Зависимость роста побегов и корней, а также содержания и метаболизма цитокининов от чувствительности растений арабидопсиса к этилену

Автор: Васинская Анна Николаевна, Коробова Алла Владимировна, Кудоярова Гюзель Радомесовна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Природопользование
Статья в выпуске: 5-3 т.13, 2011 года.
Бесплатный доступ
В настоящей работе проведено сравнительное изучение роста побегов и корней, а также содержания и метаболизма цитокининов у растений арабидопсиса исходной линии Columbia и их этилен-нечувствительных мутантных растений ETR-1. Показано, что этилен способен снижать содержание цитокининов и тем самым поддерживает накопление биомассы корней, тогда как влияние цитокининов на этот процесс происходит, по-видимому, без вовлечения этилена. Сделан вывод о том, что торможение удлинения корней цитокининами происходит с помощью этиленового сигналинга.
Этилен, цитокинины, рост корней
Короткий адрес: https://sciup.org/148200427
IDR: 148200427 | УДК: 581.1
Dependence of root and shoot growth, contentand metabolism of cytokinins from ethylene sensibility of arabidopsis plants
A comparative study of shoot and root growth, cytokinin content and metabolism of wild type Arabidopsis plants (Columbia) and their ethylene insensitive mutants (ETR-1) have been carried out. Ethylene was shown to be able to decrease cytokinin content and therefore to maintain root biomass accumulation. On the contrary, cytokinins influence on this process apparently without involvement of ethylene. The involvement of ethylene signaling in inhibition of root elongation by cytokinins is concluded.
Текст научной статьи Зависимость роста побегов и корней, а также содержания и метаболизма цитокининов от чувствительности растений арабидопсиса к этилену
камере при освещенности 120 μмоль фотонов/(м2 с), температуре 23оС днем и 19оС ночью, продолжительности светового периода 16 ч.
После прорастания семян относительную влажность песка поддерживали на уровне 65%.
В возрасте 28 сут у растений определяли сырую массу побегов и корней и фиксировали их в 80%-ном этаноле для экстракции цитокининов.
Экстракцию, очистку и концентрирование цитокининов проводили как описано ранее [11]. Концентрацию цитокининов в побегах и корнях растений определяли с помощью твердофазного имму-ноферментного анализа, используя антитела к рибозиду зеатина, имеющие также высокую кроссреактивность к зеатину и зеатиннуклеотиду [12].
Для определения активности цитокининоксида-зы использовали метод деградации изопентенила-денозина (ИПА) в присутствии белкового экстракта из растительной ткани.
Растительный материал гомогенизировали в холодном (6°С) 0.1М имидазол(HCl-буфере (pH 7.1). Гомогенат центрифугировали при 12 000 g в течение 25 мин. Белки из супернатанта осаждали добавлением равного объема насыщенного раствора (NH 4 ) 2 SO 4 и центрифугировали при 5000 g в течение 15 мин. Осадок ресуспензировали в имидазол-HCl-буфере (pH 7.1). Далее 50 мкл препарата добавляли к 50 мкл смеси, содержавшей стандарт ИПА и имидазол(HCl-буфер (pH 7.1) в конечной концентрации 0.04(0.06 М. Для контроля в смесь вместо белкового препарата добавляли 50 мкл соответствующего буфера.
Реакционные смеси инкубировали в термостате при 37°С в течение 3 ч. Реакцию останавливали добавлением 0.2 мл холодного этанола, и смесь центрифугирование при 5000 g в течение 10 мин. Содержание ИПА в супернатанте оценивали с помощью иммуноферментного анализа в тест-системе для определения изопентениладенози-на/изо-пентениладенина [13].
Статистическую обработку проводили по стандартным программам MS Excel.
На рисунках и в тексте представлены средние значения и стандартная ошибка показателей из трех и более биологических повторов (n).
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Известно, что этилен является ингибитором роста корней в длину [10, 14].
Это подтверждается в нашем опыте: корни нечувствительных к этилену мутантных растений были длиннее, чем у растений исходной линии (табл.).
Масса корней у мутантных растений была достоверно меньше, чем у растений исходной
Таблица. Длина корней, масса сырого вещества побегов и корней, соотношение массы побег/корень 28дневных растений арабидопсиса ( Arabidopsis thaliana ) дикого типа (COL) и их этиленнечувствительных мутантов (ETR-1), n = 30
Потеря чувствительности растений к этилену не влияла на накопление массы побегов (табл.). Таким образом, у нечувствительных к этилену растений наблюдалось увеличение соотношения массы побега и корня, что свидетельствует об относительной активации роста побега по сравнению с корнями у мутантных растений в сравнении с исходными (табл.).
Такая реакция является характерным ответом растений на цитокинины [15, 16]. Важно то, что по сравнению с растениями исходного генотипа у мутантных растений мы действительно наблюдали повышение содержания цитокининов, особенно ярко выраженное в их корнях (рис. 1).
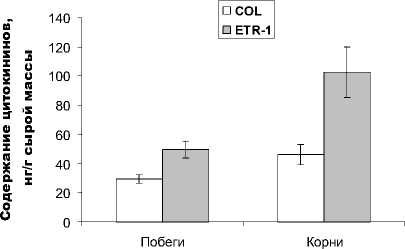
Рис. 1. Содержание цитокининов в побегах и корнях 28-дневных растений арабидопсиса дикого типа (COL) и их этиленнечувствительных мутантов (ETR-1), n = 3
В литературе есть сведения о способности этилена активировать распад цитокининов [6]. Поэтому неудивительно, что мы обнаружили повышенное содержание цитокининов у растений, нечувствительных к этилену. Известно, что избыточные концентрации цитокининов подавляют рост и раз- витие корневой системы [17]. Можно было предполагать, что этилен способствует накоплению биомассы корней растений через нормализацию уровня цитокининов и предотвращение их чрезмерного накопления.
Известно, что распад цитокининов катализирует фермент цитокининоксидаза [13, 18] Мы оценили активность этого фермента у растений двух генотипов. В наших экспериментах активность цитоки-ниноксидазы у нечувствительных к этилену растений также была выше, чем у растений исходного типа (рис. 2). Поэтому более высокое содержание цитокининов у растений потерявших чувствительность к этилену не могло быть следствием изменения активности цитокининоксидазы. Скорее оно могло быть его причиной. По литературным данным активность цитокининоксидазы может повышаться вслед за увеличением концентрации эндогенных цитокининов [18]. Следовательно, и в наших экспериментах происходила субстратная регуляция активности фермента. Снижение уровня цитокининов под влиянием этилена могло быть связано с необратимой конъюгацией цитокининов или ингибированием их синтеза.
Наши результаты помогают лучше понять механизм действия цитокининов на рост корней. По данным литературы, эти гормоны тормозят как накопление биомассы, так и удлинение корней [17]. Однако в наших опытах у мутантных растений, потерявших чувствительность к этилену, накопление цитокининов в корнях не сопровождалось торможением их удлинения. Наоборот, они были длиннее, чем у растений исходной линии. Следовательно, наши результаты подтверждают, что способность цитокининов подавлять удлинение корней осуществляется через этилензависимый каскад [19]. Вместе с тем, цитокинины подавляли накопление биомассы корней у нечувствительных к этилену мутантов. Эти результаты указывают на то, что это свойство цитокининов является этилен независимым.
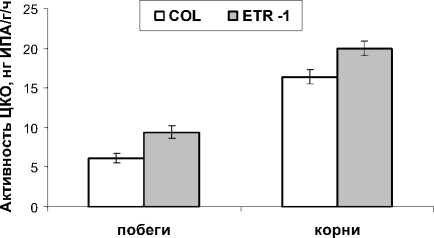
Рис. 2. Активность цитокининоксидазы в побегах и корнях 28-дневных растений арабидопсиса дикого типа (COL) и их этиленнечувствительных мутантов (ETR-1), n = 3
Таким образом, этилен способен ингибировать удлинение корней, однако он, по-видимому, необходим для поддержания накопления массы корней. Это может быть связано с его способностью ограничивать накопление цитокининов в растениях. С другой стороны, снижение прироста массы корней у нечувствительных к этилену растений на фоне повышенного содержания эндогенных цитокининов может говорить о том, что ингибирующее влияние цитокининов на этот процесс, по-видимому, не связано с этиленовым сигналингом.
Список литературы Зависимость роста побегов и корней, а также содержания и метаболизма цитокининов от чувствительности растений арабидопсиса к этилену
- Кулаева О.Н. Цитокинины, их структура и функции. М.: Наука, 1973. 264 с
- Haberer, Kieber. Cytokinins. New Insights into a Classic Phytohormone//Plant Physiology. 2002. V. 128. Р. 354-362
- Mok D.W.S., Mok M.C. Cytokinin metabolism and action//Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001. V. 52. P. 89-118.
- Kurakawa T., Ueda N., Maekawa M., Kobayashi K., Kojima M., Nagato Y., Sakakibara H., Kyozuka J. Direct control of shoot meristem activity by a cytokinin-activating enzyme//Nature. 2007. V. 445. P. 652-655.
- Cary A.J., Liu W., Howell S.H. Cytokinin action is coupled to ethylene in its effects on the inhibition of root and hypocotyl elongation in Arabidopsis thaliana seedlings//Plant Physiology. 1995. V. 107. P. 1075-1082.
- Taverner E., Letham D.S., Wang J. et al. Influence of ethylene on cytokinin metabolism in relation to Petunia corolla senescence//Phytochemistry. 1999. V. 51. P. 341-347.
- Pierik R., Sasidharan R., Voesenek L.A.C.J. Growth control by ethylene: adjusting phenotypes to the environment//J. Plant Growth Regulation. 2007. V. 26. P. 188-200.
- Vandenbussche F., van der Straeten D. One for all and all for one: cross-talk of multiple signals controlling the plant phenotype//J. Plant Growth Regulation. 2007. V. 26. P. 178-187.
- Ракитин В.Ю., Прудникова О.Н., Карягин В.В. и др. Выделение этилена, содержание АБК и полиаминов в Arabidopsis thaliana при УВ-В стрессе//Физиология растений. 2008. Т. 55. № 3. С. 355-361.
- Stepanova A.N., Hoyt J.M., Hamilton A.A., Alonso J.M. A link between ethylene and auxin uncovered by the characterization of two root-specific ethylene-insensitive mutants in Arabidopsis//Plant Cell. 2005. V. 17. P. 2230-2242.
- Kudoyarova G.R., Vysotskaya L.B., Cherkozyanova A., Dodd I.C. Effect of Partial Rootzone Drying on the Concentration of Zeatintype Cytokinins in Tomato (Solanum lycopersicum L.) Xylem Sap and Leaves//J. Exp. Bot. 2007. V. 58. P. 161-168.
- Vysotskaya L.B., Kudoyarova G.R., Veselov S., Jones H.G. Unusual Stomatal Behaviour on Partial Root Excision in Wheat Seedlings//Plant Cell Environ. 2004. V. 27. P. 69-77.
- Веселов С.Ю., Симонян М.В. Использование иммуноферментного анализа цитокининов для оценки активности цитокининоксидазы//Физиология растений. 2004. Т. 51. С. 297-302.
- Achard P., Vriezen W.H., van der Straeten D., Harberd N.P. Ethylene regulates Arabidopsis development via the modulation of DELLA protein growth repressor function//Plant Cell. 2003. V. 15. P. 2816-2825.
- Arkhipova T.N., Prinsen E., Veselov S.U., Martinenko E.V., Melentiev A.I., Kudoyarova G.R. Cytokinin producing bacteria enhance plant growth in drying soil//Plant and Soil. 2007. V. 292. № 1-2. P. 305-315.
- Vysotskaya L.B., Korobova A.V., Veselov S.Yu., Dodd I.C., Ku-doyarova G.R. ABA mediation of shoot cytokinin oxidase activity: assessing its impacts on cytokinin status and biomass allocation of nutrient deprived durum wheat//Funct. Plant Biol. 2009. V. 36. № 1. P. 66-72.
- Kuderova A., Urbankova I., Valkova M. et al. Effects of conditional IPTdependent cytokinin overproduction on root architecture of Arabidopsis seedlings//Plant and Cell Physiology. 2008. V. 49. P. 570-582.
- Brugiere N., Jiao S., Hantke S. et al. Cytokinin Oxidase Gene Expression in Maize Is Localized to the Vasculature, and Is Induced by Cytokinins, Abscisic Acid, and Abiotic Stress//Plant Physiology. 2003. V. 132.P. 1228-1240.
- Ruzicka K., Simaskova M., Duclercq J. et al. Cytokinin regulates root meristem activity via modulation of the polar auxin transport//Proc. Natl. Acad. Sci. USA. 2009. V. 106. P. 4284-4289.
