Закономерности распространения сапонинов и экдистероидов в растениях: хемотаксономический и молекулярно-филогенетический подходы

Автор: Шадрин Д.М., Пылина Я.И., Родионов А.В., Володина С.О., Ткаченко К.Г., Володин В.В.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Биологические ресурсы: флора
Статья в выпуске: 1-3 т.12, 2010 года.
Бесплатный доступ
С использованием хемотаксономического и молекулярно-филогенетического подходов изучено распределение стероидных и тритерпеновых гликозидов в сем. Fabaceae Lindl. и экдистероидов в сем. Asteraceae. Установлено, что большинство представителей сем. Fabaceae содержат тритерпеновые гликозиды, стероидные гликозиды обнаружены только у некоторых филогенетически обособленных таксонов в трибах Loteae, Thermopsideae и Trifolieae. Выявлены существующие филогенетические связи между видами сем. Asteraceae, у которых экдистероиды, возможно, выполняют экологическую роль. Разработан научно обоснованный прогноз поиска ресурсных видов растений - продуцентов вторичных метаболитов мевалонатного пути биосинтеза сапонинов и экдистероидов.
Стероидные гликозиды, тритерпеновые гликозиды, экдистероиды, хемотаксономия, молекулярная филогения
Короткий адрес: https://sciup.org/148199058
IDR: 148199058 | УДК: 581.192:57.088.6:582
Regularities of saponins and ecdysteroids distribution in plants: chemotaxonomical and molecular phylogenic approaches
With the use of chemotaxomomical and molecular-phylogenetic approaches distribution of steroid and triterpenic glycosides in family Fabaceae Lindl. and ecdysteroids in family Asteraceae is studied. It is established, that the majority of representatives family Fabaceae contain triterpenic glycosides, steroid glycosites are found out only in the some phylogenetic isolated taxons in tribes Loteae, Thermopsideae and Trifolieae. Existing phylogenetic communications between kinds of family Asteraceae are revealed, at which ecdysteroids, probably, carry out an ecological role. Scientifically proved forecast of search the resource kinds of plants - producers of secondary metabolites of mevalonant ways of saponines and ecdysteroids biosynthesis are developed.
Текст научной статьи Закономерности распространения сапонинов и экдистероидов в растениях: хемотаксономический и молекулярно-филогенетический подходы
проекта INTAS с участием нашей лаборатории была сделана попытка построения молекулярно-филоге-нетического древа сем. Caryophyllaceae [1]. Более обширные исследования на основе хемотаксономического и молекулярно-филогенетического подходов в изучении распределения алкалоидов и непротеиногенных аминокислот на примере сем. Fabaceae были проведены Винком [2].
Настоящая работа направлена на выявление связей между распространением двух биогенетически близких групп вторичных метаболитов, связанных общим мевалонатным путем биосинтеза – сапонинов (на примере сем. Fa-baceae ) и экдистероидов (на примере трибы Car-dueae, сем. Asteraceae ) и филогенетической классификацией растений, что позволит оценить их значимость в качестве хемотаксономических маркеров.
Для определения изучаемых нами вторичных метаболитов растений использованы известные экспрессные физико-химические и биологические методы тестирования растений (сапонины): способность к пенообразованию, гемолизу эритроцитов, а также ТСХ- и ВЭЖХ-хроматография. Для установления тонкой структуры соединений использовали методы масс-спектрометрии и ЯМР-спект-роскопии. Для выявления закономерностей распространения сапонинов и экдистероидов реконструирована молекулярная филогения сем. Fabaceae и трибы Cardueae, сем. Asteraceae. При проведении анализа близкородственных таксонов использовали последовательности внутренних транскрибируемых спейсеров ITS1 и ITS2 из базы данных GenBank. Последовательности генов выравнивались вручную и с помощью программы ClustalW, входящей в пакет программ MEGA 4. Нами использованы два метода: метод максимальной парсимонии и метод объединения ближайших соседей. Генетические расстояния рассчитывались исходя из модели Kimura-2. Данная модель успешно используется для анализа близкородственных таксонов [3].
На содержание тритерпеновых (ТГ) и стероидных гликозидов (СГ) исследовано 93 образца 46 видов растений из 14 родов (см. табл.). Установлено, что подавляющее большинство изучаемых видов содержит тритерпеновые гликозиды. Стероидные гликозиды обнаружены у представителей родов Trigonella (триба Trifo-lieae), Thermopsis (триба Termopsideae) и Cor-onilla (триба Loteae). Распределение сапонинов двух классов, стероидных и тритерпеновых часто является взаимоисключающим, что может указывать на более экономичное использование защитных ресурсов растений. Тем не менее, в литературе имеются сведения о том, что вид Vicia tenuifolia L. содержит и тритерпеновые, и стероидные гликозиды [4]. Указанный выше факт требует дополнительных исследований, доказывающих возможность одновременного присутствия двух типов сапонинов в растениях.
Таблица. Распространение сапонинов в сем. Fabaceae Lindl.
|
Таксон |
Литературные данные |
Экспериментальные данные |
|||||
|
общее кол-во видов |
содержат: |
общее кол-во видов |
содержат: |
||||
|
ТГ |
СГ |
сумма |
ТГ |
СГ |
|||
|
Триба Hedysareae |
|||||||
|
род Hedysarum L. |
6 |
3 |
- |
3 |
2 |
2 |
- |
|
род Onobrichus Hill. |
1 |
- |
- |
1 |
4 |
4 |
- |
|
Триба Loteae |
|||||||
|
род Anthyllis L. |
2 |
1 |
1 |
- |
1 |
1 |
- |
|
род Coronilla |
1 |
- |
1 |
- |
1 |
- |
1 |
|
Триба Tripholeae |
|||||||
|
род Medicago L. |
9 |
9 |
- |
- |
3 |
3 |
- |
|
род Melilotus L. |
2 |
1 |
1 |
- |
2 |
2 |
- |
|
род Tripholium L. |
4 |
3 |
- |
1 |
9 |
9 |
- |
|
род Trigonella L. |
5 |
- |
4 |
1 |
2 |
- |
2 |
|
род Ononis L. |
4 |
- |
- |
4 |
- |
- |
- |
|
Триба Genisteae |
|||||||
|
род Lupinus L. |
1 |
1 |
- |
- |
1 |
1 |
- |
|
род Chamaecytisus Link. |
1 |
- |
- |
1 |
- |
- |
- |
|
Триба Phaseoleae |
|||||||
|
род Phaseolus L. |
1 |
1 |
- |
- |
1 |
1 |
- |
|
род Glicine Willd |
1 |
1 |
- |
- |
- |
- |
- |
|
Триба Galegeae |
|||||||
|
род Astragalus L. |
24 |
13 |
- |
11 |
4 |
4 |
- |
|
род Caragana Fabr |
4 |
1 |
- |
3 |
1 |
1 |
- |
|
род Galega L. |
2 |
- |
- |
2 |
- |
- |
- |
|
род Glycyrriza L. |
4 |
4 |
- |
- |
- |
- |
- |
|
род Halimadendron DC. |
1 |
- |
- |
1 |
- |
- |
- |
|
род Oxitropis DC. |
9 |
9 |
- |
- |
2 |
2 |
- |
|
Триба Vicieae |
|||||||
|
род Lathyrus L. |
1 |
1 |
- |
- |
4 |
4 |
- |
|
род Vicia L. |
8 |
8 |
1 |
- |
4 |
4 |
- |
|
род Pisum L. |
1 |
1 |
- |
- |
1 |
1 |
- |
|
Триба Termopsideae |
|||||||
|
род Thermopsis R.Br. |
1 |
- |
- |
1 |
3 |
- |
3 |
|
род Baptisia |
1 |
1 |
- |
- |
2 |
2 |
- |
|
Сумма |
94 |
58 |
8 |
29 |
47 |
41 |
6 |
Наличие стероидных гликозидов в трибе Trifolieae в роде Trigonella и у вида Melilotus tauricus [5], в трибе Loteae в роде Coronilla и у вида Anthyllis macrocephala [6], а в трибе Ther-mopsideae – только в роде Thermopsis, но не у других представителей этой трибы. Это позволило заметить, что в сем. Fabaceae стероидные гликозиды присущи видам и родам, не связанным монофилитическими отношениями на уровне триб, к которым они принадлежат (рис. 1) и обнаружены только у некоторых филогенетически обособленных таксонов в трибах Loteae, Thermopsideae и Trifolieae. При этом в трибе Trifolieae, как указанно выше, стероидные гликозиды обнаружены только в двух родах (Trigonella и Melilotus), которые на молекулярно-филогенетической кладограмме образуют отдельную кладу, обособленную от других представителей этой трибы, что в свою очередь отражает эволюцию этих таксонов. На данном этапе исследования выявленная нами закономерность не может быть истолкована однозначно. С одной стороны, ее можно объяснить тем, что гены, кодирующие биосинтез ферментов, ответственных за синтез стероидных гликозидов, были приобретены в процессе эволюции в нескольких таксонах, не зависимо друг от друга.
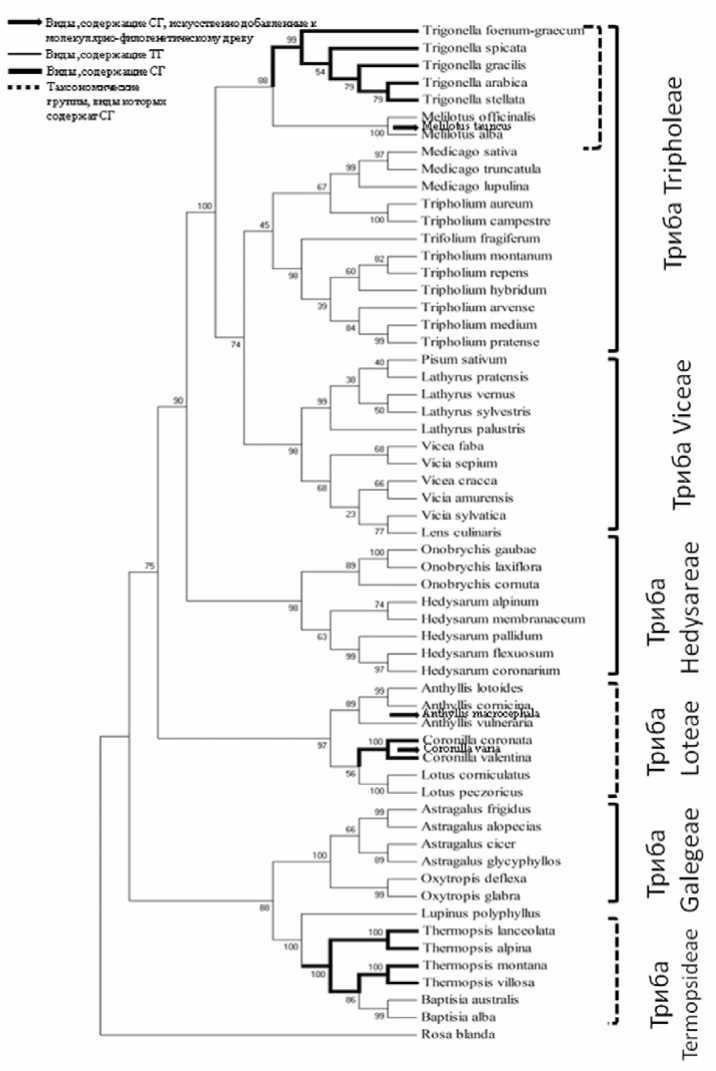
Рис. 1. Филогенетическое древо растений сем. Fabaceae Lindl.
С другой стороны, не исключается гипотеза о том, что эти гены, были приобретены на ранних этапах эволюции цветковых растений, однако их экспрессия происходит только в определенных таксонах, что находит свое отражение и на уровне изучаемого семейства.
Установленный факт, что тритерпеновые гликозиды являются характерными для подавляющего числа видов наследственных Caesalpinioideae /Mimosoideae и большинства триб Papilionoideae, куда входят исследованные нами виды, указывает на генетическую предрасположенность видов сем.
Fabaceae к биосинтезу сапонинов именно этой группы.
Закономерности распространения тритерпеновых и стероидных гликозидов среди представителей сем. Fabaceae использованы нами для составления хемотаксономического прогноза их обнаружения во флоре Европейского Северо-Востока России. Таким образом, большинство видов флоры содержит тритерпеновые гликозиды, стероидные гликозиды можно искать в ограниченных таксонах, указанных выше. Ранее считалось, что не существует связи между распространением экдистероидов и филогенетической классификацией растений, поскольку эк-дистероиды встречаются как в филогенетически близких, так и удаленных семействах. Внутри семейств и даже внутри родов могут встречаться как экдистероидсодержащие, так и лишенные этих соединений виды [7]. Работами сотрудников лаборатории биохимии и биотехнологии Института биологии было показано, что биохимический признак наличия экдистероидов на внутрисемейственном уровне закреплен в трибах, хотя эта закономерность и не носит абсолютного характера. Сказанное относится к концентрациям экдистероидов в растениях, обнаруживаемым с помощью биотеста на культуре клеток Drosophila melanogaster (т.е. гормонально- активным концентрациям по отношению к насекомым). При этом оказалось, что следовые количества экдистероидов, не активные в биотесте, но обнаруживаемые с помощью радиоиммунного анализа в концентрации менее 4 мкг/г содержат едва ли не все виды растений. Это свидетельствует о том, что гены, кодирующие ферменты биосинтеза экдистероидов присутствуют у всех растений, но их выраженная экспрессия происходит только в группах близкородственных видов, ограниченных определенными родами и трибами, в которых, по-видимому, экдистероиды выполняют экологическую функцию.
В сем. Asteraceae, насчитывающем по разным источникам 20-25 тыс. видов, экдисте-роиды обнаружены только у представителей трибы Cardueae. На основании данных GenBankа нами было построено молекулярнофилогенетическое древо трибы Cardue подсемейства Lactucoideae. Эта триба включает две подтрибы Carduinae (B), виды которой содержат экдистероиды только в следовых количествах, и Centaureinae (A), которая, по данным В.В. Володина и И.Ф. Чадина, включает виды с экологически значимой концентрацией, т.е. «положительные» в биотесте (рис. 2) [8].
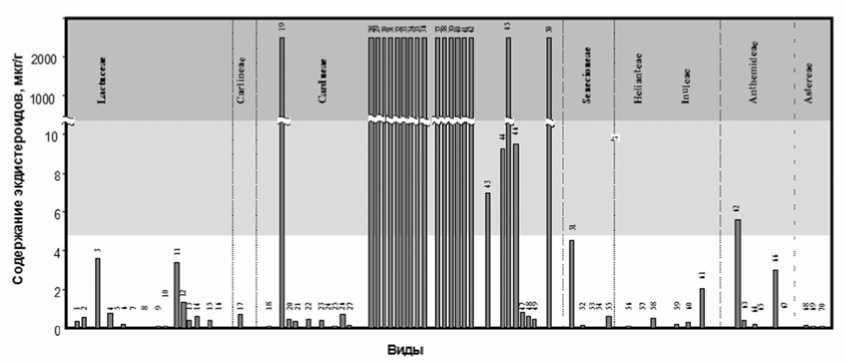
Рис. 2. Распространение экдистероидов в семействе Asteraceae :
темная окраска – высокие концентрации, обнаруживаются традиционными методами (положительный ответ в биотесте); более светлая окраска – умеренные концентрации; светлое поле – низкие концентрации, обнаруживаются РИА (отрицательный ответ в биотесте)
LACTUCOIDEAE: Lactuceae: 1 - Hieracium sibiricum , 2 - H . umbellatum , 3 - Leontodon autumnalis , 4 - Picris hei-racioides , 5 - P . hieracioides , 6 - P . umbellatum , 7 - P . umbellatum , 8 - Scorzonera humilis , 9 - Lactuca sativa , 10 - L . tatarica , 11 - Crepis paludosa , 12 - C . praemorsa , 13 - C . sibirica , 14 - C . tectorum , 15 - Taraxacum officinale , 16 - Sonchus asper , Carlineae: 17 - Carlina sp . Cardueae: Carduineae 18 - Arctium tomentosum , 19 - Saussurea latifolia , 20 – S . alpina , 21 - S . parviflora , 22 - Carduus nutans , 23 - Cirsium heterophyllum , 24 - C . oleraceum , 25 - C . palustre , 26 - C . setosum , 27 - C . vulgare , Cardueae: Centaurineae: 28 - Serratula algida , 29 - S . centauroides , 30 - S . coro-nata , 31 - S . inermis , 32 - S . lyratifolia , 33 - S . procumbens , 34 – S . quinquefolia , 35 - S . sogdiana , 36 – S . xeran-themoides , 37 - Rhaponticum carthamoides , 38 - Rh . integrifolium , 39 - Rh . karatavicum , 40 - Rh . lyratum , 41 - Rh . nitidum , 42 - Rh . pulchrum , 43 - Acroptilon repens , 44 - Centaurea scabiosa , 45 - C . rothrokii , 46 - C . phrygia , 47 - C . americana , 48 - C . jacea , 49 - C . fischeri , 50 - Amberboa moschata ;
ASTEROIDEAE: Senecioneae: 51 – Ligularia sibirica , 52 - Petasites spurius , 53 - Cacalia hastata , 54 - Senecio nemoriensis , 55 - S . vulgaris ; Heliantheae: 56 - Bidens cernua , 57 - B . tripartita , 58 - Ambrosia sp .; Inuleae: 59 - An-tennaria dioica , 60 - Omalotheca sylvaticum , 61 - Inula salicina L . Anthemideae: 62 - Achillea millefolium , 63 - A . ptarmica , 64 - Matricaria recutita , 65 - M . suaveolens , 66 - Leucanthemum vulgare , 67 - Artemisia campestris ; As-tereae: 68 - Solidago virgaurea , 69 - Erigeron acris , 70 - E . canadensis
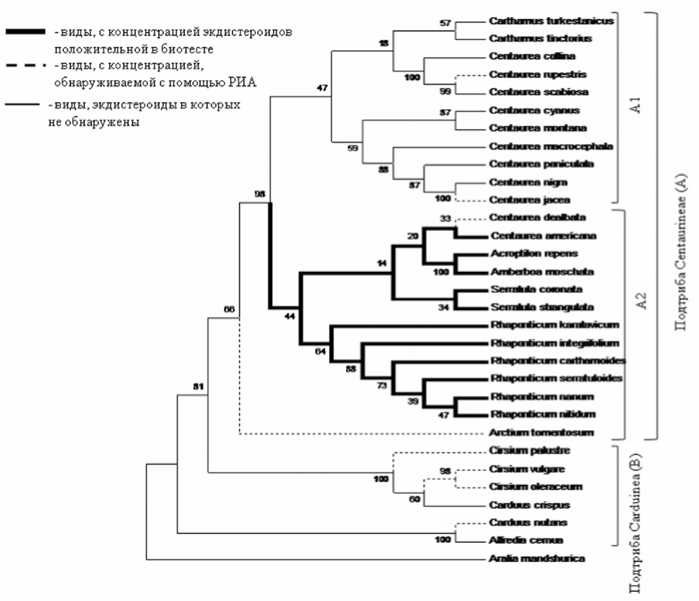
Рис. 3. Филогенетическое древо растений трибы Cardueae сем. Asteraceae Dumort.
Подтриба Centaureinae образует обособленную кладу, включающую две подклады (A1 и A2) с высоким коэффициентом Бутстрепа (рис. 3). Следует отметить, что виды, давшие отрицательную реакцию в биотесте, представлены двумя родами Carthamus и Centaurea и образуют подкладу А1. На молекулярнофилогенетическом древе подкладу А2 образовали виды, давшие положительную реакцию в биотесте и представленные пятью родами: Rhaponticum, Acroptilon, Amberboa, Serratula, Centaurea. Причем в роду Centaurea из 11 исследованных видов – только у представителя C.americana. Другой из исследованных Сарке-ром и Дайнаном в этом роде вид Centaurea mo-schata, в котором было обнаружено высокое содержание экдистероидов [9], по флоре СССР, считается видом другого рода - Amberboa mo-schata, филогенетически близкого роду Acropti-lon. Исходя из данного замечания, уже не кажется неожиданным обнаружение высокой концентрации экдистероидов как в образцах растений Acroptilon repens, так и в Amberboa mo-schata. Полученные данные соответствуют представлению M. Dittrich (1977 год) о разделении на основании палинологических, морфологических и анатомических признаков подтрибы Centaureinae на группы родов, согласно которому Acroptilon входит в одну группу с родами Rhaponticum и Serratula, а роды Carthamus и Centaurea относятся к другой группировке родов [10]. В свою очередь, на построенном нами молекулярно-филогенетическом древе представители родов Acroptilon и Amberboa имеют общего, предположительно, экдистероидсодер-жащего предка. Следовательно, данные распространения фитоэкдистероидов в сочетании с методами молекулярной филогении позволяют выявить существующие филогенетические связи между видами, у которых экдистероиды, возможно, выполняют экологическую роль.
Выводы: использование хемотаксономи-ческого и молекулярно-филогенетического подходов позволило изучить закономерности распространения двух классов вторичных метаболитов в растениях и оценить их значимость в качестве хемотаксономических маркеров и разработать научно обоснованный прогноз поиска ресурсных видов растений – продуцентов вторичных метаболитов мевалонатного пути биосинтеза сапонинов и экдистероидов.
Работа выполнена при финансовой поддержке Программы Отделения биологических наук РАН «Биологические ресурсы России, оценка состояния и фундаментальные основы мониторинга» (проект « Состояние ресурсов полезных растений европейского Северо-Востока России: мониторинг и разработка биотехнологических подходов по рациональному использованию и воспроизводству».
Список литературы Закономерности распространения сапонинов и экдистероидов в растениях: хемотаксономический и молекулярно-филогенетический подходы
- Distribution of phytoecdysteroids in the Caryophyllaceae/L. Zibareva, V. Volodin et al.//Phytochemistry. -2003. -Vol. 64. -P. 499-517.
- Wink, M. Evolution of secondary metabolites from an ecological and molecular phylogenetic perspective//Phytochemistry. -2003. -Vol. 64. -P. 3-19.
- Nei, M. Molecular Evolution and Phylogenetics/M. Nei, S. Kumar. -NY: Oxford University Press, 2000. -333 p.
- Куваев, И.Б. Предварительная химическая оценка лекарственных растений тибетской медицины, произрастающих в Забайкалье/И.Б. Куваев, К.Ф. Блинова//Вопросы фармакогнозии. -1961. -Вып. 1. -С. 213-262.
- Triterpene and steroid saponins isolated from two Melilotus species/Khodakov, G.V., Akimov, Y.A., Shashkov, A.S., et al.//Adv-Exp-Med-Biol. -1996. -P. 405211-40522.
- Галкин, М.А. Дикорастущие полезные растения Северного Кавказа/М.А. Галкин, А.Л. Казаков. -Ростов-на-Дону, 1980. -128 с.
- Lafont, R. Phytoecdysteroids: structures, occurrence, biosynthesis and possible ecological significance/R. Lafont, A. Bouthier, I.D. Wilson//Conf. Incect Chem. Ecol.: Programme and Abstracts. -Tabor, 1991. -P.197-214.
- Фитоэкдистероиды/Под ред. В.В Володина. -СПб: Наука, 2003. -293 с.
- Occurrence of Ecdysteroids in the Genus Centaurea (Compositae): 20-Hydroxyecdysone from Centaurea moschata/S. Sarker, L. Dinan et al.//Biochemical Systematics and Ecology. -1997. -Vol. 25. -P. 367-368.
- Dittrich, M. Cynareae-systematic review//The biology and chemistry of Compositae/Eds V.H. Heywood, I.B. Harborne, B.L. Turner. -London, 1977. -P. 999-1015.
