Жизненные формы и архитектурные модули болотных видов ив бореальной зоны Евразии

Автор: Недосеко О.И., Костина М.В.
Журнал: Фиторазнообразие Восточной Европы @phytodiveuro
Статья в выпуске: 1 т.20, 2026 года.
Бесплатный доступ
На примере четырех видов болотных ив выделены четыре жизненные формы. У двух видов в зависимости от экологических условий проявляется морфологическая поливариантность развития, приводящая к формированию во взрослом состоянии нескольких жизненных форм: у Salix rosmarinifolia формируется две жизненные формы – эпигеогенно-геоксильный (до 2.5 м) и гипогеогенно-геоксильный (до 3.2 м) кустарники, у S. aurita формируется три жизненные формы – эпигеогенно-геоксильный (до 3.4 м) и гипогеогенно-геоксильный (до 4 м) кустарники и жизненная форма деревце (до 1.5 м). У S. lapponum и S. myrtilloides в связи с однотипными условиями произрастания во взрослом состоянии образуется одна жизненная форма – низкий длинноксилоризомный гипогеогенно-геоксильный кустарник (до 1.7 м). Архитектурный модуль ив – это трехлетняя побеговая система (ТПС), основанная на трех признаках: варианте ветвления, размере зоны отмирания вегетативных побегов, долговечности вегетативных частей генеративных побегов. С учетом этих признаков у изученных видов выделено три архитектурных модуля.
Бореальные виды ив, болотные виды, жизненные формы, архитектурные модули
Короткий адрес: https://sciup.org/148332994
IDR: 148332994 | УДК: 581.412 | DOI: 10.24412/2072-8816-2026-20-1-33-45
Life forms and architectural modules of swamp willow species in boreal zone of Eurasia
Using the example of four types of swamp willows, 4 life forms and three architectural modules are distinguished. Depending on the environmental conditions, morphological polyvariety of the development is manifested in two species, leading to the formation of several life forms in adulthood: in Salix rosmarinifolia, two life forms are formed – epigeogenic-geoxylic and hypogeogenic-geoxylic shrubs, in S. aurita, three life forms are formed – epigeogenic-geoxylic and hypogeogenic-geoxylic shrubs and a life form of a tree. In S. lapponum and S. myrtilloides, due to the same type of growing conditions, one life form is formed in the adult state – a low long-xylorizome hypogeogenic-geoxyl shrub. The willow architectural module is a three-year shoot system (TSS) based on three traits: the branching variant, the size of the vegetative shoot dieback zone, and the longevity of the vegetative parts of generative shoots. Based on these features, three architectural modules have been identified in the studied species.
Текст научной статьи Жизненные формы и архитектурные модули болотных видов ив бореальной зоны Евразии
С современной точки зрения для развития фундаментальных основ биоморфологии и популяционно-онтогенетического направления необходимо целенаправленно изучать разнообразие биоморф, онтогенеза и популяционной организации с позиций структурного и биологического многообразия.
Решение этих задач невозможно без изучения становления жизненных форм и архитектурных моделей растений конкретных систематических групп в разных частях их ареалов. Особый интерес представляют таксоны с большим разнообразием жизненных форм. К таковым на территории России относится род Salix L. По разным данным он включает 300–450 видов, широко представленных в разных растительных сообществах (Skvortzov, 1968, 1999; Argus, 1997). В средней полосе европейской России растёт 40 видов ив (Antsiferov, 1984; Valyagina-Malyutina, 2004), которые отнесены к трем подродам: Salix , Vetrix Dum. и Chamaetia (Dum.) Nas. Широкое распространение представителей рода Salix коррелирует с разнообразием их жизненных форм (Mazurenko, 2010), что обусловлено высокой пластичностью к ведущим факторам среды (освещение, затопление, степень аэрации и сухость субстрата).
Анализируя литературу можно заключить, что в арктических и гипоарктических районах Северо-Востока бывшего СССР среди ив наиболее часто встречается жизненная форма стланикового стержнекорневого кустарничка с надземными ветвями и древесными эпигеогенными корневищами (ксилоризомами) (Derviz-Sokolova, 1982а, b); в условиях Колымского нагорья у ив подрода Chamaetia преобладают подушковидные жизненные формы (Mazurenko, 2007); среди ив Южного Урала наиболее часто встречается жизненная форма кустарник (Getmanets, 2011).
Жизненные формы бореальных видов ив различных фитоценозов, в том числе и болотная группа, недостаточно изучены.
В последнее время в морфологии растений усилилось изучение надземных и подземных органов древесных растений на основе концепции «архитектурных моделей», предложенной и развиваемой F. Halle с соавторами (Halle, Oldeman, 1970; Halle, 1975; Tomlinson, 1978). Данная концепция основывается на изучении прежде всего деревьев экваториального и тропического бессезонного климата. Сопоставление структуры побеговых систем 15 видов лесообразующих деревьев умеренной зоны с архитектурными моделями, показало, что ни по одной модели полного соответствия не наблюдается, что обусловлено несовпадением признаков, которые нуждаются в дополнении или в уточнении применительно к условиям сезонного климата (Kostina et al., 2024). Нами предложена методика изучения архитектурных модулей на примере видов рода Salix , основанная на трех признаках – долговечность вегетативных частей генеративных побегов, вариант ветвления и размер зоны отмирания вегетативных побегов (число отмирающих верхних метамеров) (Nedoseko, 2018).
Цель исследования: на примере болотных видов ив выявить разнообразие жизненных форм и их архитектонику .
М АТЕРИАЛЫ И МЕТОДЫ
Полевые исследования проводились на территории Владимирской, Московской и Нижегородской областей. Районы исследований относятся к Валдайско-Онежской подпровинции Североевропейской таежной провинции Евразиатской таежной (хвойно-лесной) области и характеризуются умеренно-континентальным климатом (Rastitelnost…, 1980).
Объекты исследования – бореальные виды ив, произрастающие в условиях осоковых и сфагновых болот, заболоченных лугов. В настоящей работе мы используем классификацию рода Salix, предложенную А.К. Скворцовым (Skvortzov, 1968, 1999). Название видов приведено в соответствии с принципом приоритета МКБН; латинские названия сверены с данными международного сайта International Plant Names Index; название секций представлено в соответствии с A. K. Skvortsov (1999).
Нами исследовано 4 вида из двух подродов Vetrix и Chamaetia . К подроду Vetrix относятся 3 вида: S. aurita L. из секции Vetrix Dumort., S. lapponum L . из секции Villosae Rouy. , S. rosmarinifolia L . из секции Incubaceae A. Kern. К подроду Chamaetia – 1 вид S. myrtilloides L. (секция Myrtilloides Koehne).
При определении жизненных форм использовалась классификация лесных кустарников, разработанная И.И. Истоминой и Н.Н. Богомоловой (Istomina, Bogomolova, 1991).
Для выделения и характеристики онтогенетических состояний использовалась классификация Т.А. Работнова (Rabotnov, 1950) и А.А. Уранова (Uranov, 1975). Определение онтогенетических состояний деревьев и кустарников проведено по методике, разработанной и апробированной многими авторами (Smirnova et al., 1999; Evstigneev, Korotkov, 2016 и др.).
Для изучения архитектурных модулей применялась авторская методика изучения трехлетних побеговых систем (ТПС) молодых генеративных особей рода Salix (Nedoseko, 2018).
По классификации А.К. Скворцова (Skvortzov, 1968) все изученные виды – неаллювиальные, то есть могут селиться на разнообразном субстрате, в том числе на глинистом, торфяном или покрытом мхами, песчаном; они менее требовательны к аэрации субстрата и часто мирятся с застойным увлажнением или же вообще довольствуются умеренным увлажнением обычных лесных или луговых почв. S. aurita произрастает на мезотрофных окраинах болот, в светлых березовых лесах; в увлажненных вторичных местообитаниях, на кислых, бедных почвах. S. lapponum предпочитает эвтрофные и переходные болота, сырые и заболоченные луга и лесные прогалины, заболоченные леса.S. rosmarinifolia встречается на торфянистых лугах, эвтрофных и переходных болотах (Skvortzov, 1968).
S. myrtilloides предпочитает мезотрофные болота, осоково-кустарниковые и вейниковые переходные зоны по краям сфагновых болот, сырые луга, ерники, сырые и заболоченные сосняки и лиственничники; реже выходит на чистые сфагновые болота, так же редко в горных тундрах.
Р ЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Жизненные формы Salix rosmarinifolia
В литературных источниках (Skvortzov, 1968; Getmanets, 1994; Valyagina-Malyutina, 2004) жизненная форма S. rosmarinifolia определяется как низкий кустарник до 1.5-2.5 м с подземным стеблем и многочисленными, тонкими ветвями, направленными вверх. Нами установлено, что во взрослом состоянии ива розмаринолистная может формировать жизненные формы эпигеогенно-геоксильный и гипогеогенно-геоксильный кустарники (рис. 1). Растения таких жизненных форм растут в разных экологических условиях, у них разное происхождение стволиков и их различные количественные характеристики.
Онтогенез S. rosmarinifolia жизненной формы эпигеогенно-геоксильный изучен на модельных экземплярах, которые растут на заболоченных лугах (рис. 1 А). Онтогенез S. rosmarinifolia жизненной формы гипогеогенно-геоксильный кустарник изучен на особях, которые растут по окраинам осоковых и торфяных болот (рис. 1 Б).
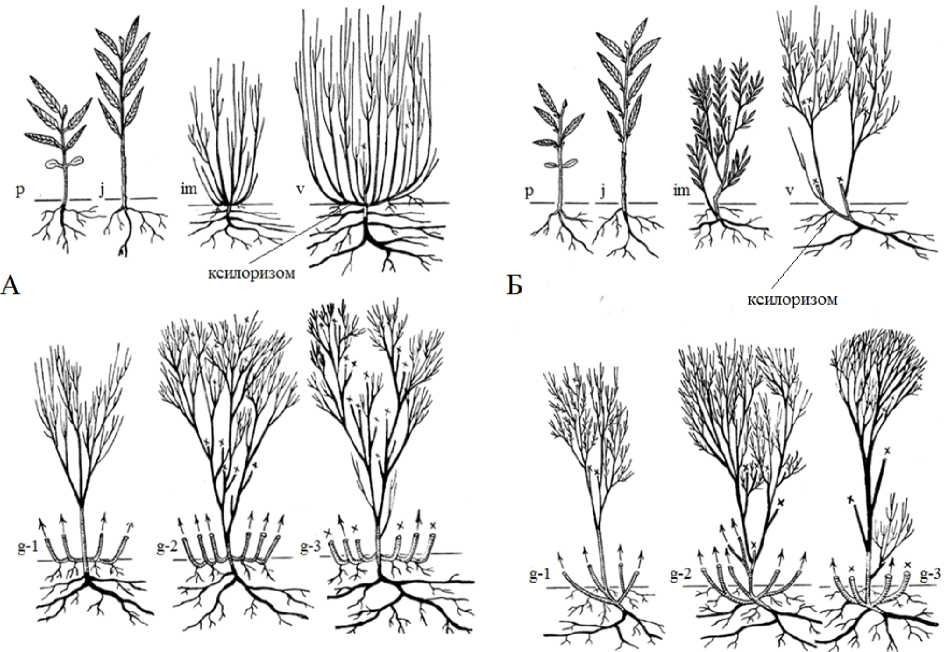
Рис. 1. Схема онтогенеза особей Salix rosmarinifolia жизненной формы эпигеогенно-геоксильный кустарник (А), гипогеогенно-геоксильный кустарник (Б)
Условные обозначения: стрелочками показано направление роста скелетных осей, крестиками – отмершие побеги
Fig. 1. Scheme of ontogenesis of Salix rosmarinifolia individuals of the epigeogenic-geoxyl shrub (A), hypogeogenic-geoxyl shrub (B)
Symbols: arrows show the direction of growth of skeletal axes, crosses – dead shoots
Первые этапы онтогенеза (развитие проростка, ювенильного растения) протекают так же, как и у эпигеогенно-геоксильного кустарника (рис. 1). Основные изменения наступают в имматурном и виргинильном состояниях.
Взрослые растения S. rosmarinifolia жизненных форм эпигеогенно-геоксильный и гипогеогенно-геоксильный кустарник формируются в результате развития побегов формирования из спящих почек базальной части имматурных или виргинильных растений (рис. 1).
Побеги формирования усиленно растут и ветвятся и к 3–5 году жизни становятся стволиками. У проростка спящими становятся почки, находящиеся в пазухах семядольных листьев. У растений последующих онтогенетических состояний спящими на годичных побегах будут мелкие почки, образующиеся в пазухах почечных чешуй, а также одна или две почки в пазухах нижних фотосинтезирующих листьев. С возрастом увеличивается число спящих почек. В результате развития побегов формирования из надземно расположенных спящих почек на саблевидно-изогнутых побегах базальной части растений, формируется жизненная форма эпигеогенно-геоксильный кустарник (рис. 1). Она представлена системой стволиков с длительностью жизни до 20 лет, которые сменяют друг друга в ходе онтогенеза и каждый из них образует систему придаточных стеблевых корней. Взрослые особи S. rosmarinifolia этой жизненной формы вырастают до 1.7–2.26 м, для них характерно компактное расположение многочисленных стволиков (до 318 скелетных осей) с диаметром у основания 1.5–2 см. Диаметр общей кроны куста достигает 8 м. В результате развития побегов формирования из подземно расположенных спящих почек короткого ксилоризома образуется жизненная форма гипогеогенно-геоксильный кустарник (рис. 1). Такие кустарники вырастают до 3.2 м, у них небольшое число стволиков (9–42), образующих систему придаточных стеблевых корней, продолжительность жизни их до 25 лет.
Жизненные формы Salix aurita
В литературных источниках жизненная форма S. aurita определяется как невысокий кустарник высотой 1–3 м (Skvortzov, 1968; Valyagina-Malyutina, 2004). Нами установлено, что во взрослом состоянии ива ушастая может формировать жизненную форму геоксильный кустарник в двух модификациях – эпигеогенно-геоксильный и гипогеогенно-геоксильный (рис. 2) и жизненную форму деревце (рис. 3). Растения этих жизненных формы обитают в различных экологических условиях, у них разное происхождение основных скелетных осей и разные их количественные характеристики.
Онтогенез растений жизненной формы эпигеогенно-геоксильный кустарник S. aurita изучен на особях, растущих на открытых участках заболоченного луга, по окраинам осоковых болот (рис. 2 А).
Растения жизненной формы гипогеогенно-геоксильный кустарник встречаются на окраинах осоковых болот в березовом лесу при наличии верхнего яруса древесной растительности (рис. 2 Б).
Первые этапы онтогенеза (проросток, ювенильное состояние) протекают так же как и у эпигеогенно-геоксильных кустов (рис. 2). Основные различия появляются в im- и v-онтогенетических состояниях. Как и у S. rosmarinifolia. , жизненные формы эпигеогенно-геоксильный и гипогеогенно-геоксильный кустарники S. aurita образуются в результате развития побегов формирования из спящих почек нижней части осей имматурных или виргинильных растений (рис. 2). К 3–5 году жизни побеги формирования становятся стволиками. Еще у проростка часть почек становится спящими – это почки, которые находятся в пазухах семядолей. У остальных растений число спящих почек увеличивается за счет мелких почек, которые формируются в пазухах почечных чешуй и нижних фотосинтезирующих листьев. В течение онтогенеза отмечено ветвление спящих почек, что приводит к увеличению их численности.
Жизненная форма эпигеогенно-геоксильный кустарник образуется при надземном развитии побегов формирования из спящих почек, расположенных на саблевидно-изогнутых базальных частях стволиков (рис. 2 А), состоит из сменяющих друг друга стволиков с продолжительностью жизни до 25 лет, которые образуют каждый раз свою систему придаточных стеблеродных корней. Такие особи вырастают до 2.2–3.4 м, содержат до 60 компактно расположенных стволиков, их диаметр 3.0–6.8 см; диаметр общей кроны куста до 3.5–4.2 м.
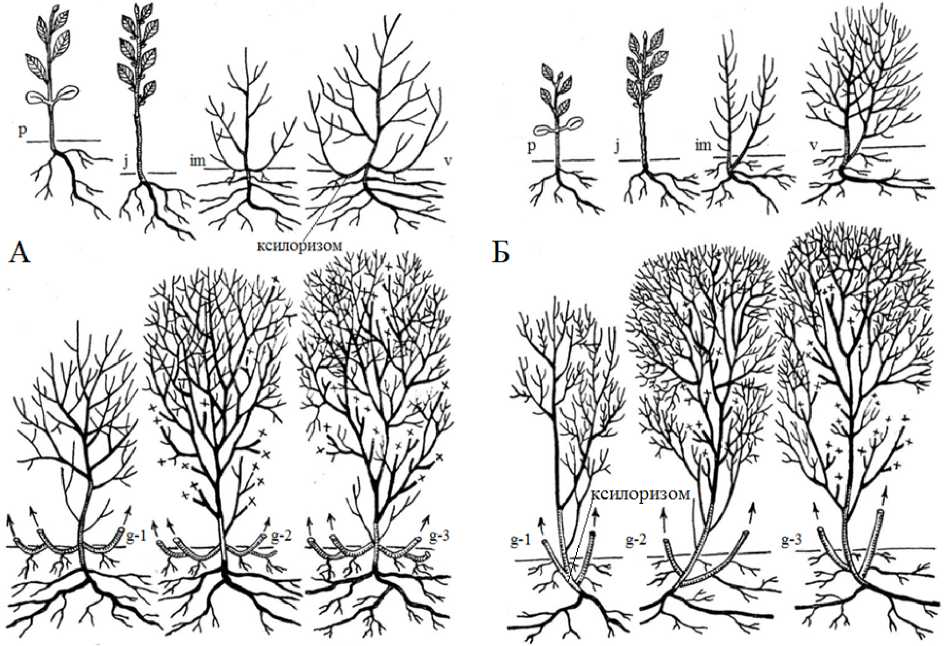
Рис. 2. Схема онтогенеза особей Salix aurita жизненной формы эпигеогенно-геоксильный кустарник (А), гипогеогенно-геоксильный кустарник (Б)
Условные обозначения: стрелочками показано направление роста скелетных осей, крестиками – отмершие побеги
Fig. 2. Scheme of ontogenesis of Salix aurita individuals of the epigeogenic-geoxyl shrub (A), hypogeogenic-geoxyl shrub (B)
Symbols: arrows show the direction of growth of skeletal axes, crosses – dead shoots
Жизненная форма гипогеогенно-геоксильный кустарник S. aurita формируется при развитии побегов формирования из подземно расположенных спящих почек ксилоризома (рис. 2 Б).
Особи этой жизненной формы вырастают до 4.0 м, у них до 2–4 стволиков, диаметр ствола 6–9
см; диаметр общей кроны куста до 1.8–3.2 м.
Жизненная форма деревце S. aurita встречена один раз на возвышении в сухом участке березового леса вблизи травяного болота (вблизи д. Пиявочное Арзамасского района) (рис. 3).
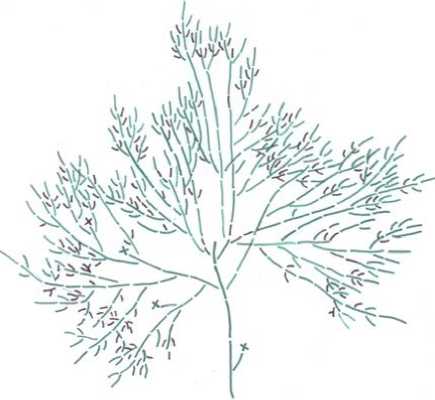
Рис. 3. Схема строения основной скелетной оси Salix aurita (жизненная форма деревце) в средневозрастном генеративном онтогенетическом состоянии
Условные обозначения: зеленым цветом обозначены побеги средней длины, черным – короткие побеги
Fig. 3. Structural diagram of the main skeletal axis of Salix aurita (life form of a tree) in a middleaged generative ontogenetic state
Symbols: green indicates shoots of medium length, black indicates short shoots
Растение данной жизненной формы находится в средневозрастном генеративном состоянии. Его высота – 1.55 м, возраст – 20 лет, его диаметр – 3.2 см, крона расположена на высоте 24 см, при этом ее диаметр 1.4 м. Порядок ветвления побегов – 8–9, длина годичных приростов побегов нарастания – 16–25 см. Из спящей почки подземно расположенного ствола сформировалась 8-летняя поросль, достигающая высоты 54 см.
Система главного корня проникает до глубины 10–15 см. Длинные поверхностные придаточные корни диаметром 1–2.5 см и длиной до 4.5 м направлены в разные стороны и достигают воды в болотце, их длина до 4.5 м, а диаметр до1–2.5 см.
Жизненные формы Salix myrtilloides и S. lapponum
По литературным данным жизненная форма S. myrtilloides определяется как низкий болотный кустарник высотой до 0.2–0.8 (1.3) м (Valyagina-Malyutina, 2004), 0.8–1.5 м (Mazurenko, 2008). В ходе наших исследований установлено, что во взрослом состоянии S. myrtilloides может иметь жизненную форму низкий длинноксилоризомный гипогеогенно-геоксильный кустарник высотой до 1 м (рис. 4 А).
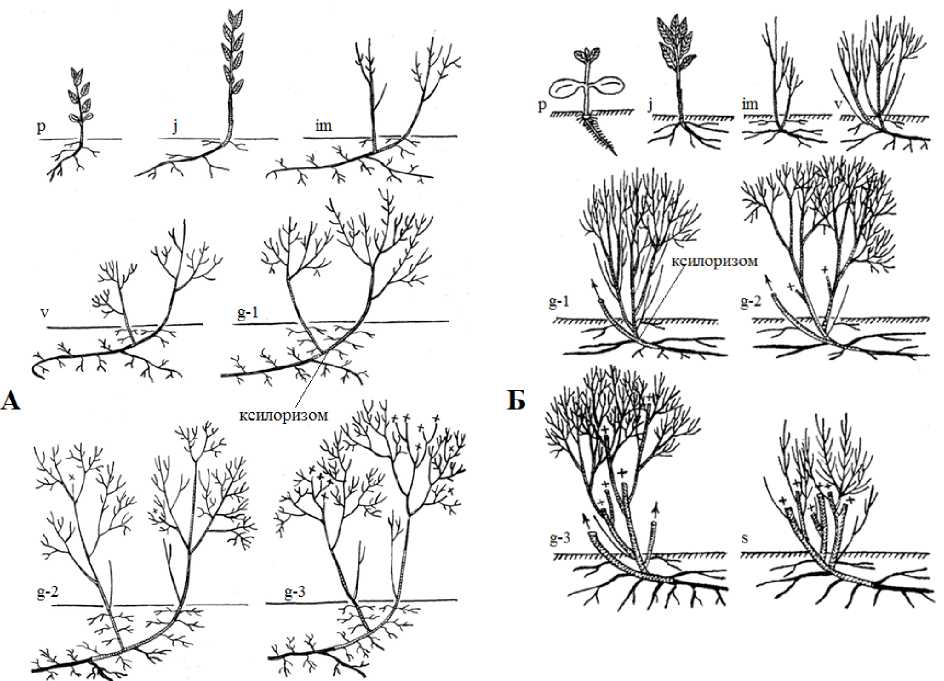
Рис. 4. Схема онтоморфогенеза особей Salix myrtilloides (А) и S. lapponum (Б) жизненной формы низкий длинноксилоризомный гипогеогенно-геоксильный кустарник
Условные обозначения: стрелочками показано направление роста скелетных осей, крестиками – отмершие побеги
Fig. 4. Scheme of ontomorphogenesis of Salix myrtilloides (A) and S. lapponum (B) individuals of the life form of a low long-xylorizome hypogeogenic-geoxyl shrub
Symbols: arrows indicate the direction of growth of skeletal axes, crosses indicate dead shoots.
Онтогенез особей S. myrtilloides данной жизненной формы был изучен на модельных экземплярах, произрастающих на окраинах осоковых и сфагновых болот (на сфагновых и осоковых кочках) при наличии верхнего яруса травяной и древесной растительности. В составе данного фитоценоза произрастают единичные экземпляры березы белой, сосны обыкновенной, ели европейской, ивы пепельной, ивы ушастой, ивы приземистой, голубики, много осоки, мха сфагнума и клюквы.
S. lapponum растет на эвтрофных и переходных болотах, заболоченных лугах. В Нижегородской области ива лапландская встречается спорадически, небольшими малочисленными группами особей. Численность имеет тенденцию к снижению (Krasnaya…, 2005). В Нижегородской области вид отмечен на территории Лысковского, Тоншаевского, Ветлужского, Шахунского, Краснобаковского, Воскресенского, Семеновского, Борского, Выксунского, Кулебакского, Балахнинского, Володарского, Арзамасского, Воротынского районов (Krasnaya…, 2005). Местообитания ивы лапландской в Нижегородской области охраняются на территории государственного природно-биосферного заповедника «Керженский», Пижемского комплексного заказника, Пустынского охотничьего заказника и 8 государственных памятников природы (болота и озера) (Krasnaya…, 2005).
В Арзамасском районе S. lapponum произрастает на болоте Козьем и болоте Мостовом, которые находятся на территории Пустынского заказника. Нами S. lapponum обнаружена на болоте около села Пошатово Арзамасского района – это новое местонахождение S. lapponum, не указанное ранее в литературных источниках. Здесь совместно с ивой лапландской произрастают береза бородавчатая, ива пятитычинковая, ива ушастая, ива пепельная, в травянистом ярусе – виды осок, сабельник болотный, шлемник обыкновенный, лапчатка прямостоячая, мох сфагнум.
По литературным данным жизненная форма S. lapponum определяется как низкий кустарник, высотой 1–1.5 м (Skvortzov, 1968; Gubanov et al., 2003). Нами жизненная форма S. lapponum определяется как низкий длинноксилоризомный гипогеогенно-геоксильный кустарник, высотой до 1.5 м, с несколькими скелетными осями (5–7), диаметром стволиков до 2 см (рис. 4 Б).
Таким образом, в ходе онтогенеза у S. lapponum , как и у S. myrtilloides , во взрослом состоянии образуется жизненная форма низкий длинноксилоризомный гипогеогенно-геоксильный кустарник. Данная жизненная форма образуется в результате развития стволиков (основных скелетных осей) из подземно расположенных спящих почек ксилоризома (рис. 4). Низкий длинноксилоризомный гипогеогенно-геоксильный куст S. lapponum состоит из небольшого числа основных скелетных осей (до 4) с длительностью жизни до 13 лет, образующих систему придаточных стеблеродных корней.
Особи данной жизненной формы наиболее часто произрастают на окраинах осоковых и сфагновых болот, т.е. в местообитаниях, покрытых сфагновыми мхами. В ходе проведенных исследований у S. lapponum и S. myrtilloides было выделено 2 группы особей по уровню жизненности: 1) особи нормальной жизненности и 2) особи пониженной жизненности (размерная поливариантность). Данные особи могут встречаться в пределах одного растительного сообщества, однако особи нормальной жизненности тяготеют к более разреженным участкам подобных ценозов, характеризующихся значительно меньшей сомкнутостью крон (в основном на свободных от растительности или с редкой растительностью сфагновых кочках), а особи пониженной жизненности встречаются на более затененных участках.
Архитектоника изученных видов ив
В качестве основной структурной единицы ив мы рассматриваем трехлетнюю побеговую систему (ТПС), которая состоит из трехлетнего, двухлетнего (одного или нескольких) и нескольких однолетних побегов последнего года вегетации (Nedoseko, 2018). Архитектурный модуль ив – это трехлетняя побеговая система (ТПС), основанная на трех признаках: варианте ветвления, размере зоны отмирания вегетативных побегов, долговечности вегетативных частей генеративных побегов.
Структура ТПС зависит от варианта ветвления: у изученных видов кустарников – мезо- и базитония. Сам вариант ветвления коррелирует с числом отмирающих верхних метамеров: у жизненных форм высотой 2.5–4.0 м на побеге отмирает до четырех, у жизненных форм, высотой 1.0–1.7 м – до половины метамеров побега.
Кроме того, структура ТПС зависит от этапности опадения генеративных побегов.
По этому признаку генеративные побеги изученных видов ив можно разделить на две группы: одноэтапно-опадающие ( S. aurita , S. lapponum ); двуэтапно-опадающие ( S. rosmarinifolia, S. myrtilloides ) (Nedoseko, Viktorov, 2017; Nedoseko, 2018).
Так как двуэтапно-опадающие генеративные побеги опадают в два этапа, и их нижняя олиственная часть остается на двулетнем побеге до осени, то их, необходимо учитывать в составе ТПС. Учитывая долговечность вегетативных частей генеративных побегов, вариант ветвления и размер зоны отмирания вегетативных побегов у изученных видов можно выделить следующие архитектурные модули:
-
1 – модуль I, основанный на мезотонном ветвлении с одноэтапно-опадающими генеративными побегами. Характерен для кустарников средней величины ( S. aurita ) (рис. 5);
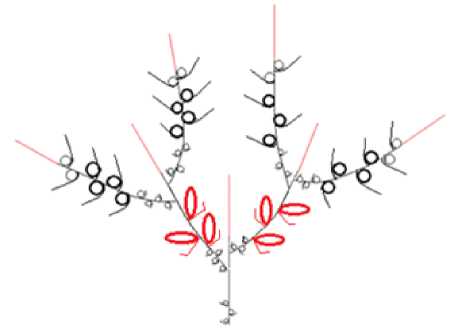
Рис. 5. Архитектурный модуль I / Fig. 5. Architectural module I
-
2 – модуль II, основанный на базитонном ветвлении с одноэтапно-опадающими генеративными побегами. Характерен для низких кустарников ( S. lapponum ) (рис. 6);
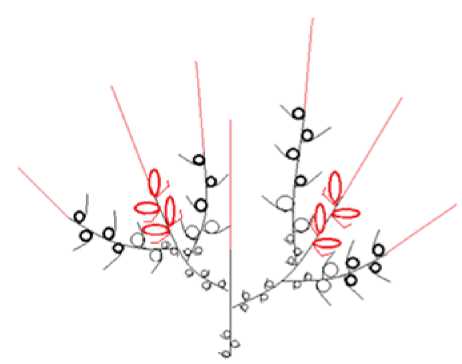
Рис. 6. Архитектурный модуль II / Fig. 6. Architectural module II
-
3 – модуль III, основанный на базитонном ветвлении с двуэтапно-опадающими генеративными побегами. Характерен для низких кустарников S. rosmarinifolia, S. myrtilloides (рис. 7).
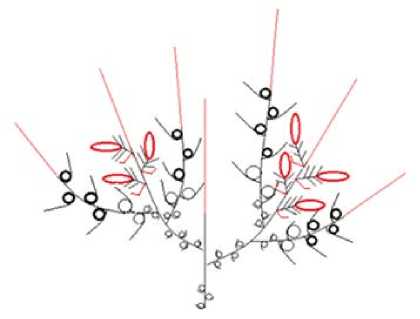
Рис. 7. Архитектурный модуль III / Fig. 7. Architectural Module III (red shows dying parts of shoots and falling parts of generative shoots)
(красным цветом показаны отмирающие части побегов и опадающие части генеративных побегов)
Условные обозначения
О- - спяшая почка,
О * вегетативная почка,
О - генеративная почка,
__У- лист,
—I - почечная чешуя,
О- соцветие,
^ - вегетативная часть генеративного побега
Спящая почка – inactive bud вегетативная почка – vegetative bud генеративная почка – fruit bud лист – leaf почечная чешуя – bud scale соцветие – inflorescence вегетативная часть генеративного побега – the vegetative part of the generative shoot
З АКЛЮЧЕНИЕ
Согласно литературным данным ивы сформировались в начале третичного периода (Krishtofovich, 1957; Graham, 1964; Pavlyutkin, 2002; Ilinskaya, Klimova, 2006) в условиях бессезонного климата. Первичной ареной трансформации жизненных форм ив исследователи считают поймы рек (Skvortzov, 1968; Afonin, 2006; Mazurenko, 2010; Getmanets, 2011). В дальнейшем бореальные виды ив эволюционировали в различных адаптивных зонах (Afonin, 2006; Getmanets, 2011). В каждой зоне сформировались жизненные формы, наиболее полно соответствующие тем или иным экологическим условиям. К анцестральной жизненной форме ив относят деревья и высокие кустарники с прямостоячими побегами, растущие по берегам рек (Skvortzov, 1968). Выходя за пределы пойм, жизненные формы ив стали претерпевать изменения. На открытых участках заболоченного луга, по окраинам осоковых болот формировалась вегетативно-подвижная жизненная форма эпигеогенно-геоксильный кустарник (рис. 1А, 2А). При этом кустарники становились более низкими, базальные части их анизотропных стволиков укоренялись придаточными корнями, погружаясь в субстрат, куст расширяется, увеличивается число стволиков и усиливается вегетативное разрастание за счет ксилоризомов.
На затененных, участках по окраинах осоковых и сфагновых болот при наличии верхнего яруса древесной растительности образовалась жизненная форма гипогеогенно-геоксильного кустарника (рис. 1Б, 2Б). Эпигеогенно-геоксильные кустарники чаще растут вне полога леса, а гипогеогенно-геоксильные кустарники растут под пологом древесной растительности. Вероятно, это связано с повышенной светолюбивостью эпигеогенно-геоксильных форм . Данные жизненные формы характерны в основном для S. rosmarinifolia, S. aurita. На сухих субстратах под пологом древесного яруса сформировалась жизненная форма «деревце» ( S. aurita, ) (3). Особи данной жизненной формы – небольшой высоты, с одним стволиком и низкорасположенной кроной с небольшим числом вариантов побегов.
На болотистых субстратах скелетные оси погружались в моховую подушку, укоренялись там с помощью придаточных корней, образуя жизненную форму низкий длинноксилоризомный гипогеогенно-геоксильный кустарник (рис.4). Растения S. myrtilloides и S. lapponum данной жизненной формы произрастают в условиях олиготрофных болот, обводненных с поверхности. Болотные виды имеют небольшую продолжительность онтогенеза, выработали следующие приспособления: способность быстрого укоренения, образование длинных ксилоризомов, небольшую высоту особей и уменьшение разнообразия побегов и побеговых систем в кроне.
В качестве основной структурной единицы побеговой системы ив мы рассматриваем трехлетнюю побеговую систему (ТПС), которая соответствует ЭПС – элементарной побеговой системе (Antonova, Lagunova, 1999). На основе ТПС с учетом трех признаков: вариант ветвления, размер зоны отмирания вегетативных побегов (число отмирающих верхних метамеров), долговечность вегетативных частей генеративных побегов у изученных видов ив мы выделяем три архитектурных модуля.
Каждой группе ив, относящихся к одинаковым жизненным формам, присущи разные архитектурные модули, что обусловлено разным вариантом ветвления и этапностью опадения генеративных побегов (табл. 1).
Таблица 1. Жизненные формы и архитектурные модули изученных видов ив
Table 1. Life forms and architectural modules of the studied types of willows
|
Виды ив Types of willows |
S. aurita |
S. rosmarinifolia |
S. myrtilloides |
S. lapponum |
|
Жизненные формы Life forms |
||||
|
Эпигеогенно-геоксильный кустарник Epigeogenic-geoxylic shrub |
+ |
+ |
||
|
Гипогеогенно-геоксильный кустарник Hypogeogenic- geoxylic shrub |
+ |
+ |
||
|
Деревце Tree |
+ |
|||
|
Низкий длинноксилоризомный гипогеогенно-геоксильный кустарник Low long-xylorizome hypogeogenic-geoxyl shrub |
+ |
+ |
||
|
Архитектурные модули Architectural modules |
||||
|
Архитектурный модуль I Architectural Module I |
+ |
|||
|
Архитектурный модуль II Architectural Module II |
+ |
|||
|
Архитектурный модуль III Architectural Module III |
+ |
+ |
||
Возможно, эта особенность позволяет ивам существовать в различных экологических условиях и проявлять поливариантность развития.
