Зуб человека из пещеры Логово Гиены (Западный Алтай)
")
Автор: Чикишева Т.А., Васильев С.К., Орлова Л.А.
Журнал: Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий @paeas
Рубрика: Археология каменного века палеоэкология
Статья в выпуске: XIII, 2007 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/14521316
IDR: 14521316
Текст статьи Зуб человека из пещеры Логово Гиены (Западный Алтай)
Пещера Логово Гиены была открыта и раскопана в 1969 г. Н.Д. Оводовым [Галкина, Оводов, 1975]. По обилию и сохранности палеотериологи-ческого материала это местонахождение до настоящего времени не имеет себе равных среди пещер Горного Алтая. Глубина пещеры (или грота) -около 20 м, за освещённой центральной камерой она переходит в круто поднимающуюся короткую галерею, тёмную и сырую. В пределах центральной камеры выделено 4 слоя, общей мощностью до 1,5 м [Галкина, Оводов, 1975; Васильев и др., 2006].
Раскопки 2006 г. затронули небольшие по площади участки основного костеносного слоя 2, сохранившегося in sity в 5 пунктах в пределах центральной камеры [Васильев и др., 2006]. В пункте 3, расположенном в центральной части грота, под массивной плитой известняка, сохранился небольшой участок (около 0,5 м2), не затронутый раскопками 1969 г. Здесь был выбран насыщенный фаунистическими остатками костеносный слой, мощностью до 15-20 см. Именно отсюда происходит зуб палеолитического человека, найденный позднее, уже при переборке промытого грунта.
Из слоя 2 в 2006 г. было извлечено 2890 определимых костных остатков, принадлежащих: Homo sapiens, Lepus tanaiticus, Lepus tolai, Marmota baibacina, Castor fiber, Canis lupus, V. vulpes, Vulpes corsak, Cuon alpinus, Ursus arctos, Martes zibellina, G. gulo, Mustela erminea, Mustela nivalis, Mustela altaica, Mustela eversmanni,, Crocuta spelaea, Panthera spelaea, Mammuthus primigenius, Equus ferus, Equus ex.gr. hydruntinus, Coelodonta antiquitatis, Cervus elaphus, Megaloceros giganteus, Bos (Poёphagus) baicalensis, Bison priscus, Capra sibirica, Ovis ammon, Ovis-Capra. Среди более чем 8 тыс. остатков мелких млекопитающих отмечены: Chiroptera gen indet, Asioscalops altaica, Sorex sp., S. minutissimus, S. minutus, Crocidura sp., Sciurus vulgaris, Eutamias sibiricus, Pteromys volans, Spermophilus sp., Apodemus sp., Cricetus cricetus, Citellus sp., Cricetulus barabensis, Allocricetulus eversmanni, Ellobius sp., Clethrionomys sp., C. rufocanus, C. rutilus, C. glareolus, Alticola sp., A. strelzowi, A. macrotis, Lagurus sp., L. lagurus, Eolagurus sp., Stenocranius gregalis, Microtus sp., M. oeconomus, M. hyperboreus, M. arvalis, M. agrestis,
M. middendirfii, Arvicola terrestris, M. myospalax, Allactaga sp., Ochotona alpina- hyperborean, Ochotona sp. [Васильев и др., 2006, 2007].
Обилие в слое 2 практически целых крупных фрагментов костей, указывает на чрезвычайно быстрое накопление основной части костеносного горизонта, что подтверждается и данными радиоуглеродного анализа. К полученной ранее дате (32700 ± 2800 лет) в 2006 г. добавилось 9 новых 14 С датировок, образцы для которых были отобраны из разных пунктов и слоёв пещерных отложений. Образец, найденный в слое 3 (100 см от поверхности грунта) оказался древнее 40000 лет, а 2 образца, отобранные выше основного костеносного слоя, из заполнения горизонтальной трещины в стене пещеры (глубина 20-30 см) показали возраст около 2424,5 тыс. лет. Костные остатки из основной части костеносного слоя (глубина около 40-80 см), датируются приблизительно 33-34,5 тыс. лет (табл. 1). Таким образом, имеющиеся датировки указывают, что основная часть костеносного слоя 2 отлагалась в завершающую треть каргинского времени.
Накопление палеофаунистических остатков в пещере Логово Гиены происходило без участия человека, в результате жизнедеятельности хищных зверей и птиц. Пещерные гиены собирали в своё логово останки павших или добытых ими животных со всех ближайших окрестностей пещеры. Среди представителей мегафауны в тафоценозе 2 слоя преобладают остатки лошадей (36,1%), из которых около 1/3 относится к очень крупной кабаллоидной Equus ferus, а 2/3 – к плейстоценовому ослу Equus ex. gr. hydruntinus. Бизону принадлежит 23.3% костей, 5% - архару и горному козлу. Среди хищных доминируют остатки гиены – 8,6%, что позволяет рассматривать пещеру Логово Гиены как сезонное убежище кланов пещерных гиен, использовавшиеся для выведения потомства [Барышников, 2005]: зубы Crocuta spelaea молочной генерации составляют здесь 37%.
Преобладание костей плейстоценового осла, бизона, присутствие остатков корсака, степного хоря, зайца-толая, гигантского оленя, байкальского яка, свидетельствует о развитии степных ландшафтов. Присутствие остатков чисто лесных видов весьма незначительно.
В природе гиены выполняют роль санитаров, поедая трупы павших животных. Их мощные челюсти с крупными коническими зубами способны дробить, а желудок, содержащий необычайно агрессивные кислоты – переваривать и усваивать практически любые костные остатки. В то же время современная пятнистая и, по всей видимости, пещерная гиены – это весьма активные коллективные охотники. В плейстоцене в высоких широтах Евразии существование пещерных гиен на диете, состоящей исключительно из падали, без высококалорийной свежей добычи, было, очевидно, невозможно, в силу чего Crocuta spelaea являлись ещё более активными охотниками, чем современные африканские пятнистые гиены [Барышников, Верещагин, 1997]. Материалы из Логова Гиены показывают, что пещерные гиены затаскивали в пещеру прежде всего дистальные отделы конечностей и головы жертв, т.е. то, что обычно остаётся после поедания
Таблица 1. Пещера Логово Гиены. Радиоуглеродные значения возраста костных остатков
Среди материалов промывки обнаружен второй правый верхний премоляр постоянной смены (Р2) человека. Учитывая редкость плейстоценовых палеоантропологических находок, мы считаем, что описание любого объекта такой древности может представлять самостоятельный интерес.

Рис. 1. Зуб человека из пещеры Логово Гиены.
1 – мезиальная проекция; 2 – вестибулярная проекция; 3 – дистальная проекция; 4 – лингвальная проекция.
Состояние зуба. Эмаль коронки хрупкая, частично отслоилась, её крупный фрагмент с дисто-лингвального угла зуба был зафиксирован клеем Cemedin C.
Критерии, по которым была определена позиция найденного зуба на челюсти.
Вторые верхние премоляры представляют собой зубы с небольшой двубугорковой коронкой, имеющей эллиптическую форму, с одним округлым в сечении корнем. На корне может быть выражена бороздка, показывающая место разделения корня. Бугорки либо равны по размерам, либо вестибулярный несколько превосходит лингвальный. Межбугорковая борозда большей частью прямая и непрерывная. Вершина лингвального бугорка сдвинута в мезиальном направлении.
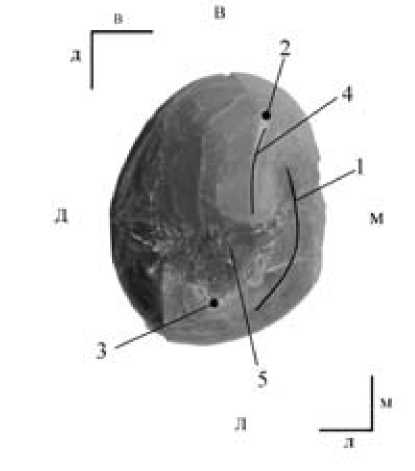
Рис. 2. Окклюзиальная проекция зуба.
1 – стёртость окклюзиальной поверхности; 2 – вестибулярный бугорок;
3 лингвальный бугорок; 4 – основной гребень вестибулярного бугорка;
5 – межбугорковая борозда.
Рассматриваемый нами зуб имеет один корень (рис. 1, 2,3,4). Коронка зуба вытянута в вестибуло-лингвальном направлении. Вестибулярный бугорок превосходит по размерам лингвальный (рис.5. 2, 5.3). Небольшая стёртость наблюдается с мезио-лингвальной стороны коронки (рис.5.1). Межбугорковая борозда глубокая и непрерывная (рис. 5.5).
Диагностические признаки зуба. Верхние премоляры Homo sapiens не обнаруживают сколько-нибудь значительной вариабельности в отношении формы коронки, которая могла бы иметь важное групподифференцирующее значение или имела более-менее определённый географический вектор. Выделяются некоторые популяции, характеризующиеся небольшими частотами одонтом (дополнительных бугорков на окклюзиальной поверхности), дополнительных бугорков на мезиальном и дистальном краевых гребнях [Scott, Turner, 1997]. Существует также система, разделяющая верхние премоляры на три типа по визуальной оценке соотношений размеров вестибулярного и лингвального бугорков. Но три типа, выделяемые по этой оценке, не имеют чётких границ и допускают значительный субъективизм в их определении [Зубов, 1973]. По данным А.А.Зубова, тип 1 (при котором вестибулярный бугорок в сильной степени превосходит лингвальный), часто встречается в арктических группах [там же, с. 109].
В одонтологических сериях разных историко-культурных периодов Южной и Западной Сибири универсальным для обоих верхних премоляров является тип 2 (при котором вестибулярный бугорок несколько больше лингвального) (неопубликованные данные Т.А.Чикишевой). На анализируемом нами зубе, эмаль которого разрушена на дистальной половине коронки, трудно уверенно провести диагностику соотношения его бугорков. Видимо, форма зуба ближе к типу 1.
Пол субъекта, к которому относился обнаруженный зуб определить пол морфологическим критериям невозможно. Такую диагностику в настоящее время могут провести молекулярные генетики.
Возраст субъекта с определённой долей условности может быть определён по интенсивности стёртости окклюзиальной поверхности зуба. Поскольку на стёртом участке не обнажён дентин, можно предположить молодой возраст индивида (в пределах конца второго – начала третьего десятка).
Патологические особенности. Корень зуба являет собой ярчайший пример гиперцементоза или оссифицирующего периодонтита . При этой патологии происходит избыточное формирование цемента зуба (грубоволокнистой костной ткани, окружающей поверхность корня). Гиперцемен-тоз возникает при хроническом периодонтите, пародонтите, аномалии развития зуба. Функционально он укрепляет зуб в альвеоле. Множественный гиперцементоз может наблюдаться при гиповитаминозе А или гипервитаминозе D.
