Значение белковых компонентов в разнокачественной икре для эмбрионального развития и жизнестойкости личинок карповых рыб

Автор: Залепухин Валерий Владимирович
Журнал: Природные системы и ресурсы @ns-jvolsu
Рубрика: Биология и биотехнология
Статья в выпуске: 4 (10), 2014 года.
Бесплатный доступ
Эндогенная разнокачественность самок и икры является одним из важнейших факторов формирования ежегодного поколения, и ее оценка важна для понимания механизмов биохимических изменений в ближайший преднерестовый период. В условиях искусственного воспроизводства карповых рыб среди всех компонентов биохимического состава только содержание протеина и b-липопротеидов обнаруживают корреляции с эмбриональным развитием и выживаемостью не питавшихся личинок. Эта тенденция справедлива и для одомашненных рыб, и для рыб, выросших в естественных водоемах.
Карповые рыбы, искусственное воспроизводство, качество икры, белок, липопротеиды, эмбриональное развитие, выживаемость личинок
Короткий адрес: https://sciup.org/14967429
IDR: 14967429 | УДК: 591.5-1.05/639.371.5-215
The significance of protein components in heterogeneous eggs for embrionic development and larvas persistence of cyprinidae
Endogenous heterogeneity of females and their eggs is one of the most important factors of annual generation's formation in nature-spawn and incubative centers. Its assessment is significant for comprehending the notion of biochemical changes in proximate prespawn period. Between the shares of egg's biochemical composition of Cyprinidae species only protein and b-lipoproteids levels are incovering constant and authentic correlations with embrionic development and larval survival in the conditions of artificial reproduction. The promotion of protein's level to 20 - 24 % (in wet weigth) in percolating eggs positively affects the fertilization and larvae vitality. This trend is fair for the domesticated and nature-spawn fish. The same significance is important for the b-lipoproteids level in ovular eggs of grass carp (Ctenopharyngodon idella Val.). The optimal concentration is 100-200 mg % in wet weight.
Текст научной статьи Значение белковых компонентов в разнокачественной икре для эмбрионального развития и жизнестойкости личинок карповых рыб
Изучение биохимической разнокаче-ственности половых продуктов в условиях искусственного воспроизводства, несомненно, важно для совершенствования методов оценки качества производителей и получаемого потомства, поскольку именно в динамике состава яйцеклеток и сперматозоидов реализуются изменения обмена веществ производителей в период гаметогенеза, созревания и нереста. При искусственном разведении карпа и растительноядных рыб отличительной особенностью технологического цикла является быстрая потеря способности икры к оплодотворению (от 0,5 до 1,5 часов) по сравнению с моментом овуляции. Качество получаемой икры оказывает существенное влияние на количественные характеристики эмбрионального развития в период инкубации и выживаемость не питающихся личинок. Именно поэтому возни- кает необходимость перехода к оценке икры по биохимическим показателям, которые дают более точную информацию о способности к оплодотворению и нормальному развитию. Можно десятилетиями работать над созданием новой породы с наилучшим экстерьером и свести на нет все усилия лишь из-за несвоевременного отцеживания половых продуктов или неудачного осеменения в условиях искусственного воспроизводства.
У рыб как многоплодных животных (имеющих большую плодовитость) в условиях искусственного разведения влияние самцов на жизнеспособность потомства гораздо слабее, чем самок. Это объясняется тем, что макромолекулы, имеющиеся в зрелых ооцитах и синтезированные в ходе развития, способны гораздо дольше обеспечивать развитие потомства по сравнению с ядром и цитоплазмой сперматозоидов. Использование для искусственного осеменения высококачественной спермы 3–6 самцов не только увеличивает внутривидовую гетерогенность потомства, усиливая тем самым генетический полиморфизм популяций, но и сводит к минимуму возможность выделения «отцовского фактора» в разнокачествен-ности потомства [5].
Объекты и методы исследования. Экспериментальный материал получен в 1990–2004 гг. в рыбоводных хозяйствах Астраханской и Волгоградской областей (V и VI зоны рыбоводства). Объектами исследований послужили зеркальный и чешуйчатый карп, белый амур (БА) и пестрый толстолобик (ПТ) из местных пользовательных стад, а также сазан и пестрый толстолобик, заготовленные на тонях Нижней Волги. В ходе анализа нами были выделены следующие группы икры в соответствии с ее рыбоводным качеством:
Группа I – «недозрелая» икра. Соответствует подфазе Е1 – незавершенной IV стадии зрелости гонад (СЗГ). У сазана и карпа характеризуется малым количеством полостной жидкости («густая» икра) и отцеживается малыми порциями. При обесклеива-нии образует трудно разбиваемые комки. У растительноядных рыб овариальной жидкости, наоборот, много, икринки мелкие и упругие на ощупь.
На контрольных гистологических препаратах и у карпа, и у растительноядных рыб не у всех самок в такой икре отмечен заметный сдвиг ядра к оболочке. Развитие осемененной икры характеризуется появлением разноразмерных бластомеров, большим количеством уродливых эмбрионов, погибающих задолго до вылупления. Процент оплодотворения низкий, не превышает 50 %, процент нормально развивающихся эмбрионов близок к нулю. У растительноядных рыб вылупившихся предличинок не отмечено, у сазана наблюдается единичный выклев.
Группа II и III – «зрелая икра», соответствующая IV завершенной СЗГ и подфазе Е2. Отцеживание икры быстрое. После оплодотворения дробление синхронное и равномерное, уродливых эмбрионов мало. К группе II (икра хорошего рыбоводного качества) отнесена икра с процентом оплодотворения 50–70 %, высоким процентом нормально развивающихся эмбрионов (80–90 %) и близким к нормативному процентом выхода предличинок (от 30 до 60 %). В группу III (икра отличного рыбоводного качества) выделена икра с максимальной оплодотворяемостью (более 70 %), наиболее высокими показателями нормально развивающихся эмбрионов (от 80 до 100 %) и выхода предличинок (более 60 %). Группы, следовательно, выделены по отношению к важнейшим рыбоводным характеристикам, выявляемым в ходе инкубации.
Группы IV и V – «постовулярно перезревшая икра» (передержанная в полости тела самок после овуляции при несвоевременном отцеживании). Соответствует IV СЗГ и IV– VI СЗГ. Характеризуется большими и заметно набухшими, легко деформируемыми при нажатии икринками, среди которых встречаются уже побелевшие. Дробление асинхронное, бластомеры разной формы часто отрываются от бластодиска. Процент оплодотворения может быть и низким, и высоким, но из-за множества уродливо развивающихся эмбрионов процент выхода предличинок незначителен (5–10 %). К группе IV отнесена икра, еще сохранившая способность к оплодотворению (подфаза Е2), а к группе V – полностью ее утратившая (подфазы Е3 – F). У сазана и карпа до 10 % икринок такую способность сохраняют вследствие субпорционного характера созревания ооцитов и выметывания икры.
Количество общего белка в икре определяли по Lowry et al. [9], а в полевых условиях – по биуретовой реакции [1]; фракционный состав белков – методом электрофореза в блоке 5 %-ного полиакриламидного геля в сравнении с параллельной разгонкой маркеров – белков сыворотки крови человека, яичного овальбумина и бычьего альбумина. b -ли-попротеиды в гомогенате икры – по Бурштейну и Самаи [там же].
Взаимосвязи между биохимическим составом икры, с одной стороны, и показателями эмбрионального развития и жизнестойкостью не питавшихся личинок – с другой, изучались с помощью статистического, корреляционного и регрессионного анализа.
Полученные результаты . Показателем, устойчиво связанным с качеством икры БА и
ПТ, является содержание общего белка. Существует немало видов, у которых данный параметр обнаруживает те или иные взаимосвязи с рыбоводными характеристиками развивающейся икры, и прежде всего – с ее оплодотворяе-мостью. Наибольшее его содержание у всех изученных нами видов нами отмечено в зрелой икре хорошего и отличного качества (табл. 1 и 2). Следует отметить, что при экзогенном стимулировании созревания в период, непосредственно предшествующий овуляции, продолжается поступление в яйцеклетки различных органических и неорганических соединений – об этом свидетельствует динамика биохимического состава недозрелой и зрелой икры (соответственно I, II и III группы).
Постовулярное перезревание икры сопровождается началом протеолиза, что вызывает уменьшение содержания белка, все более заметное по мере углубления процессов дегенерации. Перезрелая икра, сохраняющая способность к оплодотворению (IV группа), имеет относительно высокий уровень белка; икра, полностью утратившая такую возможность (V группа), характеризуется самым низким его уровнем (табл. 1–
2). Такие тенденции характерны для всех трех видов и по всем годам исследований.
Корреляционный анализ подтвердил существование достоверных связей между содержанием общего белка в овулировавшей икре двух видов растительноядных рыб с основными показателями эмбрионального развития и длиной вылупившихся предличинок (см. табл. 3). У сазана же не удалось выявить достоверных связей с рыбоводными показателями инкубации ни для одного биохимического параметра – у этого вида, сохраняющего в экологических условиях Нижневолжского региона субпорционность икрометания, при искусственном воспроизводстве показатели эмбрионального развития связаны с массой овулировавших икринок. Для белого амура и пестрого толстолобика, имеющих единовременное икрометание, содержание белка в овулировавшей икре может быть использовано в виде биохимического критерия качества, тем более что методика его определения по биуретовой реакции несложна, не требует дефицитного оборудования и реактивов и вполне применима в полевых условиях.
Таблица 1
Содержание белка в разнокачественной икре (% сырой массы), 2001–2002 гг.
|
Виды рыб |
Группы икры |
2001 г. |
2002 г. |
|
Сазан ( n = 36) |
I |
18,90 ± 0,94 |
20,27 ± 0,30 |
|
II–III |
21,05 ± 0,91 |
21,47 ± 0,40 |
|
|
IV–V |
16,97 ± 0,70 |
19,10 ± 1,32 |
|
|
Белый амур ( n = 29) |
I |
19,30 ± 1,01 |
17,57 ± 1,29 |
|
II–III |
21,23 ± 0,39 |
18,81 ± 0,79 |
|
|
IV–V |
17,50 ± 0,69 |
16,90 ± 0,42 |
|
|
Пестрый nолстолобик ( n = 30) |
I |
18,21 ± 0,82 |
18,40 ± 0,49 |
|
II–III |
20,28 ± 0,76 |
21,79 ± 0,67 |
|
|
IV–V |
16,29 ± 0,81 |
15,52 ± 0,77 |
Таблица 2
Белок в разнокачественной икре карповых рыб (% сырой массы), 2003 г.
|
Группы икры |
Сазан ( n = 35) |
Белый амур ( n = 38) |
Пестрый толстолобик ( n = 41) |
|
I |
18,74 ± 1,18 |
18,54 ± 0,18 |
20,04 ± 0,89 |
|
II |
20,24 ± 1,20 |
19,93 ± 0,96 |
19,69 ± 0,77 |
|
III |
20,23 ± 0,34 |
20,74 ± 0,51 |
20,53 ± 0,52 |
|
IV |
19,54 ± 1,39 |
20,60 ± 1,59 |
19,12 ± 0,64 |
|
V |
19,01 ±1,23 |
17,16 ± 0,84 |
18,15 ± 1,34 |
|
Резорбирующаяся икра (48 часов после овуляции) |
8,52 ± 0,92 |
13,64 ± 1,09 |
11,10 ± 0,84 |
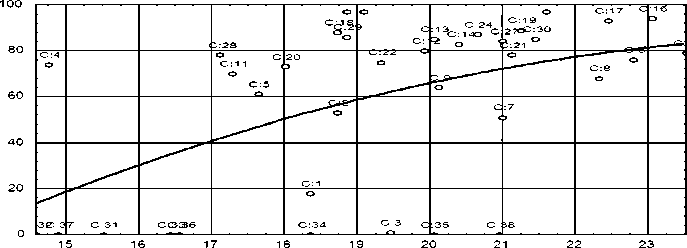
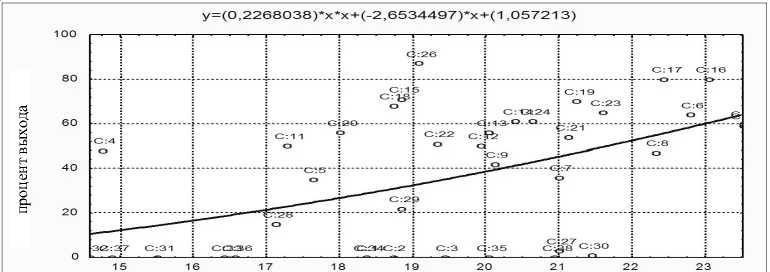
Регрессионный анализ подтверждает взаимосвязь параметров эмбрионального развития с ростом количества белка в овулировавшей икре (см. рис. 1–3).
Однако при анализе электрофоретического спектра белков нам не удалось выявить взаимосвязей между содержанием отдельных фракций и характеристиками эмбрионального развития. Это может означать, что для полноценного развития эмбрионов и личинок важен весь белковый пул в овулировавшей икре.
Значительная роль в процессах созревания гонад и построении мембран яйцеклеток и развивающихся зародышей рыб принадлежит липопротеидам. Среди них наиболее важны b-липопротеиды - белково-липид-ные комплексы с подвижностью b—глобулинов (по современной классификации – липопротеиды низкой плотности, ЛПНП). Они часто встречаются в крови половозрелых самок в период вителлогенеза, перенося в развивающиеся ооциты липовителлин – комплекс, содержащий, помимо белков и липидов, также фосфор и кальций [4]. Изучению динамики b-липопротеидов в крови рыб посвящено немало работ (см.: [2] и др.), но данных по содержанию этих соединений в икре мы не встретили.
Как показали наши исследования, в икре сазана содержится значительно больше b —ли-попротеидов, чем у белого амура и пестрого толстолобика (см. рис. 4). Количественное определение показывает, что в недозрелой и зрелой икре (I, II и III группы) концентрация b -ли-попротеидов меняется незначительно (Р > 0,5). Передержка икры после овуляции, даже при сохранении оплодотворяемости (IV группа), вызывает значительное снижение уровня b -липоп-ротеидов только у белого амура (P < 0,05), у других видов различия несущественны (P > 0,5). Лишь сильное постовулярное перезревание, сопровождающееся полной потерей способности к оплодотворению (V группа), связано со значительным падением содержания b -липоп-ротеидов у всех трех рассматриваемых видов (P < 0,05). Это согласуется с увеличением ко-
Таблица 3
Коррелятивные зависимости ( r xy ) между общим биохимическим составом икры и основными рыбоводными показателями
|
Вид |
Биохимические показатели |
Рыбоводные показатели |
|||
|
Процент оплодотворения |
Процент нормально развивающихся эмбрионов |
Процент выхода |
Длина пред-личинок на выклеве |
||
|
Сазан |
Вода |
-0,027 |
-0,095 |
-0,121 |
+0,089 |
|
Сухое вещество |
+0,027 |
+0,095 |
+0,121 |
-0,089 |
|
|
Белок |
+0,104 |
+0,243 |
+0,224 |
+0,148 |
|
|
Жир |
+0,161 |
-0,041 |
-0,132 |
+0,139 |
|
|
Мин. остаток |
-0,141 |
-0,198 |
-0,152 |
-0,243 |
|
|
«белок / жир» |
-0,095 |
+0,113 |
+0,152 |
-0,074 |
|
|
Белый амур |
Вода |
-0,179 |
-0,107 |
-0,085 |
-0,281 |
|
Сухое вещество |
+0,179 |
+0,107 |
+0,085 |
+0,281 |
|
|
Белок |
+0,516 |
+0,461 |
+0,468 |
+0,490 |
|
|
Жир |
+0,150 |
+0,171 |
+0,140 |
+0,164 |
|
|
Мин. остаток |
-0,238 |
-0,265 |
-0,257 |
-0,142 |
|
|
«белок / жир» |
+0,161 |
+0,118 |
+0,143 |
+0,127 |
|
|
Пестрый толстолобик |
Вода |
-0,362 |
-0,236 |
-0,247 |
-0,294 |
|
Сухое вещество |
+0,362 |
+0,236 |
+0,247 |
+0,294 |
|
|
Белок |
+0,382 |
+0,370 |
+0,381 |
+0,309 |
|
|
Жир |
-0,042 |
+0,213 |
+0,219 |
+0,059 |
|
|
Мин. остаток |
+0,048 |
-0,144 |
-0,117 |
+0,105 |
|
|
«белок / жир» |
+0,182 |
-0,088 |
-0,119 |
+0,015 |
|
Примечание . В корреляционной таблице выделены достоверные коэффициенты корреляции при n = 3 5 + 41 Р 0 05 > 0,3 3, Р 001> 0,42. Все расчеты проведены по сырой массе.
личества свободных жирных кислот в перезрелой икре, образующихся при распаде липопротеиновых комплексов (помимо гидролиза фосфоглицеридов).
Среди трех изученных нами видов лишь у белого амура выявлены положительные корреляции содержания b-липопротеидов с плотностью овулировавших икринок (rxy = +0,434 при P < 0,01) и средним диаметром набухшей икры (rxy = +0,504 при P < 0,01). С концентрацией b-липопротеидов в овулировавшей икре белого амура достоверно коррелируют и основные показатели развивающейся икры (см. табл. 4), что дает основания рассматривать этот параметр как один из критериев качества икры у данного вида. Прямо противоположная тенденция наблюдается для диаметра овулировавшей икры (P < 0,01).
у=(-0,5347557)*х*х+(28,14957)*х+(-283,1 863)
белок, % сырой массы
Рис. 1. Зависимость оплодотворяемости от количества белка в овулировавшей икре белого амура (R = 0,53 при P < 0,01)
белок, % сырой массы
Рис. 2. Зависимость процента выхода предличинок от содержания белка в овулировавшей икре белого амура (R = 0,51 при P < 0,01)
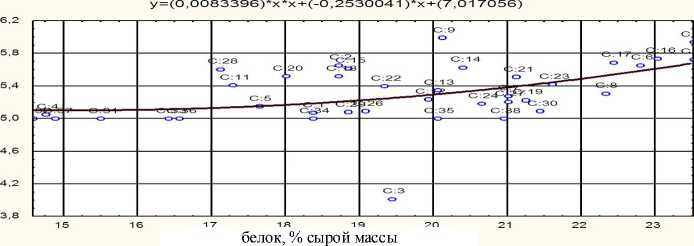
Рис. 3. Зависимость длины вылупляющихся предличинок от содержания белка в овулировавшей икре белого амура (R = 0,47 при P < 0,01)
Ни у одного из трех изученных видов не удалось выявить достоверных взаимосвязей количества b -липопротеидов в овулировавшей икре с размерно-весовыми характеристиками самок, их рабочей плодовитостью и упитанностью (табл. 4).
Поскольку важнейшей функцией липопротеидов является обеспечение избирательной проницаемости для клетки и ее органоидов [6], то уровень и динамику b-липопроте-идов в разнокачественной икре следует признать важной видовой особенностью белого амура, связанной, вероятно, со структурой мембран овулировавших яйцеклеток и развивающихся эмбрионов. Для трех изученных нами видов снижение уровня b-липопротеидов в икре (менее 250 мг % у сазана, 60 мг % у белого амура и 80 мг % у пестрого толстолобика) может служить биохимическим признаком постовулярного перезревания икры и полной потери оплодотворяемости.
Использование различных методов управления биопродукционным потенциалом размножающихся рыб, их физиологическим
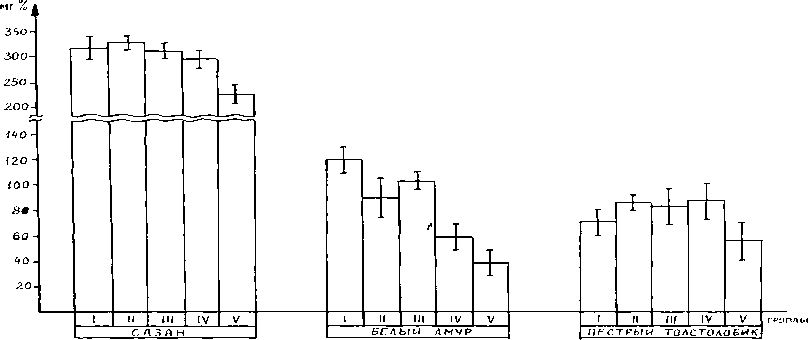
Рис. 4. b -липопротеиды в разнокачественной икре карповых рыб
Таблица 4
Корреляции ( r xy ) содержания b -липопротеидов в овулировавшей икре с биологическими характеристиками самок и развивающейся икры
|
Биологические характеристики |
Сазан |
Белый амур |
Пестрый толстолобик |
|
Возраст самок |
-0,054 |
+0,172 |
+0,095 |
|
Масса самок |
-0,272 |
+0,259 |
-0,013 |
|
Длина ( l ) самок |
-0,247 |
+0,067 |
-0,015 |
|
Высота тела самок |
-0,021 |
+0,312 |
-0,284 |
|
Индивидуальная рабочая плодовитость |
-0,311 |
+0,205 |
-0,121 |
|
Упитанность самок |
+0,074 |
+0,256 |
-0,027 |
|
Средняя масса овулировавших икринок |
-0,023 |
-0,272 |
+0,213 |
|
Средний диаметр овулировавших икринок |
+0,099 |
-0,469 |
+0,093 |
|
Плотность овулировавших икринок |
-0,145 |
+0,434 |
-0,011 |
|
Средний диаметр набухшей икры |
-0,173 |
+0,504 |
-0,053 |
|
Процент оплодотворения |
-0,166 |
+0,331 |
-0,096 |
|
Процент нормально развивающихся эмбрионов |
-0,003 |
+0,450 |
+0,005 |
|
Процент выхода предличинок |
-0,011 |
+0,382 |
+0,002 |
|
Длина вылупляющихся предличинок |
-0,004 |
+0,450 |
-0,062 |
|
n |
35 |
38 |
41 |
Примечание. В корреляционных таблицах выделены достоверные коэффициенты корреляции при n =
35 * 41 Р 0,05 > 0,33; Р 0,01 >0,42.
состоянием, качеством получаемых половых продуктов, ходом эмбрионального развития и выживаемостью личинок, несомненно, скажется на рыбоводных результатах выращивания и продуктивности водных экосистем в условиях аквакультуры. Наиболее важным является улучшение качественных характеристик (размеров и выживаемости) личинок к моменту перехода на активное питание – именно в этот период определяется возможность существования и дальнейшего развития при неблагоприятных факторах внешней среды.
На одомашненной форме сазана – зеркальном карпе – можно показать практически линейную зависимость между содержанием белка в овулировавшей икре и характеристиками личинок – их длиной и выживаемостью не питавшихся особей L100 (рис. 5–6).
Логичным путем увеличения содержания этих компонентов в икре является подкормка производителей в преднерестовый период смесями с повышенным содержанием протеина и углеводов. Именно такие предложения вносились учеными-рыбоводами Г.М. Прониным [8] и Н.И. Масловой [7], хотя они не анализировали выживаемость личинок, а исходили только из результатов инкубации или продуктивности сеголетков карпа.
Аналогичные зависимости между выживаемостью не питающихся личинок и содержанием в икре белка выявлены и для растительноядных рыб – белого амура и пестрого толстолобика (см.: [3]). Отсюда вытекает необходимость создания микрокапсулированных плавающих кормов с повышенным содержанием протеина для этих видов.
Таким образом, среди изученных составляющих биохимического состава только для белковых компонентов овулировавшей икры удается выявить корреляции и регрессионные зависимости с характеристиками эмбрионального развития и выживаемостью не питавшихся личинок.
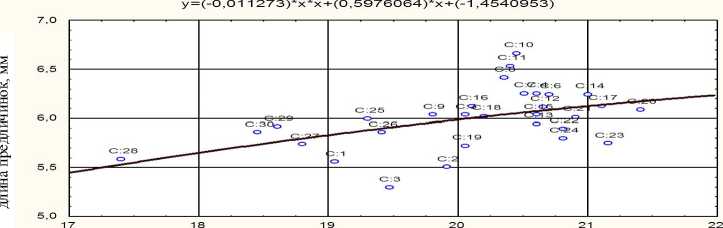
белок, % сырой массы
Рис. 5. Зависимость длины вылупляющихся предличинок от содержания белка в овулировавшей икре зеркального карпа (R = 0,48 при P < 0,01)
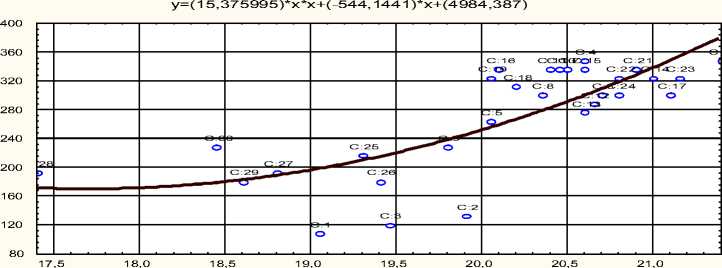
Рис. 6. Зависимость выживаемости L 100 (ось У – мм) от содержания белка в овулировавшей икре зеркального карпа (ось Х – часы)
Список литературы Значение белковых компонентов в разнокачественной икре для эмбрионального развития и жизнестойкости личинок карповых рыб
- Биохимические методы исследования в клинике. -М.: Медицина, 1969. -651 с.
- Васильева, О. Б. Липидный состав липопротеинов самок радужной форели Salmo gairdneri в годовом цикле/О. Б. Васильева, В. С. Сидоров, Е. И. Лизенко//Вопросы ихтиологии. -2004. -Т. 44, № 3. -С. 414-418.
- Залепухин, В. В. Концепция эндогенной разнокачественности в условиях искусственного воспроизводства карповых рыб: монография/В. В. Залепухин. -Волгоград: Волгоград. научн. изд-во, 2006. -320 с.
- Ипатов, В. В. Сывороточные белки рыб: гетерогенность, структура, функции/В. В. Ипатов, В. И. Лукъяненко//Успехи современной биологии. -1979. -Т. 38, вып. 1. -С. 108-124.
- Кирпичников, В. С. Генетика и селекция рыб/В. С. Кирпичников -Л.: Наука, 1987. -520 с.
- Климов, А. Н. Липопротеиды плазмы крови, их функции и метаболизм/А. Н. Климов//Биохимия липидов и их роль в обмене веществ. -М.: Наука, 1979. -С. 45-75.
- Маслова, Н. И. Физиолого-биохимические основы методов повышения продуктивных качеств племенных стад карпов: автореф. дис.... д-ра биол. наук/Маслова Неонила Ивановна. -М., 1990. -38 с.
- Пронин, Г. М. Влияние белковых компонентов пищи на биологические показатели карпа в преднерестовый период: автореф. дис.... канд. биол. наук./Пронин Геннадий Михайлович. -М., 1981. -18 с.
- Protein measurement with Folin phenile reagent/O. H. Lowry, N. J. Rosenbrough, A. Z. Porr, R. J. Randoll//J. Biol. Chem. -1951. -Vol. 193, № 1. -P. 265-275.
