Адаптационная модель потери губчатой кости при старении

Автор: Аврунин А.С., Тихилов P.M., Паршин Л.К., Шубняков И.И.
Журнал: Гений ортопедии @geniy-ortopedii
Рубрика: Оригинальные статьи
Статья в выпуске: 1, 2007 года.
Бесплатный доступ
На основе анализа данных о влиянии старения на процессы моделирования/ремоделирования и механические свойства костных структур теоретически обоснована и разработана этиопатогенетическая модель адаптационного механизма потери губчатой кости при старении. Выделены следующие пути профилактики возникновения остеопоротических переломов: разработка методов укрепления мышечной системы для сохранения величины механического сигнала у лиц с высоким риском развития остеопороза (низкая пиковая костная масса, хроническая патология и т. д.), начиная в соответствии с принципами профилактической медицины с 30-летнего возраста, то есть с момента начала потери костной массы, а не после возникновения нарушения архитектуры губчатой кости; создание медикаментозных препаратов для профилактики потери костной массы, обеспечивающих: 1) уменьшение минерализации костной ткани с помощью регуляторов остеоцитарного насоса для обеспечения вымывания Са++ из костной ткани; 2) снижение порога инициации электрических импульсов в костных клетках, что позволит оптимизировать процессы ремоделирования на более низком уровне деформаций костной ткани при изменении механических нагрузок
Остеопороз, адаптационная модель, трабекулярная кость, профилактика остеопороза
Короткий адрес: https://sciup.org/142120958
IDR: 142120958
The adaptation model of cancellous bone loss for aging
The etiopathogenetic model of adaptation mechanism of cancellous bone loss with aging has been theoretically substantiated and developed on the basis of analyzing the data of the aging effect on the processes of modeling/remodeling and the mechanical properties of bone structures. The following ways of osteoporotic fracture prevention have been highlighted: working-out the methods of muscular system strengthening to preserve the quantity of mechanical signal in subjects with high risk of osteoporosis (low peak bone mass, chronic pathology, etc.), starting from 30-year age according to the principles of preventive medicine, that means from the moment when bone mass loss begins, but not after the moment when cancellous bone architecture has already been disturbed; production of medicaments to prevent bone mass loss, which would make available: 1) decrease of bone tissue mineralization with the help of osteocytic pump regulators for Ca++ washing out from bone tissue; 2) decrease of the threshold of electric impulse initiation in bone cells, that will give the possibility to optimize remodeling processes at a below level of bone tissue deformities in case of changing mechanical loads.
Текст научной статьи Адаптационная модель потери губчатой кости при старении
После 25-летнего возраста независимо от пола начинается потеря костной массы со средней скоростью 1 % в год. У женщин в период утраты репродуктивной функции в течение первых 5-7 лет она достигает 2-3 % в год [31]. В результате к 70 годам масса губчатой кости сокращается на 60 % [17]. Этот процесс развития остеопороза или «безмолвной болезни» асимптоматичен до возникновения переломов [52], которые заставляют рассматривать его как одну из важнейших нерешенных проблем здоровья общества, характеризующуюся быстрой эскалацией социальных и медицинских затрат [55]. Только в США в результате потери губчатой кости ежегодно насчитывается 700000 переломов позвонков и 250000 – дистального отдела лучевой кости, а прямые затраты, связанные только с лечением всех типов остеопоротических переломов составляют 10 млрд. долларов [20], в целом же достигая 20,1 млрд. [47].
Общепринятая практика рассмотрения пере- лома как одного из основополагающих критериев диагностики остеопороза, по мнению Nordin, с патогенетической точки зрения ошибочна [49]. Остеопороз только увеличивает риск перелома, не являясь его причиной. В связи с этим на первый план выходит необходимость объяснить патогенетические причины возникновения переломов только в 2 % случаев от общего числа однотипных падений на бедро [47].
Минеральная плотность, геометрия, микроархитектура и свойства материала кости – все это компоненты, определяющие ее прочность, то есть способность противостоять нагрузке. Однако до настоящего времени комплексная проблема "качества" кости осмыслена недостаточно [55]. Это связано с тем, что эффекты старения скелета проявляются по мере накопления сдвигов на всех уровнях его иерархической организации и являются по сути лишь структурным отражением возрастных регуляторно-метаболических отклонений [6]. С этой точки зрения систематизация данных об ультраструктурных изменениях по- зволит теоретически обосновать и разработать принципиально новые направления профилактики остеопоротических переломов, включая создание медикаментозных препаратов. По мнению W.C. Hayes и E.R. Myers [41], такая необходимость обусловлена тем, что для предотвращения перелома при падении на бедро у больных остеопорозом необходимо увеличить проекционную минеральную плотность костной ткани проксимальной части бедренной кости в среднем более чем на 20 %. По данным же контролируемых фармакологических исследований, в этом отделе скелета ее удается повысить в лучшем случае на несколько процентов. Особенно незначительна динамика у пожилых пациентов с наибольшим риском перелома.
В настоящее время среди концепций патогенеза потери губчатой кости при старении доминируют взаимодополняющие друг друга теории, связанные с деятельностью остеокластов [32].
После утраты репродуктивной функции наблюдается увеличение глубины остеокластной резорбции, происходящей с одной или обеих сторон трабекулы, что приводит к ее перфорации при нормальной толщине.
Остеокласты преимущественно «атакуют» более тонкие пластины.
Происходит генерализованное истончение трабекул, делающее их более уязвимыми к разрушению при формировании лакун резорбции нормальной глубины.
Вероятность перфорации трабекул возрастает вследствие повышения частоты активации единиц ремоделирования1 после менопаузы.
Цель исследования . Проанализировать влияние старения на процессы моделирова-ния/ремоделирования2, на механические свойства костных структур, теоретически обосновать этио-патогенетический механизм потери губчатой кости при старении, предложить принципиально новые направления профилактики возникновения остеопоротических переломов.
-
I. ВЗАИМОСВЯЗЬ МЕХАНИЧЕСКИХ НАГРУЗОК И СТРУКТУРНОЙ ГЕОМЕТРИИ КОСТНЫХ ОРГАНОВ
Структурная геометрия на всех уровнях скелетной иерархии является результирующей процессов моделирования/ремоделирования [6, 45]. Согласно доминирующей в современной остеологии парадигме Юта, изменение механи- ческих напряжений регламентирует активацию и пространственное распределение участков синтеза и резорбции костной ткани, а гуморальные (гормоны, цитокины и др.) и структурные (внеклеточный матрикс) регуляторы служат фоном, определяющим особенности взаимосвязанного функционирования костных клеток [2, 15, 39, 40]. Изменение напряжений происходит в процессе повторяющихся сокращений мышц, обеспечивающих движение органов скелета (ригидных рычагов) против сил гравитации.
Трабекулы формируют решетку, образуя дуги опоры, синхронизированные с основными линиями передачи физиологических нагрузок [30] от суставных поверхностей к диафизу. Нагрузки вызывают локальные деформации внеклеточного матрикса, воспринимаемые костными клетками как механические сигналы, запускающие процесс механотрансдукции.
-
II. МЕХАНОТРАНСДУКЦИЯ
Преобразование механических сигналов в биохимические имеет следующие основные этапы [57]:
преобразование локальных механических деформаций внеклеточного матрикса, возникающих при изменении нагрузки, в локальные механические сигналы, инициирующие ответ костных клеток;
трансдукция механических сигналов в биохимические и электрические с активацией метаболических путей в пределах мембраны клеток и цитоскелета;
передача сигналов «клетка – клетке» от клеток датчика (остеоциты и клетки пограничного слоя) к клеткам исполнительного элемента (остеобласты или остеокласты);
ответ исполнительного элемента – формирование или резорбция костной ткани, обеспечивающие необходимые архитектурные преобразования.
На первых двух этапах инициация механотрансдукции регламентируется:
механическими свойствами внеклеточного матрикса, определяющими величину его деформации под влиянием механических нагрузок;
порогом чувствительности клеток к механической деформации внеклеточного матрикса при нагрузке;
характером и силой механических нагрузок на органы скелета.
Рассмотрим влияние старения на функционирование этих регламентирующих механизмов.
-
III. ВЛИЯНИЕ СТАРЕНИЯ НА МЕХАНИЧЕСКИЕ СВОЙСТВА ИЕРАРХИЧЕСКИ ОРГАНИЗОВАННЫХ СТРУКТУР СКЕЛЕТА
Изменение механических свойств кости при старении вызвано накоплением структурных сдвигов, формирующихся на всех уровнях орга- низации скелета, поэтому первоначально рассмотрим иерархию его структуры.
Иерархическая организация скелета . В настоящее время выделяют 7 морфоструктурных уровней [2, 15, 22, 23, 59]:
молекулярный;
надмолекулярный – объединение органических молекул в надмолекулярные комплексы с включением в их состав минералов;
тканевой – объединение надмолекулярных комплексов в костные тканевые ретикулофиброз-ные и пластинчатые структуры костной ткани;
структурно-функциональный – объединение костных пластинок в остеоны и трабекулы (полуостеоны);
уровень органообразующих структур – объединение остеонов и полуостеонов в кортикальный слой (компактное вещество) и губчатое вещество;
органный – объединение компактного и губчатого вещества в едином костном органе;
суперорганный – объединение всех костных органов в единый комплекс, т. е. скелет.
Влияние старения на ультраструктурные характеристики, определяющие механические свойства костной ткани 3 . Изменение механических свойств нарастает по мере накопления ультраструктурных преобразований внеклеточного матрикса [1, 22, 26, 59], в том числе, увеличения доли минерального компонента и снижения водной и органической фракции [22, 43, 51, 53, 54], уменьшения межмолекулярных пространств [50, 60]. Рассмотрим более детально происходящие изменения и механизмы, определяющие их возникновение.
Возрастзависимые изменения на молекулярном, надмолекулярном и тканевом уровнях. При старении изменяются связи и геометрия коллагеновых фибрилл и волокон, а также их ориентация и взаимодействие между коллагеном и минералами [59]. Эти сдвиги возникают при ремоделировании как функция возраста и связаны с денатурацией коллагеновых молекул и образованием неферментативных перекрестных сшивок коллагена в период формирования остеоида, сказывающихся на структуре молекул, ограничивающих межмолекулярные пространства и соответственно меняющих ультраструктуру костной ткани. В каждом цикле ремоделирования доля трансформированных молекул незначительна, но по мере старения сдвиги накапливаются и достигают критического уровня при наличии нарушений в 30 % и более молекул колла- гена. Результатом является изменение механических свойств [60], в том числе увеличение жесткости органического матрикса и соответственно уменьшение величины деформаций.
В участках ремоделирования ультраструктура остеоида меняется и в результате возрастзависи-мого сдвига в спектре синтезируемых остеобластами молекул [1]. Так, например, изменяется экспрессия остеобластами мРНК коллагена I типа, остеопонтина и остеокальцина [26]. В связи с тем, что остеоид формируется в процессе самосборки после секреции костными клетками его компонентов, необходимо учитывать также тот факт, что возрастные регуляторно-метаболические сдвиги, как было показано выше, влияют на ферментную и неферментную модификации их молекулярной структуры [1]. То же относится и к структуре минерального матрикса, так как кристалличность4, стехиометрия и количество минеральных кристаллов кости зависят от состояния термодинамического и кинетического равновесий, определяемых физиологической средой [22].
Отложение минерального матрикса начинается через 8 суток после образования остеоида [36]. В конечном итоге формируется сложная 4уровневая минеральная структура, являющаяся в каждом участке запрограммированным отражением органической ультраструктуры [5, 7]. При этом по мере старения степень кристалличности минералов увеличивается на 1-2 % [22], что происходит под действием остеоцитарного насоса, попеременно нагнетающего в костную ткань минералобразующие ионы и экскретирующего их из нее. В результате происходит частичное растворение или осаждение минералов. В первом случае, в первую очередь, удаляются ионы, входящие в состав аморфной фазы, так как энергия отрыва иона от кристаллической решетки существенно выше. Во втором часть ионов, оказавшихся в более устойчивом состоянии, формирует кристаллическую решетку, и в дальнейшем вероятность их отрыва при растворении минерала оказывается намного ниже [3]. По-видимому, повышение кристалличности отражает увеличение собственного возраста костной ткани, то есть большую длительность действия остеоцитарного насоса.
Как известно, в костной ткани гидроксиапатит находится в метастабильном состоянии в виде коллоидных кристаллов, радиус частиц которых варьирует от R1 до R2 [3, 19]. Увеличение кристалличности приводит к снижению растворимости минералов [22] и, соответственно, к росту их размеров, то есть возрастанию числа минералов, габариты которых находятся на верхней границе (R2). Это подтверждают данные Ю.И. Денисова-Никольского с соавторами [2002], которые установили нарастание разме- ров копланарных объединений кристаллов5 (длины на 17–20 % и ширины на 5–9 % при неизменной толщине) [11]. Расчеты показывают, что это, в свою очередь, влечет за собой рост объема этих структур на 23–31 % [16] с вытеснением водной [51, 54] и органической [53,54] фракций из внеклеточного матрикса.
Вышеизложенные сдвиги обусловливают уменьшение межмолекулярных промежутков и пространств, в которых интегрирована остеоци-тарная сеть. Так, если в период роста и развития организма канальцы короткие и широкие, а в зрелом возрасте они становятся более разветвленными, то при старении их диаметр и количество снижается [50].
При старении минерализация матрикса увеличивается более чем на 12 % [22]. После 57 лет доля остеонов, минерализованных на 75 % и выше, составляет более 40 % от общего их числа, в то время как в более молодом возрасте она не превышает 10 % [43].
По мере старения, кроме увеличения кристалличности, на 10–17 % возрастает доля нестехиометрических замен ионов фосфата на ионы карбоната в кристаллической решетке [13, 22]. Эти замены вызваны гетероионным обме-ном6 и, по сути, отражают возрастные сдвиги метаболизма [5, 7].
Описанные выше изменения размера и формы кристаллов, ухудшение симметрии кристаллической решетки и наличие вакансий в ее пределах стимулируют возникновение остаточных напряжений в соседних минералах и молекулах коллагена. В условиях подобной трансформации механические нагрузки могут индуцировать дефекты в ультраструктуре кости, а увеличение степени минерализации приводит к уменьшению подвижности фибрилл коллагена. В результате наблюдается существенное снижение величины деформаций внеклеточного матрикса под действием одной и той же механической нагрузки [22].
Следовательно, можно утверждать, что сдвиги, происходящие при старении на первых трех уровнях организации костной ткани, снижают способность кости к деформированию при той же самой нагрузке и соответственно величину механических сигналов. Рассмотрим теперь изменения, возникающие на более высоких уровнях организации кости в процессе старения.
Возрастзависимые сдвиги на структурнофункциональном и последующих иерархических уровнях характеризуются в числе прочего истончением трабекул со средней скоростью 1 мкм или (-0,6 %) толщины в год [32], и при снижении ее средней величины до 120 µм и меньше в проксимальном отделе бедренной кости человека трабекулярная кость считается остеопоротической (в норме средняя длина трабекулы 1 мм, толщина 283 µм) [30].
Одновременно с истончением трабекул увеличивается число единиц ремоделирования, прекращающих свое развитие после фазы реверсии7, что ведет к образованию пустых лакун резорбции. Появление этих полостей с двух сторон пластины при остеопорозе ослабляет структуру, повышая вероятность ее разрушения [32], так как часть трабекул, особенно поперечных, отсутствует и не компенсирует нагрузку [42]. Поперечные трабекулы препятствуют потере устойчивости сжатых продольных трабекул (в форме изгиба). При исчезновении поперечных поддержек увеличивается протяженность продольной трабекулы, а формирование лакун с обеих ее сторон приводит к разрушению при нагрузке. В результате, как подчеркивает Dempster [32], происходит нарушение архитектуры, особенно в постменопаузальном периоде. Трабекулы становятся более разобщенными, а трабекулярная сцепленность снижается. В конце концов локально могут сохраниться лишь несколько продольных трабекул и беспорядочно разбросанных поперечных [42].
Рассмотрим теперь, как же в результате всех этих процессов изменяются механические свойства губчатой костной ткани при старении.
Влияние старения на механические характеристики губчатой кости . Объективная оценка механических свойств трабекул (тонких, неровных, миниатюрных объектов) технически крайне сложна, так как существенное влияние на результаты расчетов оказывают все виды нарушений их габаритов. Поэтому при исследовании больших фрагментов трабекулярной кости получают, по существу, кажущиеся показатели, зависящие от особенностей архитектуры исследуемого фрагмента (см. табл.). Компьютерное моделирование позволяет получить лишь очень приблизительные оценки механических характеристик материала трабекул [29].
Таблица Механические свойства трабекулярной кости человека по сводным данным литературы [46]
|
Свойства |
Решетчатая кость |
|
Предел прочности при сжатии (MPa) |
0,5-50 |
|
Предел прочности при растяжении (MPa) |
10-20 |
|
Прочность на изгиб (MPa) |
2-25 |
|
Модуль Юнга (GPa) |
0,05-1 |
В связи с отсутствием данных о существенных ультраструктурных различиях между губчатым и компактным веществами кости можно утверждать, что их механические свойства на уровне межкле- точного вещества существенно не отличаются. Аналогичного мнения придерживаются А. C. Lawson [46] и J. D. Currey [29], тем более что дифференциация на компактное и губчатое вещества проводится на основе такого произвольно выбранного критерия, как степень пористости (от 5 % до 30 % и от 31 % и более соответственно) [27].
Отсутствие различий подтверждается исследованием микротвердости трабекул, согласно которому она лишь незначительно меньше, чем в близлежащей кортикальной кости и в обеих зонах изменяется пропорционально степени минерализации ткани, определенной микрорентгенографически [61]. Незначительная разница может быть обусловлена следующими факторами [29]:
минеральное содержание решетчатой кости несколько меньше, чем соседней компактной;
ориентация пластинок и кристаллов соответствует трабекулярным линиям, но организована хуже, чем в компактной кости.
Нарастающая возрастзависимая минерализация внеклеточного матрикса и образование дополнительных поперечных связей в коллагеновых фибриллах усиливает его жесткость и снижают степень деформации при одной и той же нагрузке. По данным van der Linden J. C. с соавторами [58] увеличение минерализации ткани кости на 5-10 % приводит к повышению ее ригидности на 16-33 % и возрастанию модуля упругости. В результате, при прочих равных условиях, уменьшается относительная деформация, т. е. механический сигнал, определяющий характер ремоделирования.
В связи с вышеизложенным при оценке физиологической роли механических характеристик необходимо выделить две составляющие. Первая – обеспечение прочностных свойств трабекулярной структуры, определяющих ее несущую способность, которая преимущественно зависит от особенностей ее архитектуры и степени нарушений последней, и вторая – механические характеристики жесткости собственно межклеточного матрикса, детерминирующие его деформируемость при нагрузке и, следова- тельно, интенсивность механического сигнала.
Величина относительной деформации растяжения или сжатия трабекулы согласно закону Гука [8] составляет:
F
EA ,
где E – модуль упругости, А – площадь поперечного сечения трабекулы, F – приложенная к трабекуле сила.
Согласно этому закону величина деформаций обратно пропорциональна площади поперечного сечения, а, как известно, при старении толщина трабекул уменьшается со скоростью -0,6 % в год. Для простоты расчетов представим себе поперечное сечение трабекулы в виде круга, тогда А=R22, и при среднем диаметре трабе- кул 283 µm получаем площадь поперечного сечения 62870 µм2. Поскольку радиус трабекулы уменьшается на -0,6 % в год, за 40 лет (≈ к 70 годам) он уменьшится на 34 µм, и площадь поперечного сечения станет равной 36287 µм2, то есть сократиться в 1,7 раза. Исходя из этого, величина деформаций, казалось бы, должна увеличиться аналогичным образом. Однако, как отмечено выше, костная ткань при старении становится более минерализованной и соответственно ригидной. Следовательно, истончение трабекул можно рассматривать как результат адаптационных изменений, направленных на сохранение величины деформаций (то есть величины механического сигнала) в ответ на увеличение жесткости внеклеточного матрикса.
Однако реакция остеоцитов на механический сигнал определяется не только величиной относительных деформаций, но и порогом механо-сенсорной чувствительности клеток.
-
IV. ВЛИЯНИЕ СТАРЕНИЯ ОРГАНИЗМА НА ЧУВСТВИТЕЛЬНОСТЬ КОСТНЫХ КЛЕТОК К МЕХАНИЧЕСКИМ СИГНАЛАМ
Устойчивость организма к воздействию факторов внешней среды определяется возможностью сохранения параметров гомеостаза в пределах их нижней и верхней границ [9, 14, 21]. В костной ткани межпороговый промежуток или «ленивая зона» – это тот интервал, в котором костные клетки не отвечают на изменение на-пряжений/деформаций изменением архитектуры [58]. Другими словами, при данной ультраструктуре ткани в условиях физиологической нагрузки в этом интервале величина деформаций внеклеточного матрикса является оптимальной для метаболизма костных клеток.
По экспериментальным данным C.H. Turner с соавторами [56], у старых крыс на эндостальной поверхности формирование кости увеличивается только при достижении нагрузки в 64 N, при более низких ее значениях, в отличие от более молодых животных, реакция отсутствует. Согласно расчетам этих авторов на эндостальной поверхности у молодых крыс механический порог деформаций для формирования пластинчатой кости составил 1050 μs , а у старых – более 1700 μs. [56].
Таким образом, впервые показано снижение при старении чувствительности костных клеток к механическим стимулам. По сути, эти данные можно рассматривать как подтверждение общебиологической закономерности возрастзависи-мого изменения порогов рецепторной чувствительности к различного рода сигналам. Например, как отмечает В. М. Дильман [1987], механизм выключения репродуктивной функции при старении организма осуществляется за счет постепенного повышения порога чувствительности тонического центра гипоталамуса к регуляторным сигналам (эстрогенам) [12].
Еще одним регламентирующим фактором начального этапа процесса механотрансдукции являются сила и характер механических нагрузок, определяемых, в первую очередь, функцией мышц.
-
V. ВОЗРАСТНОЕ ИЗМЕНЕНИЕ МЕХАНИЧЕСКИХ НАГРУЗОК
В процессе старения уменьшаются фактические нагрузки на скелет, так как снижается повседневная активность и сила мышц [58]. Это связано с уменьшением размеров и количества мышечных волокон, длины мышечных пучков, фибрилл II типа и силы по отношению к площади поперечного сечения [24]. Изменение двигательной функции при старении характеризуется неоднозначностью динамики ее отдельных составляющих, в том числе снижением одних показателей, стабилизацией и даже повышением других. Так, в среднем, стандартная работа за 1 мин составляет у 20-29-летних 64,2 кгм, а у 60-69-летних – 48,7 кгм, восстановление работоспособности после стандартного отдыха 56,4 % и 62,1 % соответственно, сила кисти 46,4 кг и 31,9 кг соответственно, максимальное число движений за 10 сек 48,7 и 26,3 соответственно. При этом, несмотря на значительное снижение скорости двигательных актов и возможности выполнять сложно скоординированные движения, способность к длительной мышечной работе при старении сохраняется [18].
У лиц 80-летнего возраста мышечная сила составляет менее половины ее уровня у 30-летних субъектов. В результате снижается нагрузка на скелет и активируется ремоделирование с преобладанием резорбционного компонента – эффект аналогичный наблюдаемому при иммобилизации [38]. Однако рассматривать нарушение функции мышц как основную причину потери костной массы вряд ли возможно, так как эта потеря начинается уже в 25-30 лет, т. е. в период высокой локомоторной активности. Потеря же мышечной массы, ослабление силы мышц и нарушение их функции происходят существенно позже [28, 44]. H.E. Мeema [48] нашел, что в то время как средние величины минеральной плотности кости у женщин в постменопаузальном периоде были заметно ниже, средний диаметр мышц практически не менялся, но после достижения 57 лет по мере старения происходила их прогрессирующая атрофия. Следовательно, снижение мышечной силы является существенным элементом патогенеза потери костной массы у лиц преклонного возраста. Таким образом, роль мышечного фактора в патогенезе потери костной массы можно разделить на два этапа, на первом она незначительна, на втором снижение нагрузок стимулирует этот процесс.
В результате, как видно из вышеизложенного, при старении происходят следующие процессы:
-
- ослабление механического сигнала, нарастающее по мере уменьшения деформаций межклеточного вещества в связи с изменением
ультраструктуры костного матрикса;
-
- увеличение порога чувствительности клеток к механическим сигналам;
-
- снижение величины механических нагрузок, связанное с падением мышечной массы.
В связи с этим встает вопрос: действительно ли формирование нарушений архитектуры губчатой кости является простым результатом повреждений ткани под влиянием механических нагрузок и последующей ее резорбции остеокластами? Постараемся ответить на него.
-
VI. ФОРМИРОВАНИЕ НАРУШЕНИЙ ТРАБЕКУЛЯРНОЙ АРХИТЕКТУРЫ ПРИ СТАРЕНИИ
Возникающие нарушения архитектуры приводят к уменьшению несущих характеристик трабекулярной кости и возникновению остеопо-ротических переломов позвонков [42], в которых трабекулярная кость несет более 90 % нагрузки. При подобных компрессионных переломах высота позвонков сокращается на 20-25 % [55]. Снижение трабекулярной плотности при старении вызвано разрушением трабекул под нагрузкой и при их деформации изгибом [27].
Специальным методом окраски идентифицировано три типа повреждений трабекул: дискретные микротрещины, поперечно-штриховые ультрамикротрещины и диффузные ультраповреждения [34, 35]. Первые имеют длину около 80 µм, а остальные – существенно меньше [62]. Патогенетической основой формирования микроповреждений является пластинчатая структурная организация ткани и особенности вектора нагрузки, который не всегда соответствует основному направлению трабекул [33, 34].
Однако концепция нарушения трабекулярной структуры в результате простого разрушения трабекул под влиянием механических нагрузок сомнительна, так как не согласуется с современными клинико-экспериментальными данными. Как подчеркивает N.L. Fazzalari [35], нормальная трабекулярная архитектура кости имеет структуру, способную выдержать высокий уровень деформаций и ассоциированных микротравм, причем поврежденная ткань может быть восстановлена ремоделированием кости. Существует равновесие между возникновением и регенерацией микроповреждений путем ремоделирования, гарантирующее, что они не будут накапливаться и способствовать росту риска усталостного перелома кости. В том же случае, если процесс ремоделирования не успел закончиться и произошел перелом трабекулы, ее восстановление происходит путем репаративного остеогенеза с образованием микромозоли.
Как отмечено выше, при старении процесс ремоделирования часто останавливается на стадии резорбции. Происходит истончение трабекул, причем в первую очередь разрушаются поперечные. Подобные изменения увеличивают амплитуду отклонений продольных трабекул при их изгибе. Соответственно возрастают локальные деформации растяжения и сжатия, а значит и величина механического сигнала, управляющего ремоделированием костной ткани.
На этой основе мы сформулировали следующую рабочую гипотезу:
изменение архитектуры губчатой кости при старении есть результат целенаправленного многоэтапного адаптивного процесса ремоделирования в ответ на увеличение ригидности внеклеточного матрикса и порога механосенсорной чувствительности клеток остеоцитарного ряда, а также на снижение механических нагрузок;
изменение архитектуры трабекулярной ткани при старении направлено на обеспечение необходимой величины деформаций межклеточного матрикса в конкретных условиях выполнения локомоторных функций и соответственно достаточных значений механического сигнала, инициирующего процесс механотрансдукции;
негативным моментом этих адаптивных процессов является снижение прочностных свойств губчатой кости в связи с накоплением сдвигов в ее архитектуре.
Рассмотрим аргументы в пользу данной гипотезы, используя общетеоретическое моделирование на основе законов теории сопротивления материалов.
-
VII. ОБЩЕТЕОРЕТИЧЕСКОЕ МОДЕЛИРОВАНИЕ ИЗМЕНЕНИЯ АРХИТЕКТУРЫ ТРАБЕКУЛЯРНОЙ КОСТИ В УСЛОВИЯХ ДЕЙСТВИЯ ФАКТОРОВ СТАРЕНИЯ
Поскольку прямые измерения напряже-ний/деформаций отдельной трабекулы невозможны, созданы компьютерные детализированные модели трехмерной архитектуры трабекулярной кости, построенные на основе методов визуализации высокого разрешения, например, микрокомпьтерной томографии. Однако анализ роли локальных нарушений трабекулярной структуры требует разработки универсальной трабекулярной модели кости, позволяющей изучать механику «типичной», а не индивидуальной кости, так как трабекулярные участки костей состоят из тысяч трабекул с неравномерной и уникальной геометрией, требующей очень высокой сложности моделирования. Поэтому применяется метод приближений, и трабекулы рассматриваются как гладкие стандартные элементы. В результате ни одна из опубликованных моделей не использует реальные архитектурные данные для анализа геометрии трабекулярных структур и их изменений под влиянием нагрузки [30]. Другими словами, наличие современных технологий обеспечивает возможность разработки только усредненных структур, требуя суперкомпьютерных возможностей, а в результате представляет только усредненные и очень приблизительные данные. Например, с помощью этого подхода D. Dagan [30] получил расчетные значения модуля упругости 150 MРa, в то время как J.D. Currey [29] посредством объективных физических методов – 17,2 GРa. Подобные различия являются ярким подтверждением того, что результаты компьютерного моделирования зачастую неадекватны величине затрат, необходимых на их получение.
В связи с этим в настоящем исследовании используется простейший вариант идеализированной прямоугольной стержневой модели трабекулярной ткани позвонка, находящегося под действием вертикальной сжимающей нагрузки (рис. 1). Все продольные стержни-трабекулы одинаковы и имеют постоянное сечение. В зависимости от условий, рассматриваемых ниже, эта нагрузка может вызывать потерю устойчивости продольной трабекулы, изгибающейся по синусоиде.
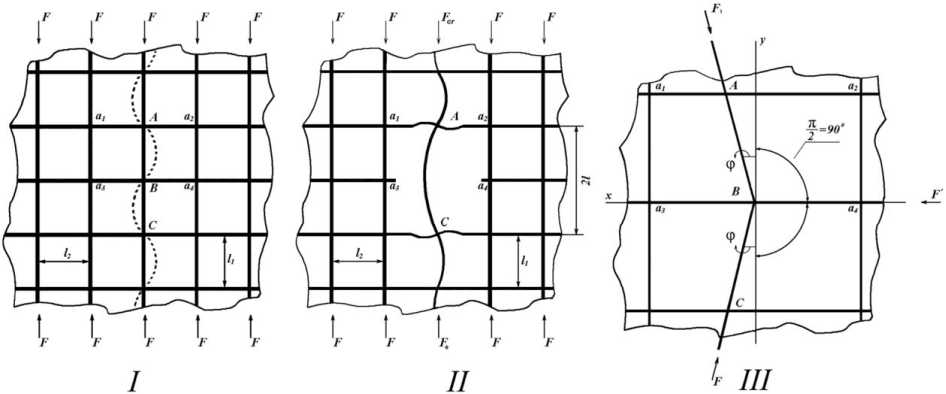
Рис. 1. Идеализированная модель участка решетчатой кости позвонка. На схеме : I – форма потери устойчивости продольной трабекулы при ее сжатии предельной нагрузкой; II – форма потери устойчивости продольной трабекулы при ее сжатии продольной нагрузкой в условиях разрушения пары противоположных поперечных трабекул. III – фрагмент трабекулярного узла при неперпендикулярности продольной трабекулы поперечным. Обозначения : вертикальные линии – продольные трабекулы; горизонтальные линии – поперечные трабекулы; F – сжимающая сила; F cr - критическая сжимающая сила; l 1 – расстояние между поперечными трабекулами; l 2 – расстояние между продольными трабекулами
Поперечные трабекулы здесь играют роль шарнирных опор, препятствующих поперечным перемещениям, но не мешающих повороту соответствующих сечений продольных трабекул. Опорные закрепления продольной трабекулы остаются на одной прямой, соответствующей начальной линии идеализированной решетки. В этих условиях в поперечных трабекулах практически не возникает реактивных усилий. Используя эту модель, поэтапно рассмотрим механизмы разрушения трабекул. При этом мы ис- ходим из того, что на каждом этапе все варианты изменений происходят одновременно, но преобладает только один.
Этап 1 . Истончение продольной трабекулы приводит к снижению ее предельной несущей способности в момент потери устойчивости. Соответстующая критическая сила Fcr , рассчитывается по формуле Эйлера [8]:
2 EI min
Fcr 2 , l1
где: E – модуль упругости материала трабекулы, Imin –минимальный момент инерции поперечного сечения трабекулы; l1 – длина трабекулы.
Длина l1 остается практически постоянной, а Imin снижается за счет уменьшения толщины трабекулы согласно формуле [8]:
πR4
Imin
.
Как следует из этой формулы, при остеопо-ротическом уменьшении толщины трабекулы (t=2R) (по данным D. Dagan, в 3,3 раза [30] с 283µм до 86µм) I min сокращается в 117 раз. Это позволяет утверждать, что влияние E на изменение критической силы малосущественно по сравнению с влиянием толщины. Проведя перерасчет, получаем, что при условии увеличения E в 1,3 раза и уменьшении Imin в 117 раз, Fcr снизится в 90 раз. Например, если при нормальной толщине трабекул средняя величина F cr на позвонок на уровне L 2 составляет 2620 N [63], то при средней толщине трабекул 86 μм их несущая способность окажется равной только 29,1 N.
Снижение несущей способности приводит к потере устойчивости трабекулы изгибам при меньшей нагрузке, что обеспечивает усиление деформаций. Рассмотрим этот процесс на примере локальной потери устойчивости отдельной трабекулы. До того, как это произошло, при сжатии прямой трабекулы (рис. 2) по всей ее толщине наблюдаются одинаковые деформации (ε8) сжатия согласно формуле (1).
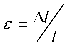
относительная деформация
Рис. 2. Различная степень изгиба отдельной трабекулы. Обозначения : F –сила; t – толщина трабекулы; R – радиус кругового сечения трабекулы; v – прогиб изогнутой оси в середине длинны трабекулы; CD – поперечное сечение в середине трабекулы
Трабекула при потере устойчивости изгибается (см. рис. 2, А и Б), и в этом варианте с ее левой стороны имеем деформацию сжатия, а справа – растяжения. Если прогиб (v) в середине трабекулы незначительный и составляет 0,25R (см. рис. 2, Б), то максимальная относительная деформация от изгиба вычисляется по формуле (1) и равна F . В итоге, согласно закону со-EA вместного действия изгибающего момента и сжимающей силы [8], относительная деформация в точке D удваивается, а в точке С равна нулю. При дальнейшем увеличении прогиба, достигающем v = R, деформации существенно нарастают и в точке С достигают значений
-
5 F , а в точке D 3 F . Таким образом, EA EA
деформации уже на начальных стадиях изгиба значимо превышают деформации сжатия прямой трабекулы и продолжают возрастать при увеличении v .
Следовательно, истончение трабекул носит адаптивный характер и обеспечивает увеличение деформаций внеклеточного матрикса и соответственно величины механического сигнала, инициирующего механотрансдукцию при сохранении трабекулярной архитектуры, но за счет уменьшения несущей способности всего трабекулярного массива кости.
Этап 2. Исчезновение одной из двух поперечных трабекул. Как отмечено выше, при строгой перпендикулярности поперечных трабекул к продольным в поддерживающих трабекулах мы имеем практически нулевые усилия. Если условие перпендикулярности не соблюдается (см. рис 1 III), то в поперечной трабекуле появляется усилие F´, которое можно найти по формуле F = 2Fsinф, вытекающей из правила равновесия. Отсюда следует, что если, к примеру, φ = 3º (sinφ ~ 0,05) то F’=0,1F. Только когда φ достигает 30º (sinφ ~ 0,5), получаем F =F. В физиологических условиях столь значительное отклонение от перпендикулярности осей продольной и поперечной трабекул встречается редко, так как эти оси формируются соответственно траекториям силовых линий, которые строго перпендикулярны друг другу в точках пересечения. Следовательно, в подавляющем большинстве случаев величина φ мала, и поэтому в поперечных трабекулах напря-жения/деформации существенно меньше, чем в продольных. Кроме этого, в неповрежденной решетке деформации поперечной трабекулы зависят от податливости ее противоположной пары. Поэтому, по мере уменьшения величины деформаций внеклеточного матрикса при старении, возрастает вероятность того, что в некоторых поперечных трабекулах механический сигнал выйдет за пределы нижнего порога чувствительности клеток. В результате эти трабекулы резорбируются, что обеспечит возникновение двух эффектов:
-
увеличится возможная степень деформаций оставшейся противоположной трабекулы, так как сопротивление этой деформации прекращается;
формируются ошибки архитектуры трабекулярной решетки, снижающие ее устойчивость к экстремальным нагрузкам по сравнению с первым этапом.
Таким образом, результатом резорбтивного удаления одной из двух противоположных поперечных трабекул является локальное усиление деформаций оставшейся, то есть адаптивный процесс направлен на сохранение величины механического сигнала в пределах нижней границы чувствительности клеток. Однако это происходит за счет формирования ошибок в трабекулярной структуре и локального снижения ее устойчивости к нагрузкам в целом.
Этап 3. Удаление второй поперечной трабекулы . Механизм этого процесса аналогичен рассмотренному выше (этап 2). В результате исчезновения двух смежных поперечных трабекул a 3 и a 4 из двух продольных АВ и ВС формируется одна – AC длиной 2 l (рис. 1). В данном случае, как следует из формулы (2), предельная нагрузка F cr для этой трабекулы в 4 раза меньше, чем для соседних с неповрежденными поперечными связями.
В подобных условиях, пока сжимающее усилие на одну трабекулу не достигло значений Fcr, внешняя нагрузка распределяется равномерно, и ликвидация двух поперечных трабекул a3 и a4 остается незамеченной. После достижения значения Fcr на трабекулу AC , которое для нее является предельным, рост нагрузки прекращается по всей линии (выше и ниже) расположенных по оси AC трабекул. В реальных условиях этот эффект заметен, примерно, на 3-6 ячеек вверх и вниз от места разрыва поперечных трабекул a1 и a2, так как поперечные трабекулы в отличие от нашей идеальной модели не вполне перпендикулярны продольной.
В результате потери устойчивости трабеку- лы АС происходит перераспределение нагрузки на соседние параллельно расположенные трабекулы, что приводит к увеличению их деформаций. По данным Dagan это вызывает их изгиб и существенное усиление напряжения по сравнению с величиной на этом участке, аналогичной неповрежденной решетки [30].
Таким образом, результатом резорбции двух противоположных поперечных трабекул является усугубление деформаций как в данной продольной трабекуле, так и в соседних параллельных с ней, что можно рассматривать как адаптивный процесс, увеличивающий деформации продольных трабекул. Однако это сопровождается одновременным снижением деформаций в трабекулах, расположенных выше и ниже по этой оси, нарастанием локальных ошибок трабекулярной архитектуры и соответственно дальнейшим снижением устойчивости к нагрузкам всего трабекулярного массива по сравнению с вторым этапом.
Этап 4 . Дифференцированное удаление более тонких продольных трабекул связано с тем, что величина деформаций при изгибе зависит от толщины трабекулы. Как было сказано выше, предельная несущая способность трабекулы в момент потери ее устойчивости изгибом (критическая сила F cr ) определяется формулой (2). При изгибе (рис. 3) изгибающий момент ( М ) в этом сечении равен:
M F cr v 2 EI 2 min v , (3)
где v – максимальный прогиб в середине длины.
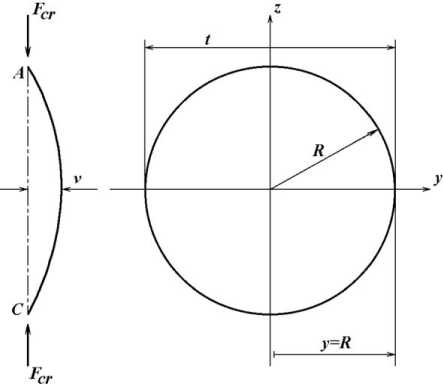
Рис. 3. Схематическое изображение изогнутой трабекулы и ее поперечного сечения. Обозначения : F cr -критическая сила; t – толщина трабекулы; R – радиус кругового сечения трабекулы; y max – координата точки, наиболее удаленной от нейтрального слоя
Возникающие в поперечном сечении трабекулы нормальные напряжения вычисляем по формуле:
My , (4)
min где y – расстояние от нейтральной линии (проходящей через центр тяжести сечения) до точки, в которой определяется напряжение.
При изгибе связь между нормальным напряжением (σ) и относительной деформацией (ε) может быть принята в простейшей форме:
а
. (5)
E
С учетом формулы (4) получаем:
My
, EI min
а с учетом формулы (3) имеем:
2 yv l 2
.
Для упрощения последующих расчетов представим толщину t трабекулы как долю от ее длины t=kl, где 0< k< 1 , а сечение трабекулы в виде круга с радиусом R . Тогда t=2R=kl или
R = 0,5kl . (8)
Максимальные деформации в поперечном сечении изогнутой трабекулы возникнут при y=R=0,5kl. (9)
Подставляя формулы (8) и (9) в формулу (7), получаем v 5k . (10)
Исходя из того, что средняя длина трабекулы составляет 1 мм, средняя толщина нормальной трабекулы – 0,283 мм, остеопоротической – 0,100 мм и меньше [30], а нижний порог дефор- маций – около 1700 μs [56], подставляем эти данные в формулу (9) и получаем необходимый прогиб этих трабекул, который должен для ос-теопоротической трабекулы быть в три раза больше, чтобы достичь той же самой деформации. Однако величина прогиба ограничена соседними трабекулами, и, следовательно, при одной и той же нагрузке в нормальной трабекуле интенсивность механического сигнала выше, чем в остеопоротической. Поэтому для тонкой трабекулы при прочих равных условиях риск резорбции выше.
Подобное явление можно рассматривать как парадоксальный эффект адаптации, обеспечивающей увеличение значений механического сигнала на первом этапе за счет истончения трабекул, тогда как на четвертом этапе это, наоборот, ведет к снижению его относительной величины. Данный процесс можно рассматривать как адаптивную дифференцированную резорбцию более тонких (остеопоротических) трабекул, приводящую к такому накоплению ошибок архитектуры, при котором физиологические нагрузки становятся экстремальными. В этих условиях энергетически более выгодно увеличить толщину наиболее толстых трабекул, чем тонких.
Этап 5. Перераспределение нагрузки с губчатой на кортикальную кость влечет за собой возрастание ее массы, например, в позвонках [32]. Однако этот прирост кортикальной кости оказывается недостаточным и не соответствует необходимому ни по скорости, ни по объему, в результате чего происходят компрессионные переломы.
ЗАКЛЮЧЕНИЕ
Все вышесказанное позволяет утверждать, что формируемые нарушения архитектуры губчатой ткани носят адаптивный характер и направлены на обеспечение процесса механотрансдукции в ответ на изменение деформационных возможностей внеклеточного матрикса, чувствительности костных клеток к механическому сигналу и силы мышц при старении. Другими словами, остеопороз – это не болезнь, а форма адаптации. В этой связи мы согласны с И.В. Давыдовским [10], который подчеркивал, что довольно широко распространено ложное представление о принципиальной раздельности физиологического и патологического. В патологии (как и в болезни) усматривают два раздельных аспекта, а именно “нарушение функций” и действие защитно-физиологических механизмов, то есть болезнь и “физиологическую меру” против нее. Автор подчеркивает, что введение в обиход слова “защита” вообще не приемлемо при анализе биологических процессов, так как каждый патологический процесс имеет свой прототип в физиологии. С этих позиций адаптация – не синоним здоровья, а болезнь - не отрицание, а форма адаптации.
Все это позволяет выделить три последовательно включающихся патогенетических механизма, приводящих к снижению механического стимула, инициирующего локально процесс механотрансдукции.
Первоначально преобладает возрастзависи-мый рост минерализации внеклеточного матрикса, усиливающий его жесткостные свойства и соответственно снижающий величины механического сигнала при одной и той же нагрузке.
По мере углубления возрастных регуляторно-метаболических сдвигов (например, потеря репродуктивной функции) увеличивается порог чувствительности клеток к механическим сигналам, что требует более высокой локальной нагрузки для дополнительного прироста деформаций.
При достижении старческого возраста происходит существенное снижение мышечной силы и соответственно снижение механических нагрузок, обеспечивающих необходимую величину механических сигналов в физиологических условиях.
В ответ на описанные сдвиги в губчатой ткани развивается 5-этапный адаптивный процесс перестройки трабекулярной архитектуры с преобладанием на каждом этапе формирования сдвигов определенного типа. Основываясь на представленной в настоящей работе концепции, для научно-клинических разработок можно выделить следующие направления.
Разработка методов укрепления мышечной системы для сохранения величины сигнала у лиц с высоким риском развития остеопороза (низкая пиковая костная масса, хроническая патология и т. д.), начиная с 30-летнего возраста, то есть с момента начала потери костной массы, а не после возникновения нарушения архитектуры губчатой кости, что соответствует принципам профилактической медицины.
Профилактика потери костной массы возможна также посредством создания медикаментозных препаратов, обеспечивающих (1) уменьшение минерализации костной ткани с помощью регуляторов остеоцитарного насоса для обеспечения вымывания Са++ из костной ткани [4, 16] и (2) снижающих порог инициации электрических импульсов. Последнее обусловлено тем, что на начальном этапе механотрансдукции происходит инициация электрических сигналов, распространяющихся по остеоцитар-ным отросткам от клетки к клетке (аналогично распространению электрического импульса в нервной ткани), и эти сигналы обеспечивают передачу информации об изменении механических напряжений [57]. Это позволит оптимизировать процессы ремоделирования на более низком уровне деформаций костной ткани при изменении механических нагрузок.
