A study of human bones from a dwelling at Ust-Voikar, in the subarctic zone of Western Siberia

Author: Batanina O.V., Garkusha Y.N., Novikov A.V., Pozdnyakov D.V., Zubova A.V.
Journal: Archaeology, Ethnology & Anthropology of Eurasia @journal-aeae-en
Section: The metal ages and medieval period
Article in issue: 4 т.47, 2019.
Free access
Short address: https://sciup.org/145145456
IDR: 145145456 | DOI: 10.17746/1563-0110.2019.47.4.140-153
Text of the review article A study of human bones from a dwelling at Ust-Voikar, in the subarctic zone of Western Siberia
The Ust-Voikar settlement is located in the Shuryshkarsky District of the Yamal-Nenets Autonomous Okrug, on the left bank of the Gornaya Ob branch (one of the channels of the Malaya Ob River), northeast of the mouth of the Voikarsky Sor. The first researchers of the site identified it with Fort Voikar, a settlement known from written sources and folklore (Fedorova, 2006: 11). This was one of the aboriginal medieval “forts” that served as local centers of different functions for the natives of the northwestern Siberia (Perevalova, 2004: 214). According to the dendrochronological data for the wooden buildings found during the first excavation at the site in early 2000s, the early dwellings are dated to the late 13th to early 14th centuries AD. A later period of building activity at the site likely falls into the second half of the 17th century, and single dwellings were built during the 19th century (Gurskaya, 2008: 218, 223; Fedorova, 2006: 16).
The ethnic composition of the population of the settlement is still a question open for debate. But taking into account the known facts of the ethnic history of the northern part of the Lower Ob region, this population can be preliminary classified as Ugro-Samoyedic, while the presence of a Komi-Zyryan component can be reasonably
suggested as well. The zone of northern taiga where the site is located was historically, during medieval and modern times, an area of intense contacts between the ethnic groups mentioned above. Those contacts led to the formation of the northern (Lower Ob) group of the Khanty, in particular the ethnic group of the Voikar Khanty, which emerged as a result of the ethnogenetic processes taking place in the Voikar River basin (Martynova, 1998, 2005; Perevalova, 2004; and others).
In 2016, an extraordinary archaeological object— a double burial intentionally placed into the hearth of dwelling No. 11 (according to the numeration for the objects excavated since 2012)—was found at the site (Novikov et al., 2016). The building was of the frame-and-pillar type of construction, with a separate internal space, the walls of which were made using the plow technique. The external walls were made of vertically placed planks. The hearth, of rectangular shape, occupied the central part of the internal space. Dwellings of similar construction have previously been described for another aboriginal settlement in the same region, Fort Nadym (Kardash, 2009: 56–58).
The deceased were placed inside a wooden frame delineating the hearth (Fig. 1). The width of the construction was not large enough to bury the bodies of two adult people. Thus, one of the individuals was in a supine position on his back, while the second was on his left side with the legs bent at the knees. The heads of both were oriented westwards, and their legs towards the entrance of the dwelling. No trace of cremation was observed in the remains. The burial was filled with wood chips and coal-ash fractions that were probably taken from the cooled filling of the hearth.
It is of note that later, above the ruin of this dwelling, two new log houses were built one after the other, inside the area delimited by the ruin and preserving the original plan of the preceding building. Despite these reconstructions, the position of the hearth has been intact during the whole period, and the burials have been for a long time under an active fireplace. As a result, the total thickness of the layer of coal and ash covering the burials was 50–55 cm.
Dwelling No. 11 and the adjacent dwelling were dated using tree-ring analysis: they were most likely built in the middle of the last third of the 17th century, and the early 18th century, respectively*. It was not possible to determine precisely the time of inhumation; thus, it may be broadly dated to the whole period of construction, i.e. from the last third of the 17th century to the early 18th century.
The main purpose of this work was to study the human remains from Ust-Voikar as comprehensively as possible,
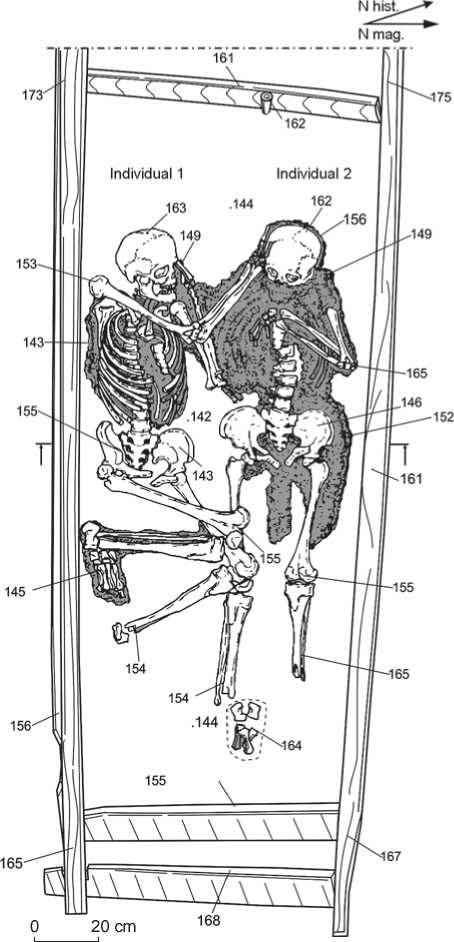
Fig. 1 . Burial 1 in the hearth of dwelling No. 11 (the area of dispersion of textile remains is depicted in gray).
using a variety of methods available to the physical anthropologist, in order to reconstruct the way of life, and the diet, of these people.
Methods
A complex protocol involving cranial metrics and dental traits was used to infer the possible origins and population affinities of the buried individuals (Alekseev, Debets, 1964; Zubov, 2006; Zubova, 2013). A description of their dental pathologies was carried in order to reconstruct the health conditions and diets of the individuals, with a distant aim of reconstructing their social status (Angel, 1984; Goodman, Martin, Armelagos, 1984; Buzhilova, 1998: 128).
Postcranial metrics were collected following a conventional protocol; stature and skeletal proportions were reconstructed based on the raw measurements (Alekseev, 1966). The individual values of the postcranial metrics were assigned to the categories developed by D.V. Pezhemsky (2011).
The postcranial non-metric protocol included two parts. The first was devoted to describing the morphology of entheseal sites, using the grade system of V. Mariotti for assessing muscular activity, enthesophyte development, and erosion of bone tissue (Mariotti, Facchini, Belcastro, 2004). Only muscles with a similar type of attachment to bone were employed, in order to avoid a bias in scoring the grades (Razhev, 2009: 253–254). The second part dealt with porous changes in articular surfaces and deformations of the contours of the joints. All large joints, as well as surfaces of the cervical, thoracic, and lumbar vertebrae, were studied.
Results
Ages at death of the deceased
A detailed examination of the two skulls and postcranial skeletons led us to the conclusion that the previously determined ages at death of the individuals (35–40 years for one, and 45–50 for another) (Novikov et al., 2016) need revision. Different skeletal markers of age (degree of suture closure, epiphyseal fusion, dental and articular surface status) contradict each other. Incomplete fusion of the vertebral arches (well-defined fusion lines are observed), heads of the ribs, iliac crests, clavicular proximal diaphyses, and acromions (only individual 2) suggest that the age at death of both individuals should be placed between 22 and 25 years (Schaefer, Black, Scheuer, 2009). But the status of cranial suture closure points towards an older age at death: 35–40 years for individual 1, and not less than 45–55 years for individual 2. The dental age of the two is determined as 40–45 and 45–50 years, respectively. The established discrepancy between the velocity of epiphyseal fusion, suture closure, and the degree of teeth wear is probably explained by a pathological process of endogenous character. If all the skeletal age markers are considered together, the age at death of both deceased may be broadly determined as adultus – maturus .
Cranial and dental morphology
Skelet on 1. The neurocranium is of small length and medium width. The cranial index lies at the border between meso- and brachycranic values. The cranial height measured from porion displays a very small value, while the height measured from basion is medium. The frontal arch is the longest component of the sagittal arch. The frontal bone is moderately wide at the level of the temporal lines, but is substantially wider at the coronal suture. The squama of the frontal bone is convex, only slightly inclined, but fairly protruding in the transverse plane. The robustness of the supraorbital region is weakly pronounced. The face is mesognathic according to the angles of vertical profile, but orthognathic according to the index of facial protrusion. The face is low, medium in width, and strongly horizontally protruding at both levels. The orbits are of medium width and height, of mesoconchal shape. The piriform aperture is mesorrhine; low and medium in width. The dimensions of the nasal bones are large at the level of dacryon, while simotic width and subtense are medium. The nasal bones are strongly protruding. All main dimensions of the mandible are medium, excluding the height of the ramus and the condylar width: these variables display large values (Table 1).
A spacing is observed between the central teeth; the cutting edge of the incisors is straight; lingual or vestibular shoveling is absent, as is the case for cingular derivatives. The degree of reduction of the hypocone of the upper second molars is moderate (4–), the Carabelli cusp is absent. In the distal part of all three molars in the raw, elements of the posterior fovea are observed. Both first and second lower premolars are strongly differentiated. The first molars display at least five cusps (a sixth cusp might have been also present), the second molars four cusps, and the third molars five cusps. A distal accessory tubercule is present in the second and third molars, while its presence or absence cannot be scored for the first molars owing to their strong attrition. The vestibular cingulum is enlarged. Protostylids are present in the third molars, while C7 is found in the second molars. The distal trigonid crest and epicristid are absent, the degree of convexity of the axial crest of the metaconid cannot be scored owing to its strong attrition. All three lower molars exhibit an “X”-pattern of grooves.
Skeleton 2. The skull displays mesocrany, and is more elongated than that of skeleton 1. The height of the vault is small, both from porion and basion . As in Skeleton 1, the frontal arch is the longest element of the sagittal contour. This bone is fairly convex, weakly inclined, medium in width, and displays a weak robustness of the supraorbital region. The face is low and moderately wide, clinoprosopic. The widths of the orbits and piriform aperture are larger than in skeleton 1. The nasal bridge is more protruding at dacryon, the angle of protrusion of the nasal bones is smaller, the simotic width is larger, and the simotic index smaller. The mandible is slightly taller and wider than that of skeleton 1 (Table 1).
Table 1. Cranial metrics of individuals 1 and 2
|
Variable |
1 |
2 |
|
1 |
2 |
3 |
|
1. Cranial length |
175 |
182 |
|
8. Maximum cranial breadth |
140 |
142 |
|
8 : 1. Cranial index |
80.0 |
78.0 |
|
17. Cranial height (basion-bregma) |
134 |
130 |
|
17 : 1. Height-length index (from basion) |
76.6 |
71.4 |
|
17 : 8. Height-transversal index (from basion) |
95.7 |
91.5 |
|
20. Cranial height (porion-bregma) |
105 |
103 |
|
20 : 1. Height-length index (from porion) |
60.0 |
56.6 |
|
20 : 8. Height-transversal index (from porion) |
75.0 |
72.5 |
|
5. Cranial base length |
100 |
103 |
|
9. Minimum frontal breadth |
94 |
94 |
|
10. Maximum frontal breadth |
123 |
117 |
|
9 : 10. Frontal index |
76.4 |
80.3 |
|
9 : 8. Fronto-transversal index |
67.1 |
66.2 |
|
11. Cranial base breadth |
121 |
115 |
|
12. Occipital breadth |
115 |
|
|
29. Nasion-bregma chord |
111 |
113.6 |
|
30. Bregma-lambda chord |
108.5 |
112 |
|
31. Lambda-opisthion chord |
93.5 |
97.5 |
|
26. Sagittal frontal arch |
128 |
135 |
|
27. Sagittal parietal arch |
121 |
125 |
|
28. Sagittal occipital arch |
112 |
119 |
|
25. Total sagittal arch |
361 |
379 |
|
26 : 25. Fronto-sagittal index |
35.5 |
35.6 |
|
27 : 25. Parieto-sagittal index |
33.5 |
33.0 |
|
28 : 25. Occipito-sagittal index |
31.0 |
31.4 |
|
28 : 27. Occipito-parietal index |
92.6 |
95.2 |
|
29 : 26. Frontal curvature index |
86.7 |
84.1 |
|
h. Transverse frontal curvature subtense |
20 |
20 |
|
h : 9. Transverse frontal curvature index |
21.3 |
21.3 |
|
Transverse frontal curvature angle |
133.9 |
133.9 |
|
Sub.NB. Longitudinal frontal curvature subtense |
26 |
31 |
|
Sub.NB. : 29. Longitudinal frontal curvature index |
23.4 |
27.3 |
|
Occipital curvature height |
25 |
23 |
|
45. Bizygomatic breadth |
134 |
133 |
|
9 : 45. Fronto-bizygomatic index |
70.1 |
70.7 |
|
45 : 8. Transversal facio-cerebral index |
95.7 |
93.7 |
|
40. Basion-prosthion length |
95 |
99 |
|
40 : 5. Facial protrusion index |
95.0 |
96.1 |
|
48. Nasion-alveolare height |
68 |
68 |
|
48 : 17. Vertical facio-cerebral index |
50.7 |
52.3 |
|
47. Nasion-gnathion height |
116.5 |
113 |
Table 1 (continued)
|
1 |
2 |
3 |
|
43. Upper facial height |
102 |
107 |
|
46. Midfacial breadth |
95 |
101 |
|
60. Alveolar length |
53 |
56 |
|
61. Alveolar breadth |
62.5 |
65 |
|
61 : 60. Alveolar index |
117.9 |
116.1 |
|
62. Palate length |
45.5 |
46 |
|
63. Palate breadth |
38.5 |
42.5 |
|
63 : 62. Palate index |
84.6 |
92.4 |
|
55. Nasal height |
50 |
50 |
|
54. Nasal breadth |
24.5 |
26.4 |
|
54 : 55. Nasal index |
49.0 |
52.8 |
|
51. Orbital breadth from mf. |
41 |
44.5 |
|
51а. Orbital breadth from d. |
38 |
42 |
|
52. Orbital height |
31.5 |
33.2 |
|
52 : 51. Orbital index from mf. |
76.8 |
74.6 |
|
52 : 51а. Orbital index from d. |
82.9 |
79.0 |
|
Frontomalareorbitale (bimalar) breadth |
95 |
99.5 |
|
Subtense from nasion to bimalar breadth |
18 |
20.5 |
|
Zygomaxillary breadth |
100 |
101 |
|
Subtense from subspinale to zygomaxillary breadth |
28.5 |
26 |
|
Nasomalar angle |
138.6 |
135.2 |
|
Zygomaxillary angle |
120.6 |
125.6 |
|
SC. Simotic chord |
8 |
12.2 |
|
SS. Simotic subtense |
4 |
5 |
|
SS : SC. Simotic index |
50.0 |
41.0 |
|
MC. Maxillofrontal chord |
20 |
18 |
|
MS. Maxillofrontal subtense |
6.5 |
7 |
|
MS : MC. Maxillofrontal index |
32.5 |
38.9 |
|
DC. Dacrial (interorbital) chord |
23 |
20.5 |
|
DS. Dacrial subtense |
12 |
12.5 |
|
DS : DS. Dacrial index |
52.2 |
61.0 |
|
FC. Canine fossa depth (mm) |
5 |
4.5 |
|
Zygomatic curvature height (following Woo) |
7.5 |
11.5 |
|
Zygomatic breadth (following Woo) |
53.5 |
50.2 |
|
Zygomatic curvature index |
14 |
22.9 |
|
32. Frontal profile angle from nasion |
80 |
87 |
|
GM/FH. Frontal profile angle from glabella |
76 |
85 |
|
72. General facial angle |
83 |
82 |
|
73. Mid-facial angle |
86 |
86 |
|
74. Alveolar angle |
72 |
67 |
|
75. Nasal bones inclination index |
52 |
53 |
|
75 (1). Nasal protrusion angle |
31 |
29 |
|
68 (1). Mandibular length from condyles |
104 |
114 |
Table 1 (end)
|
1 |
2 |
3 |
|
79. Mandibular ramus angle |
118 |
127 |
|
68. Mandibular length from angles |
77 |
82 |
|
70. Ramus height |
65 |
61 |
|
71а. Minimum ramus breadth |
31 |
33.5 |
|
65. Mandibular condylar width |
123 |
113 |
|
66. Mandibular angular width |
96.5 |
101.5 |
|
67. Mandibular anterior width |
44 |
48 |
|
69. Symphiseal height |
30.5 |
31.5 |
|
69 (1). Mandibular corpus height |
29 |
31 |
|
69 (3). Mandibular corpus breadth |
9 |
11 |
|
C*. Mental protrusion angle |
72 |
63 |
|
Cranial shape in the horizontal plane |
Ovoid |
Ellipsoid |
|
Cranial shape in the lateral plane |
Ellipsoid |
″ |
|
Cranial shape in the occipital plane |
Roof-like |
Roof-like |
|
Intercilium (following Martin. 1–6) |
3 |
1 |
|
Browridges (1–3) |
2 |
1 |
|
External occipital tuber (following Broca. 0–5) |
3 |
1 |
|
Mastoid process (1–3) |
1 |
2 |
|
Lower margin of the piriform aperture |
Anthr. |
Anthr. |
|
Anterior nasal spine (following Broca. 1–5) |
3 |
4 |
A spacing is observed between the upper central incisors. These teeth are asymmetrical: the crown of the left central incisor is bent in its basal part and shifted labially with respect to the root, while its cutting edge is inclined lingually. The marginal ridges of the lingual surface are very weakly pronounced in both central and lateral incisors (shoveling grade 1). The latter display a moderate reduction (grade 1). The upper canines display distal accessory ridges. The Carabelli cusp is present in the first upper molars, while the distal accessory cusps are absent. Reduction of the hypocone of the second molars is more strongly pronounced than in skeleton 1: 3+ in the right molar, and 4 in the left molar.
Shoveling is absent in the lower front teeth. The lower first premolars are not differentiated, the second display two cusps, with some rudimentary elements of the grooves separating the lingual cusp. The morphology of the first lower molars cannot be described, while the second molars are four-cusped and display an “X”-pattern. In the third molars, protostylids are present. The right third molar is four-cusped and displays an “X”-pattern.
Possible origin of the deceased
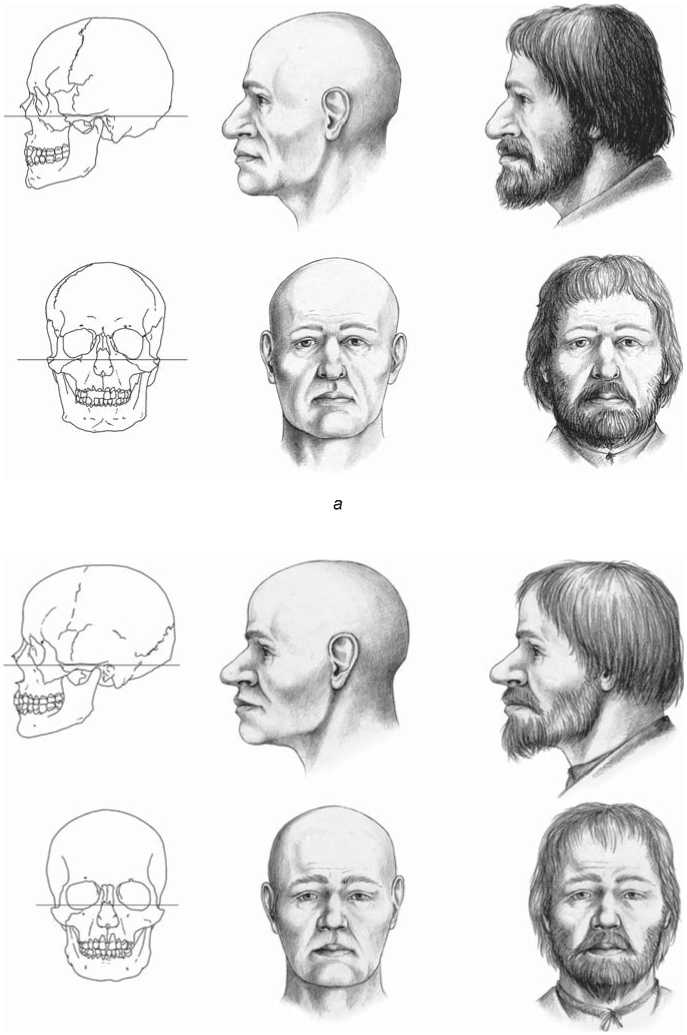
Main anthropological features of the studied individuals can be described on the basis of the morphological traits of the skeletons. First, both of them belong to the same anthropological type, while slight differences between them are explained by individual variation. This type displays a relatively small, low and meso-brachicranial vault, with smooth contours and moderate cranial robustness. The face is low, medium in width, and strongly protruding at both horizontal levels. The dimensions of the orbits and piriform aperture are medium. The nasal bridge and nasal bones are strongly protruding. Summing up, the morphological features of both individuals undoubtedly place them in the range of variation of the Causcasoid race. In our opinion, the closest analogs to this type are found in the samples of the Eastern Slavic population of the 17th to 19th centuries (Alekseev, 1969) (Fig. 2).
The dental pattern observed in the individuals is close to the maturized European dental complexes. It should be pointed out, however, that the possibility of individual diagnosis of dental taxonomic status in recent groups is limited. To determine this status, the following traits are important: presence of protostylid, enlarged cingulum of the lower molars, lingual inclination of the crowns of the upper incisors, and elements of the posterior fovea in the upper molars. But the protocol employed for the study and publication of the recent Ugrian, Russian, and Slavic population did not include these traits (see, e.g., (Vostochnye Slavyane…, 1999:
b
Fig. 2 . Graphic facial reconstruction of the deceased (author D.V. Pozdnyakov). a – skeleton 1; b – skeleton 2.
Tabl. XII-1–XII-3); thus, the area of origin of the studied individuals can be hypothesized only very tentatively. The morphological variants close to “steppe” types, with an increased frequency of four-cusped lower molars and a moderate reduction of the premolars, can be excluded from the list of potential ancestors of the Voikar individuals. The same applies to the Baltic dental type. The closest parallels to the studied specimens were observed in the medieval (11th to 13th centuries) dental samples from the Vologda Region: Chaigino-2, Volodino, Novinki (unpublished data of A.V. Zubova). The ethnic composition of those samples is complex. The cemetery of Chaygino is thought to belong to the Veps (Sankina, 2008: Tabl. 7); Volodino and Novinki as admixed, Slavic-Finnish and Slovensko-Krivich, respectively (Goncharova, 1995). An influence of similar complexes was also present, though much more weakly pronounced, in the 11th to 12th centuries sample from Staraya Ladoga (unpublished data of A.V. Zubova). Thus, to the best of our knowledge, all analogs to the specimens from Voikar are found in the Russian North, where both Slavic and Finno-Ugric elements are present.
Paleopathological characteristics and reconstruction of diet
The first important observation can be made is that there were no signs of peri-mortem, potentially lethal, trauma observed in the skeletons. The articular surfaces of the elbow, wrist, and knee joints of both individuals demonstrate changes due to pathological bone formation. These are “islands” of amorphous tissue that resemble, to some extent, markers of osteoarthritis (Fig. 3). A visual and X-ray examination of the long bones, as well as an assessment of their cross-section shape, revealed a thickening of the compact bone layer. In addition, well-pronounced lines of epiphyseal fusion and incomplete fusion of the acromion may point towards a delay in skeletal growth.
In the dentition of the deceased, numerous pathological manifestations are observed. Carious lesions were detected in the upper left third molar of skeleton 1, and the upper and lower third molars of skeleton 2. Lineal enamel hypoplasia (LEH) was observed in the upper and lower incisors, first premolars, lower canines, and second premolars of skeleton 1, and in the upper right canine of skeleton 2. Both individuals exhibit manifestations of periodontal disease: the roots of the teeth are naked for about one third of their length, and signs of a vascular reaction are observed in the alveoli and palate. Strong deposits of dental calculus are found, predominantly on the lingual surface of the premolars and molars and on the vestibular surface of the incisors, both upper and lower. The upper right first incisor and lower left molars of skeleton 2 display enamel defects. The lower left first and third molars were lost ante-mortem. The lower right first molar exhibits manifestation of a probable defect of dentinogenesis: its occlusal surface demonstrates a bowl-shaped deepening without any external relief. The deepening reaches the level of the tooth’s neck, but is covered with a smooth layer of enamel. The non-carious nature of this pathology was confirmed by a sounding, which demonstrated that the root canals were closed. As the left first molar was lost ante-mortem, it was not possible to determine if the pathology was symmetrical.
The suite of traits observed in the skeleton and dentition of both individuals suggests that they might have experienced deficiencies of vitamin C and phosphorus during their childhood and adolescence (Ortner, Ericksen,

Fig. 3 . Manifestations of pathological bone formation in the right radius (skeleton 1).
1997). The pattern of dental pathologies points towards a diet rich in carbohydrates, and generally towards a high level of biological stress in the population to which the deceased belonged. The latter suggestion is based on the presence of LEH—the marker of episodic stress of various etiologies: famine, parasitic invasions, hereditary diseases, etc.
Postcranial metrics and reconstruction of physical activity
The full and articular (functional) lengths of the long bones of both individuals are medium or below medium, while the lengths of the tibia display small values (Table 2). The robustness index is high. The diaphyses of the humerus and ulna are rounded in a transverse section, and the radius displays a flattened section. The section through the femur of skeleton 1 at the level of diaphysis is round, while in skeleton 2 it is flattened.
The radio-humeral index in skeleton 1 points towards an equal ratio between the lengths of the two bones, while the femur is long relative to the tibia. Indices show that the humerus is slightly shortened in respect to the humer, while the forearm demonstrates a substantial elongation relative to the tibia. According to the intermembral index, the body proportions of skeleton 1 can be described as brachymorphic. The radio-humeral index in skeleton 2 suggests a relative elongation of the forearm, while the humero-femoral index shows a substantial elongation of the humerus relative to the femur.
The stature of the individuals was estimated using various formulae. For skeleton 1, these estimates are 168.5 cm (according to M. Trotter and G. Gleser), 167.7 cm (According to E. Breitinger), and 166.4 cm (according to M. Cherny and S. Komenda); and for skeleton 2 these values are 159.7; 162.4, and 158.8 cm (Pezhemsky, 2011: 299–307).
Table 2. Postcranial metrics of individuals 1 and 2
|
Variable |
1 |
2 |
|
1 |
2 |
3 |
|
Humerus |
||
|
1. Maximum length |
322/320* |
313/… |
|
2. Total length |
319/317 |
305/303 |
|
3. Proximal epiphyseal breadth |
54/56 |
55/55 |
|
4. Distal epiphyseal breadth |
68/67 |
63/… |
|
5. Maximum midshaft diameter |
23/22.7 |
21.5/21.5 |
|
6. Minimum midshaft diameter |
19/19 |
18/19 |
|
7. Minimum circumference |
65/67 |
65/66 |
|
7а. Midshaft circumference |
73/71.5 |
67/67 |
|
8. Head circumference |
…/153 |
|
|
9. Maximum head diameter |
…/46 |
|
|
10. Vertical head diameter |
48/… |
|
|
Radius |
||
|
1. Maximum length |
246/247 |
231/… |
|
2. Physiological length |
229/… |
220/… |
|
4. Transverse midshaft diameter |
17/18 |
16.5/17 |
|
5. Sagittal midshaft diameter |
11/12 |
11.5/12 |
|
3. Minimum midshaft circumference |
45/45 |
43/43 |
|
Ulna |
||
|
1. Maximum length |
267/272 |
259/… |
|
2. Physiological length |
230/234 |
223/220 |
|
11. Antero-posterior midshaft diameter |
13/14 |
13.5/14 |
|
12. Transverse diameter |
19/19 |
16/17.5 |
|
13. Upper transverse diameter |
22/21 |
15.5/… |
|
14. Upper sagittal diaphyseal diameter |
25.3/24.5 |
22.5/… |
|
3. Minimum diaphyseal circumference |
38/41 |
37/37 |
|
Clavicle |
||
|
1. Maximum length |
144/141 |
139/142 |
|
6. Midshaft circumference |
40/40 |
38/37 |
|
Scapula |
||
|
1. Morphological breadth |
153/156 |
167/167 |
|
2. Morphological length |
102/104 |
93/95 |
|
Femur |
||
|
1. Maximum length |
…/446 |
416/414 |
|
2. Natural length |
…/444 |
410/404 |
|
21. Condylar breadth |
…/86 |
85/81 |
|
6. Midshaft sagittal diameter |
…/26 |
24.5/25 |
|
7. Midshaft transverse diameter |
…/25 |
26.5/26.5 |
|
9. Upper transverse diaphyseal diameter |
…/29 |
29/28.5 |
|
10. Upper sagittal diaphyseal diameter |
…/23.5 |
24/24 |
|
8. Midshaft circumference |
…/81 |
82/82 |
Table 2 (end)
|
1 |
2 |
3 |
|
Tibia |
||
|
1. Total length |
343/342 |
|
|
2. Condylo-talar length |
…/322 |
296/… |
|
1а. Maximum length |
350/351 |
|
|
3. Proximal epiphyseal breadth |
75/79 |
78/… |
|
6. Distal epiphyseal breadth |
56/… |
|
|
8. Midshaft sagittal diameter |
24/26 |
25/… |
|
8а. Sagittal diameter at for. nutr. |
29.5/27 |
30/… |
|
9. Midshaft transverse diameter |
19.5/20.4 |
23/… |
|
9а. Transverse diameter at for. nutr. |
22/22 |
26/… |
|
10. Midshaft circumference |
71/75 |
79/… |
|
10b. Minimum diaphyseal circumference |
66/… |
|
|
Indices |
||
|
Intermembral |
…/72.14 |
|
|
Tibiofemoral |
…/77.03 |
|
|
Radiohumeral |
76.40/77.19 |
73.80/… |
|
Humerofemoral |
…/72.07 |
76.34/… |
|
Radiotibial |
71.72/72.22 |
|
*Right side/left side.
Markers of physical activity are similar between the two individuals: the forelimb and its girdle experienced much higher loadings than the hindlimb. Entheseal changes in the form of enthesopathy and diaphyseal shape deformations are particularly evident in the sites of attachment of M. deltoideus, teres major , and m. latissimus . Entheseal markers are also developed at the sites of the forearm bones: m. biceps , pronators and supinators, flexors of the hand and fingers. An analysis of most stereotypic movements of the forelimbs points towards a frequent forceful flexing of the forearm and hand, and extensions and adductions of the humerus, which can be interpreted as an activity aimed at drawing an object to the trunk. The most typical movement for both individuals was a retraction of the prone shoulder backwards followed by an adduction of the shoulder. Such a suggestion is supported by a relocation of the articular surfaces of the shoulder joint in dorsolateral direction, and by the presence of an additional angle of the scapula (Fig. 4, 5). The latter might be explained by permanent loadings to m. teres major , which is mainly involved in the types of physical activity described above.
On the basis of the analysis of entheseal and articular surface changes, the stereotypic motions can be integrated into activity complexes. The pathological manifestations in the shoulder joint described above are interpreted in some studies as a result of elevating and moving heavy objects. On the other hand, movements of the legs were mostly aimed at keeping a static position of the body, its bending and extension, as well as to flexion and extension of the foot. Importantly, the femoral muscles of the anterior, posterior, and lateral groups were moderately or weakly developed. This means that regularly carrying heavy weights is not a likely type of activity for the studied individuals, as such an activity implies a substantial loading on the hindlimb muscles.
Besides the activity patterns discussed above, there are traces of dynamic loadings requiring simultaneous work from the pectoral and dorsal muscles, particularly those involved in contraction and extension of the scapula. Thus, the shoulder joint was performing a rotation accompanied by bending and unbending in the elbow joint. As a similar complex of activity markers is observed in modern academic rowers (Smirnov, Dubrovsky, 2002: 529–532), we hypothesize that the two studied individuals rowed regularly. Judging from the observations made in the Sadlermiut (an isolated group of the Paleoeskimo) and the Thule Paleoeskimo samples, Hawkey and Merbs described the following set of features associated with rowing: arthritis of the acromioclavicular, humeroulnar, and humeroradial joints, and left-sided injuries of the joint between the ulna and metacarpal bones (Merbs, 1983: 68–72;

Fig. 4 . Accessory articular surface of the right humerus (skeleton 2).
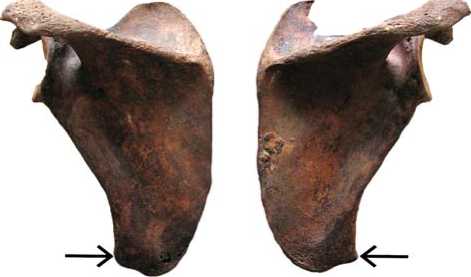
Fig. 5 . Accessory scapular angle at the m. teres major attachment site (skeleton 2).
Hawkey, Merbs, 1995: 329–334). These are in only a partial agreement with the features observed in the present studies. This can be explained by the fact that the populations studied by Hawkey and Merbs were using kayaks with only one light paddle for their aquatic travels.
This is not what was typical for the Voikar individuals, for whom another type of rowing can be suggested: namely the use of vessels with heavy rowing-equipment, where the main loadings fell on the pectoral and dorsal muscles, while the shoulder and forearm muscles were only indirectly involved. Elaborating the hypothesis on the occupational activity of the individuals from Voikar as shipmen, it is worth noting that not only were heavy paddles used by the Russians in the Ob basin during the 17th to 18th centuries for navigating river vessels of various types, but also the rig and string draft were employed. Usual carrying of the vessels across the ground should not be overlooked either (Vershinin, 2001: 90, 92, 97, 99, 103, 104). These probable occupational factors may indirectly explain the presence of markers of extreme physical loadings observed in the skeletons: relocation of the articular surface of the humerus, and substantial development of the iliopsoas muscles and m. glutei maximi that bend and unbend the body (Fig. 6). Another very feasible type of physical activity might be dragging motions associated with the extraction of fishing nets. Archaeological data suggest a substantial role for fishing, including trawling, in the lifestyle of the Russian population of the northwestern Siberia (Vizgalov, Parkhimovich, 2008: 110–111).
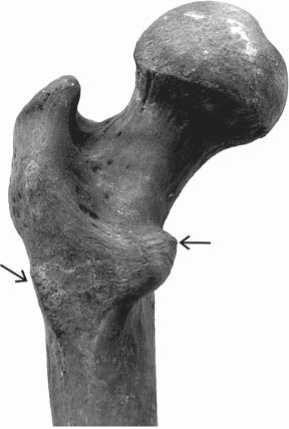
Fig. 6 . Entheseal changes at the m. gluteus maximus and m. iliopsoas attachment sites (skeleton 1).
Discussion
All the results obtained in the present study suggest that the intramural burial found at the North Siberian aboriginal settlement contained the remains of newcomers, probably of Eastern Slavic origin. The Eastern Slavs have been following Christian burial traditions, notably unified across Siberia in the 16th to 18th centuries, according to archaeological data (Tataurova, 2010: 28, 42). But the people who made the inhumation at Voikar were not familiar with the Christian burial tradition, or did not find necessary to follow it.
Were examples of such unusual inhumations are known in the burial practice of native peoples of the region? The very scarce archaeological data on the burial rites of the medieval population of the Lower Ob region are dated mostly to the 6th to 13th centuries, and afterwards there is a chronological gap until the 19th century. The burial complexes are represented by flatgrave burial grounds, where individual inhumations with grave constructions of various types were predominant. The deceased were typically in an extended supine position, with the head oriented to southwest, south, or southeast (see, e.g., (Zeleniy Yar, 2005: 69–70, 143– 149; Gusev, 2016). Such a position of the deceased was as typical in the late 19th century as well (Murashko, Krenke, 2001: 29–30). Medieval burials with a flexed position of the body are found very rarely and mostly in the tundra zone of the Yamal Peninsula (cemeteries of Kheto-Se-1, Bukhta Nakhodka-2, Yur-Yakha III) (Brusnitsyna, 2000: 37; Kardash, Gaydakova, 2017; Plekhanov, 2016). The single burial studied by V.N. Chernetsov at Khaen-Sale settlement, on the northeast coast of the peninsula, probably belongs to a later period: according to the grave goods it was attributed to the 16th century AD (1957: 236). Researchers point to some similarity between medieval cemeteries in the north of the Lower Ob region and the chronologically close burial complexes to the south of them, in the taiga zone of the Ob region (Zeleniy Yar, 2005: 288). Besides these common features, it is of note that double or collective burials were exceptional finds in this region (for the Surgut region of the Ob see, e.g., (Zykov, 2012: 84, 96, 104)).
Turning to the tradition of intramural burials, it should be noted that in Western Siberia such burials are found mostly in the forest-steppe zone during the Bronze Age and transition to the Iron Age; and even in that period, these were extraordinary (Novikova, 2011). Some cases have been observed in the taiga region in medieval times as well (Adamov, Turova, 2003; Kazymskiy… kompleks, 2018: 137–138).
Only single finds of human remains in dwellings are known for the northern part of Western Siberia in medieval and recent times. These can be divided into three groups. The first groups includes individuals who died under ruins of dwellings (Fort Nadym (Kardash, 2009: 30)). The second group includes intentional burials in dwellings demolished before making a burial (Fort Monkys Uriy (Kardash, Vizgalov, 2015: 316–329)). In both groups, the deceased were victims of armed attacks on settlements. The last group includes intentional burials in dwellings that were, according to the researchers, inhabited at the moment of creating a burial. The burials are found in different parts of the dwellings. This group includes inhumations at Khaen-sale (Chernetsov, 1957: 236) and Barsova Gora IV/26 (Surgut region of the Ob) settlements (Beltikova, 2002).
To the best of our knowledge, the only case (except the one described in the present study) in the Lower Ob region where the space of the hearth was used for ritual manipulations with human remains was also described for the Ust-Voikar settlement. In the hearth of one of the dwellings built, according to dendrochronological data (Gurskaya, 2008: 221–222) in the early 14th century, a human scalp with remains of hair was found (Etnicheskaya arkhitektura…, 2008: 48).
Thus, the burial at Voikar is completely atypical for both Christian burial rites and the traditions of the native population of the Lower Ob region. The interpretation of its semantic status in the system of beliefs of the native population of the northwestern Siberia requires further investigation.
Conclusions
The results of a complex anthropological study of the human remains found in a hearth of a dwelling at Ust-Voikar lead to several conclusions. First, the remains belonged to two adult males ( adultus – early maturus ), likely belonging to the same population. This population was probably related to the population of the north of the East European plain. Life conditions of this ancestral population were not advantageous, as suggested by the manifestations of childhood biological stresses, including vitamin C and phosphorous deficiencies, observed in both individuals.
The habitual diet of the deceased was rich in carbohydrates. Our reconstruction of the stereotypical motions of the individuals points towards a regular use of river vessels equipped with heavy paddles, and fishing as a part of common occupational activity. The cause of their death cannot be determined by bioarchaeological methods, though the absence of peri-mortem trauma in the skeletons might suggest that they were not victims of a military encounter.
The Lower Ob region has been, for a while, a route of dispersion of the East European population into Siberia, while northern routes across the Urals (“over the Stone”) were widely used during the 17th century as well (Bakhrushin, 1928: 68; Perevalova, 2004: 33; Vershinin, 2018: 65–66). However, it does not seem possible to determine exactly what historical events of the last third of the 17th to early 18th centuries led to the inhumation of representatives of the East European population in the territory of an inhabited aboriginal settlement. We can only suggest that this unusual event was somehow related to the process of incorporation of Western Siberia into the Russian state.
