Заметки о современном состоянии гидробиологии континентальных водоемов

Автор: Алимов А.Ф.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Водные экосистемы
Статья в выпуске: 1 т.8, 2006 года.
Бесплатный доступ
В статье рассмотрены основные положения продукционной гидробиологии и теории биологической продуктивности и некоторые применения этих положений для решения теоретических и практических задач. Обращено внимание на необходимость разработки теории функционирования водных экосистем и слабую изученность потоков вещества и особенно потоков информации в экосистемах, е также на необходимость знания пространственно-временных связей в экосистемах различных водоемов. Такие знания необходимы для понимания механизмов функционирования водных экосистем.
Короткий адрес: https://sciup.org/148197823
IDR: 148197823 | УДК: 574.5
Notes on the current status of hydrobiology of continental water-bodies
The article deals with the main conceptions of production hydrobiology and the theory of biological productivity and some applications of these conceptions for the resolution of theoretical and practical tasks. Emphasis is placed on the necessity to develop a theory of the functioning of aquatic ecosystems and on the scanty knowledge of matter flows and particularly of information flows in ecosystems and on the necessity to gain knowledge of spatio-temporal relationships in ecosystems of different water-bodies. Such knowledge is needed for understanding the mechanisms of the functioning of aquatic ecosystems.
Текст научной статьи Заметки о современном состоянии гидробиологии континентальных водоемов
Зоологический институт РАН, г. Санкт-Петербург
В статье рассмотрены основные положения продукционной гидробиологии и теории биологической продуктивности и некоторые применения этих положений для решения теоретических и практических задач. ОТращено внимание на необходимость разработки теории функционирования водных экосистем и слабую изученность потоков вещества и особенно потоков информации в экосистемах, т также на необходимость знания пространственно-ттеменных связей в экосистемах различных водоемов. Такте знания необходимы для понимания механизмов функционирования водных экосистем.
В силу разных причин водные, главным образом пресноводные экосистемы оказались наиболее хорошо изученными. В гидробиологии раньше, тем в экологии суши, ттчалось изучение надорганизменных форм организа-ции биологических систем (планктона, бентоса , нектона и т.д.). Многие основные понятия , широко используемые в современной экологии, такие, например, так биомасса, биоценоз , продукция, трофические уровни, потоки энергии, тещества и другие, тыли сформулированы на основании изучения жизни в различных водоемах и водотоках.
Продукционная гидробиология - раздел гидробиологии, изучающий процессы образования (продукции) органических веществ в водоемах и их использования в водных эко -системах и направленный на разработку те -ории биологической продуктивности водо-емов, ттдотоков и теории функционирования водных экосистем. Основы продукционной гидробиологии были заложены Г.Г. Винбер-гом в начале 30-х годов XX столетия.
При продукционных исследованиях вод -ных экосистем используется балансовый под -ход и энергетический принцип. Баланс органических веществ занимает особое место в экосистемах. Он охватывает широкий круг разнородных и сложных процессов в водо-емах, с ним связаны балансы биогенных и многих других элементов. Его нельзя рассматривать как простую разность между поступ- лением органических веществ в водоем и выносом их из него, по скольку органические вещества могут синтезироваться в самом во -доеме и использоваться в его экосистеме. Поэтому большое значение имеют исследо-вания скорости образования (продукция) и разрушения (деструкция) органических веществ в водоемах.
Энергетический принцип изучения тро-фических связей, б иотического круговорота веществ и биологической продуктивности водоемов отражает фундаментальное поло -жение о том, что энергия в цепи трофических превращений не исчезает, а может переходить лишь из одной формы в другую. Каждое из многообразных явлений, составляющих биотический баланс в экосистемах, занимает определенное положение по отноше-нию к потоку энергии, направленному от организмов-накопителей к организмам-потребителям .
В соответствие с такой энергетической трактовкой Г.Г. Винтерг предложил выразить биотический баланс в виде простого равен -ства:
ArR = ± P, где: A - новообразование органических веществ в водоеме, R - превращения органических веществ, твязанные с процессами метаболизма у гидробионтов, Р - продукция. ттак ± перед последним членом балансово- го равенства говорит о том, что баланс органических веществ может быть положитель-ным и отрицательным.
В принципе, каждый из членов балансового равенства следует рассматривать как ре -зультат или сумму соответствующих функций отдельных организмов. Количественное выражение участия популяций отдельных ви-дов в общих процессах превращения веществ и трансформации энергии в водоемах воз -можно лишь при знании скоростей их роста, скорости обмена у них, величин продукции популяций и сообществ организмов конкрет-ных видов , которая представляет собой скорость прироста их биомассы.
В результате многочисленных исследова-ний было убедительно показано, что скорости обмена ( R ), ооста ( dW / dt ), питания ( C) , экскреции ( E) , размножения животных, как и скорости увеличения численности в их по -пуляциях ( rm ), ооходятся в закономерной связи с массой организмов. Эти связи выражаются степенными уравнениями, о вполне определенными и биологически значимыми ве-личинами параметров:
R = a i W °' 75 ; dW / dt = a 2 W b- a 3 W b 1 ;
Приведенные зависимости были получе-ны при использовании сведений по эколо -гической физиологии отдельных видов, что позволило количественно выразить важней -шие жизненные функции организмов, определить изменения их скоростей, о зависимости от изменений условий среды. Это позволило выделить, как раздел экологии, функциональную экологию животных [1], которая на основе аутэкологических исследований от -дельных видов количественно оценивает зна-чение популяций и сообществ животных в процессах биологической продуктивности, превращения веществ и трансформации энер-гии в экосистемах.
Из приведенных выше уравнений видно, что именно с массой организмов связаны все наиболее значимые функциональные харак-теристики животных. Сказанное соответствует представлениям о том, что с концентрацией массы живого вещества закономерно связана его функциональная активность [7]. Массу всех представителей органического мира можно рассматривать как меру скорое -ти или интенсивности обмена у них. Это отличает живые организмы от предметов не -живой природы. В месте с тем масса живых организмов, кок и масса тел неживой природы , является мерой их инерционных и гравитационных свойств. Следовательно, масса биологических объектов является одно -временно и мерой инерции, гравитации, и мерой скорости или интенсивности обмен -ных процессов, обеспечивающих жизнедеятельность организмов.
Один из важнейших вопросов, стоящих перед биологами и экологами - сколько видов может обитать на некоторой территории? Такие исследования по отношению к неко-торым конкретным немногочисленным труп -нам водных животных с разной степенью пе-риодичности ведутся в течение многих лет [например, 9,10,11,12]. В результате было показано , что число видов в пределах отдельных таксономических групп животных и ра-стений имеет тенденцию к увеличению с
C = a 3 W 0.6 ; E = a 4 W b 2 ; rm = a 5 W "С"
уменьшением географической широты и с увеличением площади водоема.
Количество видов в пределах экосистем конкретных водоемов определяется различ-ными факторами, в том числе и историей формирования фауны. Воесте с тем по отношению к каждой экосистеме важнейшие фак -торы, которые могут определять возможное количество видов, оходящих в ее состав, - ото территория или обитаемое пространство и орофические условия. Порвый фактор может быть оценен площадью или объемом водо-ема, второй - воличиной его первичной продукции .
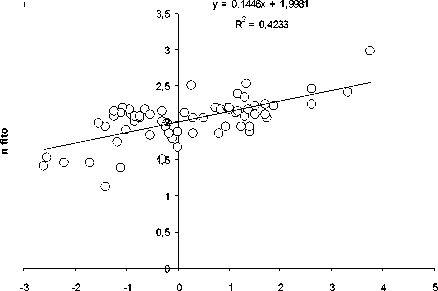
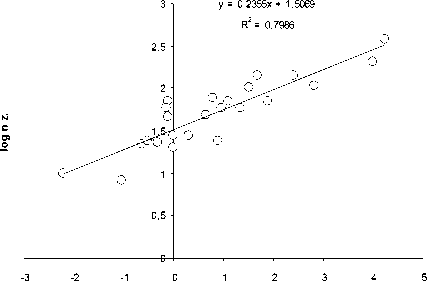
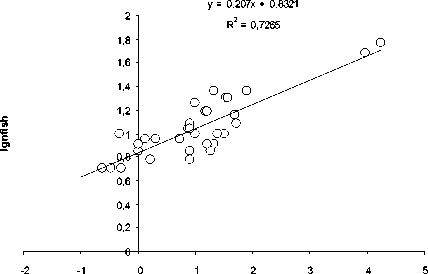
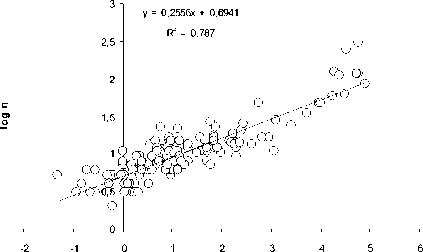
Как видно из рис. 1, количество видов ( n ) ф ито-, зоопланктона, бентоса и рыб в озерных экосистемах, н езависимо от типа водоемов , их географического положения и трофического статуса возрастает с увеличением ооощади зеркала озер ( S, км2):
n = aS b (1)
logS,km2
logS
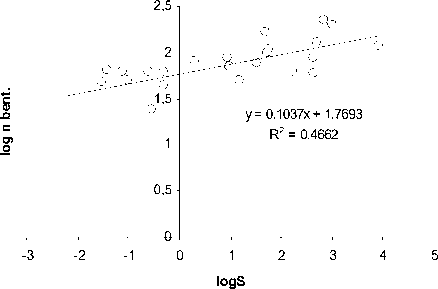
lgS
log S
Рис. 1. Зависимость логарифма числа видов (log n) фитопланктона (А), зоопланктона (Б), бентоса (В), рыб (Г) и общего числа видов (Д) от логарифма площади ( км 2) (log S) различных озер
При этом величина показателя степени в уравнении (1) составляет 0.26.
Пока нет биологически сколько - нибудь обоснованной интерпретации такой величи-ны показателя степени в уравнении связи числа видов организмов с площадью водо-ема. Была попытка представить величину этого показателя равной 0.25 [10], т.е. такой же, как и средней для всего животного мира в уравнении связи интенсивности обмена с массой животных. Обоснованность возможности такой величины показателя степени была оценена с позиций статистики [8]. Однако полученные результаты пока не оказа-лись убедительными для биологического обо -снования величины показателя степени рав-ной 0,25.
Число видов в сообществах гидробионтов возрастает по мере увеличения объема воды в водоемах ( V , км3) [3]:
n = a i V b 1 (2)
Из уравнений (1,2) следует, что ккличкство видов в водоемах на единице площади или в единице объема воды находится в обратной зависимости от их площади или объема:
n/S = aS b 1; n/V = a 1 V b Г 1
Следовательно, крупные по размерам водоемы оказываются, по сравнению с небольшими водоемами, как бы менее насыщены видами. Это позволяет предполагать, что, например , именно первые более удобны для инвазии в них новых видов. В этом отношении показательно, что количество видов ин-тродуцентов в Великих Северо-Американских озерах увеличивалось с возрастанием их площади.
Поскольку средняя глубина озера ( h ) может быть определена из соотношения между объемом воды в озере и его площадью ( h = V/S ) , из уравнений (2) и (1) следует: n = a 2 h b 3 , т.е. к озср ах с большей средней глубиной при прочих равных условиях следует ожидать большего количества видов.
Кроме того, кисло видов, иск в отдельных подсистемах и в целом в экосистемах, зависит от величины первичной продукции план -ктона. Наибольшее количество видов гетерот рофных организмов отмечено в водоемах, в которых величина первичной продукции планктона близка к 1400 ккал/м2 за год [3]. В ккдоемах большей или меньшей продуктив-ности количество видов снижается.
Когда структура сообществ животных ха -рактеризуется просто числом видов, не принимаются во внимание взаимоотношения между ними, керяется информация о редкости одних видов и обычности других, т.е. о сложности организации структуры сообществ организмов.
Для оценки сложности структуры сооб-ществ животных используют различные ин -дексы разнообразия, ереди которых наиболее широкое применение получил информацион-ный индекс Шеннона (Н ):
H = ^Ni/N-lg2(Ni/N), где: N- численность i-го вида, N - численность всех видов в сообществе.
Этот индекс суммирует большое количе-ство информации о численности и видовом составе сообществ, ииитывaeт число видов и степень доминирования отдельных видов. Возрастание индекса указывает на увеличе-ние однородности структуры изучаемой си -стемы, снижение - показывает, что структура менее однородна и возрастает доминиро-ванне отдельных элементов. В первом случае структура более сложно организована.
Показано (9), что отношение продукции сообществ животных к тратам на обменные процессы у животных и индекс разнообра-зия, как обобщенная характеристика структурной сложности сообществ, находятся между собой в обратной зависимости (рис. 2):
P/R = е-е " ?н, где: (к, Р-параметры уравнения, к - основание натурального логарифма.
Приведенное уравнение показывает, что по мере усложнения структуры сообществ и экосистем (увеличения их разнообразия) возрастает доля рассеиваемой энергии в виде тепловой по отношению к энергии, заключенной в продукции этих биологических си -стем. Более сложно организованные системы характеризуются большим разнообрази -
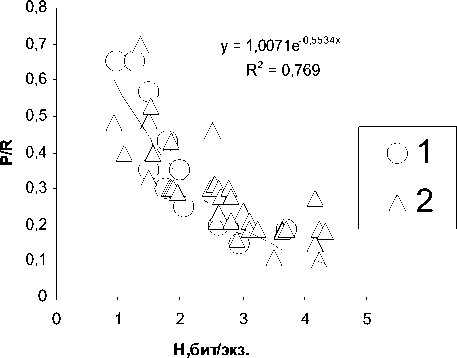
. 2. (P/R)
( ) (1)
ем, меньшей продуктивностью, большей долей диссипируемой энергии по отношению к продуцируемой.
Из сказанного следуют два важных выво-да. Во-первых, структура сообществ организмов и экосистем, с значит, и их разнообразие сохраняется не за счет установления устой -чивых связей между элементами (как у объектов неживой природы), с з с счет постоянных затрат энергии на поддержание упорядочен -ности и воспроизводства элементов систе-мы, их структур и структур организмов. Пр и этом затраты энергии на поддержание струк-туры в тех экосистемах, соторые существуют главным образом за счет притока внешней энергии антропогенного происхождения зна-чительно выше, сем в тех, соторые существуют в основном за счет своих продукционных возможностей. Во-вторых, установленные количественные связи между структурными и функциональными характеристиками био -логических систем дают основание надеять-ся на получение количественных соотноше-ний между потоками энергии и информации в водных экосистемах. Необходимо отметить, что в формировании структуры экосистем важнейшая роль принадлежит информацион-ным связям, сак сегуляторам взаимодействий и связей между популяциями отдельных ви-дов. Каждый перенос энергии и массы в пет лях обратных связей вызывает встречный поток информации. Ин формационные связи и потоки информации в экосистемах много -численны, сложны и практически не исследованы . Однако следует понимать, что без знания организации информационных сетей и потоков информации невозможно разрабо-тать полноценную теорию функционирова-ния экологических систем.
Состояние сообществ гидробионтов и эко -систем может быть описано структурными и функциональными характеристиками. Изменение структуры системы, связанное, например , с исчезновением отдельных видов или появлением новых видов (изменение ее разнообразия ), с сменением трофических связей сриводит к изменению функциональных ха -рактеристик системы, и система переходит в новое состояние, определяемое новыми значениями структурных и функциональных ха -рактеристик. Переход системы из одного состояния в другое обусловлен изменениями ссловий среды, в сом числе и поступлением энергии, и происходит не в один момент, а с сечение некоторого промежутка времени.
Сказанное дает возможность разработки способов расчета ожидаемых усилий при же -лании или необходимости перевода системы в новое состояние и поддержания ее в но -вом состоянии. Например, при желании по- лучить определенный экономический эффект от увеличения продуктивности за счет удоб-рения водоемов, или удаления из них сорных рыб и заселения их ценными видами рыб, при развитии прудового рыбоводства, аквакуль-туры и т. . Однако для достижения постав-ленных целей необходимо проведение ряда мероприятий, направленных на поддержание системы в новом состоянии. Это возможно лишь при условии приложения вполне он-ределенных усилий, внешних по отношению к системе, которые в наиболее общей форме могут быть оценены в виде затрат энергии. Можно ожидать, что для перевода экосисте-мы водоема из эвтрофного состояния в ме-зотрофное должно быть затрачено большее количество энергии, чем из мезотрофного в олиготрофное. Поскольку известно, что в эко-системе при переходе с одного трофического уровня на другой рассеивается около 90% энергии, получение высокой продуктивное-ти сообществ организмов или экосистем воз-можно лишь при упрощении их структуры, т. . сокращении разнообразия.
В результате исследований структуры и функционирования экосистем пресноводных водоемов и их составляющих, изучения эко-логической физиологии водных организмов была разработана теория биологической про-дуктивности водоемов, создана продукцион-ная гидробиология . Активное применение положений продукционной гидробиологии позволило оценить продукционные показа-тели отдельных водоемов, разработать схему потоков энергии в экосистемах водоемов, представить направления возможных их из-менений при разных антропогенных воздей-ствиях .
Проведенные исследования на различных водоемах позволили разработать схему коли-чественного выражения продукционного процесса в водной экосистеме на всех уров-нях в виде потока энергии или преложенно-го Г . . «биотического ба-ланса». Балансовый подход и энергетический принцип стали основополагающими в иссле-дованиях экосистем разных водоемов.
Антропогенные воздействия на экосисте-мы могут приводить либо к их упрощению, что наблюдается, например, при стрессовых на них воздействиях, либо к их усложнению . В последнем случае сложность и разнообра-зие систем возрастают. -сто при восстановлении нарушенных экоси-стем или, например, при разумной акклима-тизации организмов. При антропогенном увеличении сложности экосистем такие вне-шние по отношению к экосистеме воздей-ствия выступают как фактор, влияющий на ход сукцессионного процесса или запускаю-щий его . Так , усиление пресса рыб как резуль-тат увеличения числа их видов или количе-ства рыб в озерах-питомниках изменяет структуру пищевых цепей и продуктивность сообществ донных животных, блокирует ос-новные направления сукцессионного процес-са и возвращает сообщество на ранние ста-дии сукцессии [5].
Активное применение положений продук-ционной гидробиологии позволило оценить продукционные показатели отдельных водо-емов, а также представить направления воз-можных их изменений при загрязнении, ЭВ-трофировании, ацидофикации, эксплуатации водоемов в хозяйственных целях и других антропогенных воздействиях на них.
Развитие продукционной гидробиологии подошло к такому этапу, -посылки к разработке теории функциониро-вания водных экосистем, основные положе-ния которой впервые были изложены авто-ром [3]. Необходимость такой теории пони-мается не только научным сообществом, но и наиболее прогрессивными представителями организаций, ответственных за охрану при-роды , сохранение видового разнообразия, неистощимого использования природных экологических систем.
Методологической основой экологических исследований служит системный подход, ориентированный на раскрытие целостное-ти объекта и обеспечивающих ее механиз-мов , на выявление многообразных связей сложного объекта и сведения их в единую теоретическую картину. ,и экологическая система в том числе, понима-ется как совокупность взаимосвязанных элс-ментов, образующих определенную целост-
,.
-
,
.
,-
, - жет быть охарактеризовано ее структурными
.-
-
-
.-
-
-
. -
-
,
,-
.-
-
,-
-
.
В результате взаимодействия организмов
-
-
, .-
,
,-
, .-
-
-
,
-
[8]. ,,
-
-
.-
-
-
,-
.
Одна из функциональных характеристик
-.- дукции экосистемы по отношению к общим тратам на обменные процессы в экосистеме увеличивалась по мере возрастания в ней
[3]:
P p / R e = 0.360 B e 0.19 -щим тратам на обменные процессы всеми --
(. 1).
Таблица 1. В ел и чины отношения продукции экосистемы и общих трат на обмен в (P/R)e
|
водоем |
Рр, ккал/м2 год |
(P/R)e |
|
. |
1581 |
0.49 |
|
. |
1420 |
0.31 |
|
. |
1256 |
0.2 |
|
. |
265 |
0.32 |
|
. |
857 |
0.52 |
|
. |
2361 |
0.104 |
|
. |
1655 |
0.29 |
|
. |
3064 |
0.08 |
|
: 3 |
992 |
0.21 |
|
5 |
1696 |
0.18 |
|
6 |
3060 |
0.49 |
: ,
. 1,
(P/R)e-
-
0,29, - 0,32.
,
,- системах озер закономерно возрастает по мере
.-
-
-
.
,-
-
- нально увеличению содержания хлорофилла « » [9].
[2], -
-
: = 0.126 ( / 2 ), доля биомассы гетеротрофов в общей биомас -се озерных экосистем снижается по мере воз -растания первичной продукции экосистемы: Вg/ Be = 1.72 рРр - °-338 .
Скорость оборота биомассы в отдельных подсистемах экосистем озер увеличивается с возрастанием первичной продукции в них. Такая же закономерность прослеживается и при эвтрофировании конкретного озера. В то же время скорость оборота биомассы в рас -чете за год в экосистеме в целом остается по -стоянной независимо от величины первич-ной продукции и практически близка к еди-нице [3].
В качестве другой важной функциональ-ной характеристики экосистемы, как и сообществ организмов, может рассматриваться отношение продукции экосистемы к тратам на обмен всеми гидробионтами в нее входя -щими ( Р / R )e. Поскольку для озерных экосистем , это отношение в среднем составляет 0,32, легко рассчитать, что ожидаемая величина коэффициента К2 для экосистемы в целом (Тб2е) Тлизка к 0,24. Это выше, тем средняя величина К 2- коэффициента для сообществ планктонных и донных животных (0,2) т близка к величине этого коэффициента для популяций (0,26). Следовательно, примерно четверть энергии, аесимилированной в экосистеме , тесходуется на создание ее продукции , что позволяет говорить о достаточно высокой эффективности функционирования озерных экосистем, которая может быть оценена величиной К 2- коэффициента.
Таким образом, во-первых, экосистема -более сложно организованная, чем сообщества организмов, в нее входящие, - характеризуется высокой эффективностью функцио-нирования. Во-вторых, екосистему нельзя рассматривать как некую сумму входящих в нее компонентов. Она представляет собой систему с вполне определенными внутрен-ними связями, определяющими ее стабильность в конкретных условиях и может быть охарактеризована такими важнейшими фун-кциональными характеристиками, как скорость оборота биомассы за год (P/В) и эффективность функционирования (К2е), которые не зависят от величин первичной про - тукции экосистем.
В общих процессах круговорота веществ в озерных экосистемах участвуют аллохтонные органические вещества. В экосистемах, т которых (Pe > 0) аллохтонная органика играет незначительную роль в биотическом круто -тороте, скорее всего в большей части, о та поступает в донные отложения или окисля-ется за счет кислорода в воде, т не биотичес-ттм путем. В тех же экосистемах, в которых Pe < 0 аллохтонные органические вещества должны активно вовлекаться в биотический круговорот. При допущении об одинаковом количестве аллохтонных органических ве-ществ в озерах первой и второй группы - в первых заиление должно происходить более интенсивно.
Упрощение структуры сообществ организ-мов или экосистем сопровождается возрас-танием вариабельности динамики биомассы, которую можно рассматривать как показатель устойчивости экосистем [3]. Вариабельность динамики биомассы может быть оценена ве-личиной отношения минимальной за год биомассы к максимальной ее величине за тот же период. Чем больше вариабельность биомассы , ттм более устойчива система к изменениям условий среды, в том числе антропогенным . Вариабельность динамики биомассы возрастает по мере упрощения струк-туры экосистемы, что в первую очередь связано с уменьшением числа видов, сокращением трофических связей, упрощением системы потоков энергии, вещества и информации , т.е. т уменьшением разнообразия системы . Пр и этом доминирование переходит к видам с широкими экологическими спектра -ми.
Отметим, что проблема инвазий новых видов в экосистемы в настоящее время стала актуальной. Для таких экосистем характерно большое видовое разнообразие, наличие ре-ттктовых и эндемичных видов, преобладание стенобионтных видов, значительное превышение процессов деструкции над процес-сами продуцирования. Такие сложно организованные системы оказываются мало вынос -тивыми к различным по качеству и силе ант -ропогенным воздействиям. Экосистемы эво- люционно более старые оказываются более устойчивыми к внешним воздействиям. Для них характерно меньшее видовое разнообра-зие, преобладание эврибионтных видов и преобладание процессов продуцирования.
Увеличение биологического разнообразия экосистем благодаря вселению в водоемы новых видов - это естественный процесс , однако скорость которого возрастает в резуль-тате антропогенных воздействий. Во многих случаях вселение новых видов не оказывает существенного влияния на функционирова-ние экосистем, но в ряде случаев виды-все-ленцы могут существенно изменять функци-ональные характеристики экосистем. При этом вселение нового вида может полностью реконструировать пищевые цепи или приве-сти к созданию новых пищевых цепей. Такая , поскольку может нанести прямой вред его хозяйственной деятельности (как , например , это было при вселении гребневика Mnemiop-sis leidyi A.Agassiz 1865 в Черное море).
Структура сообществ организмов и экоси-стем сохраняется не за счет установления ус-тойчивых связей между элементами, как у объектов неживой природы, а за счет посто-янных затрат энергии на поддержание уно-рядоченности и воспроизводства элементов и структур элементов системы и структур организмов. При этом затраты энергии на поддержание структуры в экосистемах, кото -рые существуют главным образом за счет при-тока внешней энергии, значительно выше, чем в тех , которые могут существовать в ос-новном только за счет своих продукционных возможностей.
Экосистема находится в стабильном состо-янии, пока на нее с постоянной силой воз-действуют конкретные по качеству факторы среды . Она характеризуется конкретными структурными и функциональными характе-ристиками . При изменении силы или каче-ства воздействия на экосистему она перехо-дит в новое состояние, характеризующееся новыми структурными и функциональными показателями . Переход экосистемы из одно-го стабильного состояния в другое стабиль-ное состояние обусловлен изменениями ус- ловий среды, в том числе поступлением энергии, и происходит не мгновенно, а в те-чение некоторого промежутка времени.
Способность к стабилизации представля-ет собой основное свойство экосистемы, ко -торое отличает ее от популяций и сообществ гидробионтов. Это свойство экосистем обус-ловлено динамическим взаимодействием потоков энергии, вещества и информации. Развитие и активное использование резуль-татов продукционной гидробиологии позво-лило установить и количественно выразить основные потоки энергии в экосистемах и построить генеральную схему этих потоков. При этом показана большая роль рыб в ста-билизации потоков энергии, как в сообще-ствах животных, так и в экосистеме в целом. Кроме того, рыбы способствуют поддержа-нию высокого разнообразия в экосистемах. Менее подробно изучены потоки вещества в сообществах гидробионтов и экосистемах. Наиболее подробно изучены потоки биоген-ных элементов , главным образом фосфора и в меньшей степени азота.
Анализ функционирования водных экоси-стем показал, что экосистема представляет собой действительно систему, -ются внутренние механизмы, стабилизирую-щие ее функционирование, как при неизмен-НОЙ, так и при измененной ее структуре .
В формировании структуры экосистем важнейшая роль принадлежит информацион-ным связям . Именно они выступают как ре-гуляторы взаимодействий и связей между популяциями отдельных видов, относящих-ся к одному или разным функциональным блокам . Информация присутствует как в структурах организмов, так и в потоках меж-ду структурами . Каждый перенос энергии и массы в петлях обратных связей вызывает встречный поток информации .
Биологические системы представляют со-бой сложные и упорядоченные объекты, об-ладающие уникальной способностью сохра-нять и передавать информацию в виде струк-тур и функций, которые возникают в резуль-тате длительной эволюции . Синтез веществ и аккумуляция энергоносителей, их трансфор-мация и разрушение любых веществ в живых организмах идут только при участии АТФ и других макроэргов, в том числе нуклеозидов .
В биологии чаще всего передача информа-ции изучается и описывается независимо от вещества и энергии. Но на самом деле есть материальные носители информации, напри-мер , энергия, которая необходима при запи-си и считывании информации . Информация присутствует как в структурах организмов, так и в потоках между структурами.
Информационные связи и потоки инфор-мации в экосистемах многочисленны, слож-ны и крайне слабо исследованы. По крайне мере в водных экосистемах они изучены зна-чительно хуже, чем потоки вещества и тем более энергии. Точнее следует сказать, что исследования информационных связей и по-токов информации в экосистемах находятся лишь в зачаточном состоянии и пока что- то не видно исследователей, нацеленных на та-кие работы. Для изучения и количественно-го выражения информационных связей и потоков информации в водных экосистемах предстоит выполнить огромный объем рабо-ТЫ . Совершенно ясно, что такая задача мо-жет быть решена лишь скоординированны-ми усилиями многочисленных коллективов по единой программе, основу и структуру ко-торой еще предстоит разработать. Но для это-го сначала необходимо достижение понима-ния необходимости работ в этом направле-НИИ . Однако при современном положении в гидробиологической науке пока такое пони-мание к сожалению не прослеживается. Вме-сте с тем без понимания структуры и органи-зации информационных сетей и потоков ин-формации и количественной оценки их взаимосвязей с другими потоками в экосис-темах невозможно представить успешность разработки теории функционирования экоси-стем .
При антропогенном эвтрофировании, не-токсичном загрязнении вод органическими веществами в экосистемах происходят те же процессы, что и при эволюционном разви- тии водоемов, но с более высокими скорое-тями. С ростом продуктивности водоемов в их экосистемах происходит увеличение чис-ла событий и ускоряется ход времени в них.
Фактор времени постоянно присутствует при исследованиях потоков в экосистемах, так как они имеют размерность скорости. Поэтому понятно, что для понимания меха-низмов функционирования экосистем необ-ходим учет и анализ временного фактора. Оценки биологического времени в экосисте-мах разной структуры необходимы, учитывая , что разные организмы существуют в собствен-ном пространстве и времени, но внутри эко-системы структура времени приобретает упо-рядоченность, выражающуюся в подобии времени в размерном спектре [6].
Я не ставил перед собой задачи рассмот-реть все основные результаты, полученные российскими гидробиологами, как и опре-делить задачи и направления дальнейших работ. -ние на наиболее общие и, с моей точки зре-НИЯ, наиболее значимые результаты и следу-ющие из них задачи, без решения которых не вижу возможности решительного движения вперед. Одновременно считаю необходимым подчеркнуть, что именно в нашей стране была разработана продукционная гидробиология, основные положения которой успешно ис-пользовались и используются в теоретичес-ких исследованиях и во многих практичес-ких проблемах, как в нашей стране , так и за ее рубежами. Однако, к сожалению, в после-дние годы наметились опасные тенденции потери или не использования этих знаний, отсутствие стремления движения вперед, ис-чезает любопытство у исследователей, а ра-боты часто становятся все менее обобщаю-щими и все более конъюктурными .
Работа поддержана грантами 05-04-49703, 1634 2003 4, грантом Президиума РАН «Ис-следование современной динамики биораз-нообразияэкосистем».
Список литературы Заметки о современном состоянии гидробиологии континентальных водоемов
- Алимов А. Ф. Функциональная экология пресноводных двустворчатых моллюсков. Л.: Наука, 1981.
- Алимов А.Ф. Исследование биотических балансов экосистем пресноводных водоемов в СССР//Гидробиол. журн. 1987. Т. 23, № 6.
- Алимов А.Ф. Элементы теории функционирования водных экосистем. СПб.: Наука, 2000.
- Алимов А.Ф. Исследование биоразнообразия в сообществах планктона, бентоса, рыб и в экосистемах пресноводных водоемов разной продуктивности//Известия РАН. Сер. биол. 2001. №1.
- Голубков С.М. Динамика пищевых цепей и сукцессия сообществ донных животных в пресных водах//Биол. внутр. вод. 1997. № 1.
- Мусатов А.Л. Пространственно-временная структура водных экосистем. М, 1994.
- Хайлов К.М., Празукин А.В., Минкина Н.И., Павлова Е.В. Концентрация и функциональная активность живого вещества в сгущениях разного уровня организации//Успехи соврем. биол. 1999. Т. 119.
- Bayers R.J., Odum H.T. Ecological microcosms. N.Y., London.1993.
- Connor E.F., McCoy E.D. The statistics and biology of the species -area relationship//Amer. Nat. 1979. Vol. 113.
- Connor E.F., McCoy E.D., Cosby B.J. Model discrimination and expected slope values in species -area studies//Amer. Nat. 1983. Vol. 122.
- Dodson S. Species richness of crustacean zooplankton in European lakes of different sizes//Verh. Intern. Limnol. 1991. Vol. 24.
- Dodson S. Predicting crustacean zooplankton species richness.//Limnol. Oceanogr. 1992. Vol. 37, № 4.
