момент закладки ППП (табл. 1; [4]). Результатом семеношения таких деревьев должен был бы стать новый этап динамики ЦП ели – появление подроста, т.е. следующего поколения. Это отразилось бы на распределении по диаметру в виде возвышения в крайней левой части диапазона.
В работах многих авторов [5, 8, 9 и др.] описаны еловые леса, включающие более одного поколения ели, и показано, что деревья ели в них распределяются по размеру (например, по диаметру ствола) полимодально. У такого полимо-дального распределения абсолютный максимум расположен в крайней левой части (т.е. в насаждении наиболее многочисленны деревья самых малых размеров или возрастов), а у максимумов, расположенных правее, амплитуда все меньше и меньше (чем больше размер или возраст деревьев, тем меньше их численность). Подобное по-лимодальное распределение выявляли в разных природных зонах, в различных типах леса (местообитаниях) и при различной сомкнутости полога, но обычно путем включения в анализ данных о древостоях разного возраста, произрастающих на нескольких более или менее близко друг от друга расположенных участках в пределах единой, довольно большой, территории. В середине ХХ века в лесных массивах таежной зоны еще встречались подобные пространственные сочетания древостоев разного возраста, рассматриваемых как последовательные поколения в составе одного и того же лесного массива [5, 8, 10]. Процесс их возникновения можно было представить как естественное заселение деревьями участков, последовательно высвободившихся в ходе естественного же распада предшествующих древостоев. Вопреки ожиданиям, основанным на таких представлениях, на 3-х из обсуждаемых ППП (рис. 1, I-III) при первом обследовании совсем не были обнаружены ни живые, ни сухие экземпляры ели из самых малых ступеней диаметра. Это указывает на наличие препятствий для возобновительных процессов ели в этих насаждениях.
На ППП № 4 (рис. 1, IV) при первом обследовании хотя бы присутствовали единичные экземпляры ели из самых малых ступеней диаметра (0-4 см), но они вовсе не формировали максимума в левой части распределения, как это ранее теоретически предполагали для интенсивных возобновительных процессов в ЦП и нередко описывали в ходе наблюдений. Можно с большим основанием думать, что в момент закладки этой ППП ели диаметром 4 см на ней это были наиболее угнетенные экземпляры из числа дополнительно посаженных.
Таким образом, во всех рассматриваемых вторичных еловых насаждениях уже старовозрастных к началу наблюдений не было условий для успешного возобновления ели и не возникло предпосылок для восстановления ЦП в случае естественного распада древостоя или разрушительных для него внешних воздействий. На каждой ППП параметры распределения деревьев ели по диаметру постепенно менялись так же, как описывал С.А. Дыренков [8] для одного элемента леса: диапазон расширялся вправо, туда же сдвигался единственный максимум, одновременно уменьшаясь по амплитуде (рис. 1, I); общий тип распределения становился все ближе к нормальному. Все это объективно отражает постепенный текущий рост деревьев в составе элемента леса и отмирание некоторой части из них (наиболее угнетенных). Особенно интересно постепенное формирование распределения нормального типа в ЦП ели на ППП № 4 (рис. 1, IV), куда еще до начала наблюдений была искусственно добавлена новая когорта деревьев, которая могла сформировать здесь еще один элемент леса (и второй максимум в общем распределении деревьев по диаметру). Однако эта когорта, как показывает динамика состояния слагающих ее деревьев, включилась в состав ранее существовавшего элемента леса. В нем она оказалась самой угнетаемой частью древостоя (рис. 2, А) и отреагировала на это более высокими темпами отмирания, чем у деревьев, которые по календарному возрасту были значительно старше. Невозможность формирования нового элемента леса была, возможно, обусловлена сложными фитоценотическими условиями, то есть нехваткой необходимых для этого свободных ресурсов в сообществе.
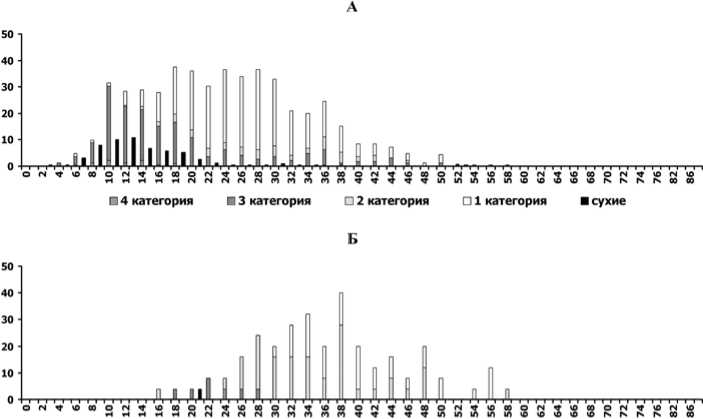
в
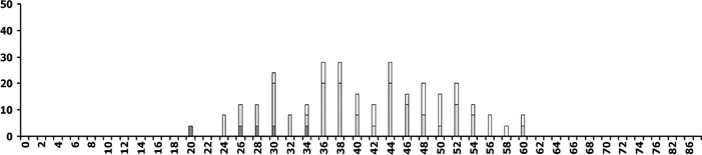
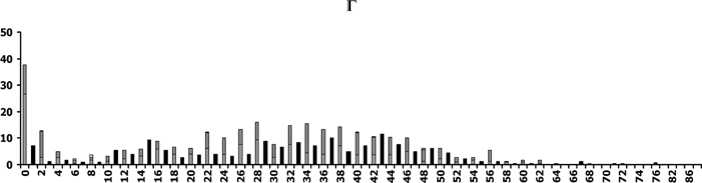
Рис. 2. Распределение деревьев ели по диаметру ствола и категориям состояния [13] на разных этапах динамики ценопопуляций:
А - расцвет господствующей когорты деревьев (ППП № 4 в 1980г.), Б, В - перед распадом господствующей когорты деревьев (ППП №№ 2 и 3 в 1997г.), Г - в ходе распада господствующей когорты (ППП № 4 в 2011 г.). По оси абсцисс - ступени диаметра ствола (см) на высоте 1,3 м, по оси ординат - число деревьев (шт./га)
Напомним, что на всех 4-х ППП при первом обследовании сухие деревья ели были единичны [4]. В это время численно преобладали деревья 1-й категории санитарного состояния [13], т.е. без признаков ослабления (рис. 2, А), причем они обнаруживали почти нормальное распределение по ступеням диаметра. В течение следующих 30 лет на всех ППП засыхали только тонкомерные (с малым диаметром ствола) деревья [4], несмотря на случавшиеся экстремальные погодные явления и вспышки численности вредителей [12], способные ухудшить состояние крупных деревьев. Позже признаки ослабления постепенно распространялись и среди крупных деревьев, в результате чего в ЦП стали преобладать деревья 2-3-й категорий (рис. 2, Б и В). Теперь уже их распределение по ступеням диаметра приобрело характер нормального, тогда как распределение деревьев 1-й категории стало более равномерным. В последние годы равномерно во всех ступенях диаметра стали появляться усыхающие (4-я категория) и сухие деревья ели (рис. 2, Г). Деревьев 1-й и 2-й категорий в этом элементе леса практически не осталось, а к нормальному типу приблизилось распределение сильно ослабленных (3-я категория). Все это указывает на скорый окончательный распад древостоя в состарившемся элементе леса [2]. Другие признаки скорого распада, заметные на той же гистограмме, симметричность мономодаль-ного распределения деревьев по диаметру и очень малая амплитуда его максимума. Судя по наблюдениям, в 2009-2011 гг. уже на 2-х ППП (рис. 1, I и IV) древостои достигли этапа распада.
Еще одна важная тенденция в динамике распределений ели по диаметру –для большинства ППП за 30 лет наблюдений не были выявлены существенные изменения в крайней левой части (в самых малых ступенях диаметра), отражающей результаты процесса возобновления. Новые деревья ели не появились в исследуемых ЦП совсем (рис. 1, I и III) или появлялись единично на небольшой период времени (рис. 1, II), после чего отмирали (это ясно из того, что последующими обследованиями не были выявлены сдвиги новых максимумов распределения вправо). Некоторое отличие можно заметить на ППП № 4 (рис. 1, IV): одновременно с началом интенсивного распада первоначального древостоя (2011 г.) там было отмечено появление довольно многочисленных тонкомерных экземпляров ели и наконец-то стал заметен абсолютный максимум численности в крайней левой части распределения. Однако все эти тонкомерные экземпляры относятся к категориям сильно ослабленных или усыхающих (рис. 2, Г), поэтому не приходится ожидать, что они смогут сформировать долговечный элемент леса, способный заместить предшествующий.
Таким образом, за период, охваченный систематическими наблюдениями, процессы возобновления ели в обсуждаемых насаждениях либо не стали более интенсивными, чем ранее, либо, несмотря на интенсивность, не оказались более успешными. И по-прежнему возобновление не создало предпосылок для восстановления ЦП ели после отмирания деревьев, обусловленного их старостью или катастрофически разрушительными внешними воздействиями.
Выводы: в ходе 30-летних наблюдений на ППП в еловых старолесьях Подмосковья удалось показать характерные черты естественной динамики спелых еловых насаждений. Созданные человеком или возникшие в результате его влияния, они уже не воспроизведут себя в следующем поколении. Подрост ели, даже многочисленный, не выдерживает длительного пребывания под пологом леса и не может сформировать основу будущего древостоя. Пока еще невозможно с уверенностью прогнозировать теоретически, на основе изучения одного лишь строения древостоев, какова закономерная для средней полосы европейской части России спонтанная динамика леса. Только проводя многолетние наблюдения динамики древостоев можно понять ее закономерности и узнать действительные лесоводственные свойства основных лесообразующих пород. Это совершенно необходимо для разработки рациональных способов ведения лесного хозяйства.
Список литературы Закономерности динамики еловых насаждений в старолесьях Подмосковья
- Абатуров, А.В. Динамика ельников на территории лесопаркового защитного пояса Москвы/А.В. Абатуров, В.В. Антюхина//Динамика хвойных лесов Подмосковья. -М.: Наука, 2000. С. 86-109.
- Абатуров, А.В. Возрастная структура и состояние лесов Ближнего Подмосковья//Мат-лы 5-й Междунар. конф. «Проблемы лесной фитопатологии и микологии». Москва, 7-10(13) октября 2002 г. -М.: ВНИИЛМ, 2002. С. 13-16.
- Абатуров, А.В. 150 лет Лосиноостровской лесной даче. Из истории национального парка «Лосиный Остров»/А.В. Абатуров, О.В. Кочевая, А.И. Янгутов. -М.: Аслан, 1997. 237 с.
- Абатуров, А.В. Естественная динамика леса на постоянных пробных площадях в Подмосковье/А.В. Абатуров, П.Н. Меланхолин. -Тула: Гриф и К, 2004. 336 с.
- Воропанов, П.В. Ельники Севера. -М.-Л.: Гослесбумиздат, 1950. 179 с.
- Гиляров, А.М. Популяционная экология. -М.: Изд-во МГУ, 1990. 215 с.
- Демографический энциклопедический словарь. -М.: Советская энциклопедия, 1985. 610 с.
- Дыренков, С.А. Структура и динамика таежных ельников. -Л.: Наука, 1984. 173 с.
- Ивашкевич, Б.А. Девственный лес, особенности его строения и развития//Лесн. хоз-во и лесн. пром-сть. 1929. № 10-12. С. 36-44.
- Казимиров, Н.И. Ельники Карелии. -Л.: ЛО «Наука», 1971. 138 с.
- Мозолевская, Е.Г. Очаги короеда-типографа в ельниках национального парка «Лосиный остров»/Е.Г. Мозолевская, В.А. Липаткин, А.Н. Щербаков, Т.В. Шарапа//Экология, мониторинг и рациональное природопользование Науч. тр. МГУЛ. Вып. 307 (1). -М.: МГУЛ, 2001. С. 9-19.
- Речан, С.П. Леса Северного Подмосковья/С.П. Речан, Т.В. Малышева, А.В. Абатуров, П.Н. Меланхолин. -М.: Наука, 1993. 314 с.
- Санитарные правила в лесах Российской Федерации/Федер. служба лесного хоз-ва России. -М., 1998. 25 с.
- Третьяков, Н.В. Закон единства в строении древостоев. -М. -Л., 1927. 113 с.
