Активные формы кислорода как архитекторы редокс-процессов в формировании стрессоустойчивости зерновых культур в условиях глобального изменения климата. Часть 1

Автор: Кади А.М.Я.
Рубрика: Актуальные проблемы развития пищевых и биотехнологий
Статья в выпуске: 4 т.13, 2025 года.
Бесплатный доступ
В статье представлены современные представления о роли активных форм кислорода (ROS) в регуляции экспрессии генов, ответственных за модификации гистонов у зерновых культур в условиях действия абиотических стрессов. Показано, что ROS выполняют не только деструктивную, но и сигнальную функцию, участвуя в формировании редоксгомеостаза и адаптивных ответов растений. Рассмотрены механизмы генерации и утилизации ROS, их участие в регуляции транскрипционных каскадов и взаимодействие с антиоксидантной и гормональной системами. Особое внимание уделено взаимосвязи между редокссигнализацией и эпигенетическими изменениями, включая ацетилирование и метилирование гистонов, которые обеспечивают перестройку хроматиновой структуры и формирование стрессовой памяти. Работа обобщает экспериментальные данные о редоксэпигенетической регуляции у пшеницы и других зерновых культур, показывая, что ROSиндуцированные эпигенетические модификации лежат в основе гибкости и наследуемости адаптационных реакций. Дополнительно обсуждаются механизмы участия ROS в координации сигналов между клеточными органеллами, а также их роль в формировании системных адаптационных ответов растений. Подчёркивается значение ROS как ключевого звена, связывающего метаболические, транскрипционные и хроматиновые уровни регуляции. Эпигенетическая сеть, управляемая ROS, представляет собой молекулярный компас для адаптации к стрессу и создания устойчивых к климату и высокоурожайных зерновых культур. Рассмотрение ROS как центрального узла окислительновосстановительной регуляции открывает перспективы для целенаправленного управления редоксэпигенетическими процессами в селекционных и биотехнологических стратегиях, направленных на повышение продуктивности и стрессоустойчивости растений в условиях глобального изменения климата.
Реактивные формы кислорода (ROS), редокс-сигнализация, злаковые культуры, эпигенетическая регуляция, модификации гистонов, стрессовая память, адаптация растений
Короткий адрес: https://sciup.org/147252423
IDR: 147252423 | УДК: 630.165.3 | DOI: 10.14529/food250401
Reactive oxygen species as architects of redox dynamics and adaptive potential in cereal crops under global climate change. Part I
The article presents current insights into the role of reactive oxygen species (ROS) in regulating the expression of genes responsible for histone modifications in cereal crops under abiotic stress conditions. It is shown that ROS perform not only destructive but also essential sig-naling functions, participating in the maintenance of redox homeostasis and the orchestration of adaptive plant responses. The mechanisms of ROS generation and scavenging are analyzed, along with their involvement in the regulation of transcriptional cascades and their interactions with an-tioxidant and hormonal systems. Special attention is given to the relationship between redox sig-naling and epigenetic modifications, including histone acetylation and methylation, which drive chromatin remodeling and the establishment of stress memory. The work summarizes experimen-tal evidence on redox-epigenetic regulation in wheat and other cereals, demonstrating that ROS-induced epigenetic modifications underlie the flexibility and heritability of adaptive responses. Additionally, the mechanisms of ROS involvement in the coordination of inter-organellar signal-ing and systemic plant responses are discussed. The study emphasizes the significance of ROS as a key integrator linking metabolic, transcriptional, and chromatin-level regulation. The epigenetic network governed by ROS is proposed as a molecular compass guiding plant adaptation to stress and the development of climate-resilient, high-yielding cereal crops. Considering ROS as a central hub of redox regulation opens new perspectives for targeted manipulation of redox-epigenetic mechanisms within breeding and biotechnological strategies aimed at improving plant productiv-ity and stress tolerance under global climate change.
Текст научной статьи Активные формы кислорода как архитекторы редокс-процессов в формировании стрессоустойчивости зерновых культур в условиях глобального изменения климата. Часть 1
Зерновые культуры являются краеугольным камнем обеспечения продовольственной безопасности и адаптации к глобальным изменениям климата [1]. Зерновые культуры, такие как рис (Oryza sativa), пшеница (Triticum aestivum), кукуруза (Zea mays), ячмень (Hordeum vulgare), сорго (Sorghum bicolor) и просо в совокупности обеспечивают более половины калорийности пищи, потребляемой человечеством [2, 3]. Помимо своей питательной роли, эти культуры являются основой экономической стабильности в мире за счет формирования рационов кормления скота и биологических материалов для промыш- ленного сектора [4, 5]. Их продуктивность напрямую определяет устойчивость продовольственных систем в условиях роста населения, урбанизации и сокращения пахотных земель [6]. Однако поддержание и повышение урожайности зерновых в условиях быстро меняющихся климатических условий остается одним из определяющих факторов благополучия и экономической устойчивости территорий [6, 7]. Современное сельское хозяйство сталкивается с одновременным давлением абиотических стрессоров – в частности, засухи, засоления, экстремальных температур и дефицита питательных веществ – и биотических ограничений, включая патогены и вреди- телей [8, 9]. Эти факторы нарушают клеточный окислительно-восстановительный гомеостаз и метаболический баланс, провоцируя избыточное образование реактивных форм кислорода (ROS) в хлоропластах, митохондриях и пероксисоме, которая содержит ферменты, участвующие в окислительновосстановительных реакциях [6, 10]. В то время как низкие уровни ROS выступают в качестве ключевых сигнальных молекул, их чрезмерное накопление приводит к окислительному повреждению липидов, нуклеиновых кислот и белков и как следствие – замедлению роста и потере урожая [8, 11]. Усиление глобального изменения климата ускоряет образование ROS за счет усиленного фотодыхания растений, в то время как засуха и засоление ограничивают ассимиляцию CO2 и транспорт электронов, усугубляя окислительный стресс [7, 9]. Таким образом, тонкое равновесие между генерацией и сжиганием ROS является центральным фактором, определяющим устойчивость к стрессу и продуктивность у злаков [9, 10].
Накопленные данные свидетельствуют о том, что ROS являются не просто побочными продуктами стресса, а выступают в качестве регуляторов эпигенетического механизма [2, 5]. Чувствительные к окислительным реакци- ям факторы транскрипции, сигнальные киназы и гистон-модифицирующие ферменты образуют взаимосвязанные сети, которые преобразуют окислительные сигналы в изменения на уровне хроматина [4, 6]. Такое ROS-опосредованное эпигенетическое перепрограммирование (см. рисунок) определяет метаболические сдвиги, гормональные перекрестные связи и пластичность развития, в итоге формируя морфологические и биохимические фенотипы растений [12]. Среди зерновых культур пшеница представляет собой убедительную модель для изучения эпигенетической модуляции под действием ROS благодаря своему сложному геному, широкому экологическому распространению и центральной роли в мировом питании [10].
Производство пшеницы сталкивается с серьезными ограничениями, связанными с жарой, засухой и засолением, все из которых провоцируют окислительный дисбаланс [10]. Недавние транскриптомные и эпигеномные исследования показали, что пшеница отвечает на ROS-индуцирующие стрессы дифференциальной экспрессией генов, кодирующих ацетилтрансферазы гистонов (HATs), деацетилазы гистонов (HDACs), метилтрансферазы гистонов (HMTs) и деметилазы (HDMs) [12, 13]. Эти изменения совпадают с изменением экспрес-
Стресс-факторы
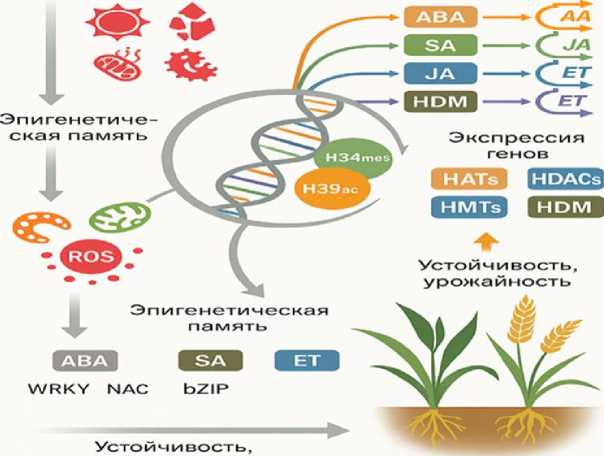
урожайность
Схематическое представление роли активных форм кислорода (ROS) в формировании эпигенетической памяти и стрессоустойчивости злаков. ROS, взаимодействуя с гормональными и эпигенетическими регуляторами (HATs, HDACs, HMTs, HDMs), модулируют экспрессию генов, обеспечивающих адаптацию и повышение урожайности
сии антиоксидантных ферментов и генов биосинтеза фитогормонов, что указывает на интегративную регуляторную систему [13, 15].
Кроме того, пшеница демонстрирует феномен стрессовой памяти, когда предшествующее воздействие окислительного стресса или засухи закладывает основу для будущих реакций [11, 16]. Это состояние «праймирова-ния» было связано с постоянными метками гистонов, в частности H3K4me3 и H3K9ac, в локусах, реагирующих на стресс [17]. Подобные находки означают, что ROS-индуци-рованные сигнатуры хроматина могут лежать в основе как непосредственной толерантности, так и трансгенерационной адаптации, представляя собой эпигенетический путь для улучшения сельскохозяйственных культур за пределами традиционной селекции [18]. Учитывая совпадающую роль ROS, модификаций гистонов и фитогормональной сигнализации в определении последствий стресса для растений, крайне необходимо комплексное понимание этих сетей [2, 5]. В данном обзоре обобщены современные представления о том, как ROS регулируют экспрессию генов модификации гистонов в зерновых – с акцентом на пшеницу – и как это влияет на фитогормоны, эпигенетическое перепрограммирование и фенотипические характеристики [3]. Соединяя молекулярную сигнализацию с морфологическими и биохимическими проявлениями, эта работа направлена на определение ROS-эпигенетической-гормональной оси как центральной детерминанты устойчивости к стрессу и оптимизации урожайности у зерновых [5].
Реактивные формы кислорода (ROS) в растениях и их сигнальные роли. Общие механизмы образования ROS
Реактивные формы кислорода (ROS) – это высокореактивные производные молекулярного кислорода, играющие двойную роль: при низких концентрациях они служат сигнальными посредниками, а при избытке – цитотоксичными агентами [6–8]. В злаковых культурах (пшеница, рис, кукуруза) ROS образуются постоянно в различных компартментах – хлоропластах, митохондриях, пероксисомах и апопласте [8–10].
Основные типы – супероксидный анион ( O2- ), перекись водорода (H2O2), гидроксильный радикал (·OH) и синглетный кислород (1O2) – возникают преимущественно в фото- синтетических и дыхательных электроннотранспортных цепях [8–11].
Редокс-гомеостаз поддерживается антиоксидантными ферментами (SOD, CAT, APX) и низкомолекулярными компонентами (аскорбат, глутатион, каротиноиды) [10–14].
Нарушение этого баланса вызывает окислительный стресс, повреждение мембран, ДНК и белков [6, 8, 13].
Тем не менее растения, включая злаки, используют временные всплески ROS в качестве сигнальных импульсов, активирующих адаптивные генетические сети [5, 10, 15].
ROS как интеграторы восприятия стресса у злаков
ROS действуют как ранние мессенджеры, соединяя сигналы окружающей среды с транскрипционными и гормональными ответами [6, 7, 8]. Во время засухи и теплового стресса хлоропласты генерируют H 2 O 2 через фотодыхательную активность гликолят-оксидазы, а митохондрии усиливают поток электронов, повышая уровень ROS [8, 16].
У риса NADPH-оксидаза OsRbohB опосредует апопластический окислительный взрыв, активируя ABA-респонсивные факторы ABF и AREB [9, 17]. Аналогично, у пшеницы засуха вызывает накопление H 2 O 2 и усиливает экспрессию TaCAT1 и TaSOD2, поддерживая фотосистему и формирование зерна [18, 19]. При солевом стрессе ROS функционируют как вторичные мессенджеры в Ca2+-зависимой сигнализации: у кукурузы ZmRbohD активирует Ca2+-каналы и MAPK-каскады, регулируя гены антиоксидантов и ионного гомеостаза [20–22]. Кроме того, контролируемое образование ROS улучшает баланс Na+/K+ и осмотическую адаптацию у устойчивых генотипов [21, 23]. В биотических взаимодействиях (например, при инфекции Blumeria graminis у ячменя или Magnaporthe oryzae у риса) окислительный взрыв в апопла-сте запускает лигнификацию, отложение каллозы и гибель клеток в зоне контакта с патогеном, ограничивая его распространение [24, 25]. Таким образом, ROS интегрируют абиотические и биотические сигналы, служа универсальным регулятором роста, иммунитета и адаптации.
Сенсинг ROS и нисходящая сигнализация
Механизм, с помощью которого растения воспринимают ROS, включает как прямое окисление сенсорных белков, так и опосредо- ванные окислительно-восстановительными процессами посттрансляционные модификации [6, 9]. У злаков окисление цистеина факторов транскрипции, таких как семейства WRKY, NAC и ZAT, действует как молекулярный переключатель, контролирующий активацию стрессовых генов [16, 18].
Например, в рисе H 2 O 2 -индуцированное окисление OsWRKY30 усиливает его ДНК-связывающую активность с промоторами, реагирующими на стресс [17]. H 2 O 2 также диффундирует через мембраны посредством аквапоринов (PIPs), обеспечивая распространение окислительных сигналов от клетки к клетке [7, 19]. Эта волна ROS, впервые описанная в Arabidopsis, была продемонстрирована в кукурузе, где системное перемещение H 2 O 2 координирует акклиматизацию дистальных тканей при тепловом стрессе [7]. Взаимодействие между ROS и потоками Ca2+ является еще одной консервативной особенностью: окислительная активация каналов Ca²⁺ запускает быстрое фосфорилирование MAPK, что приводит к транскрипционной активации защитных и антиоксидантных генов [22].
Перекрестное взаимодействие между ROS и фитогормональными сетями
Сигнализация ROS в злаках тесно интегрирована с фитогормонами, включая абсцизовую кислоту (ABA), жасмоновую кислоту (JA), салициловую кислоту (SA) и этилен (ET) [10, 21]. Во время засухи закрытие стом, вызванное АБА, облегчается за счет ROS, генерируемой сторожевыми клетками через гомологи RbohF в пшенице и рисе [10, 24]. Обратная связь ROS-ABA обеспечивает точную регуляцию стоматита и эффективность водопо-требления [10, 17].
Напротив, во время атаки патогенов SA и JA координируют с ROS экспрессию защитных генов, в то время как этилен модулирует сжигание ROS для предотвращения чрезмерного повреждения [19]. Примечательно, что гормон-ROS также взаимодействует с эпигенетической регуляцией. У кукурузы ROS-опосредованная индукция гистон-ацетилтрансферазы ZmHAC1 при солевом стрессе изменяет доступность хроматина в промоторах, реагирующих на ABA и JA [14]. Такая взаимозависимость подчеркивает, что окислительно-восстановительный статус связывает восприятие окружающей среды и экспрессию генов на эпигенетическом уровне [18].
Баланс гомеостаза ROS для устойчивости к стрессу у злаков
Для оптимальной адаптации злаки должны поддерживать динамический баланс между генерацией и утилизацией ROS [8]. Чрезмерная детоксикация может подавлять сигнализацию, в то время как неконтролируемая ROS приводит к необратимому окислительному повреждению [6, 8]. У пшеницы сверхэкспрессия TaNAC69 повышает устойчивость к засухе за счет тонкой настройки гомеостаза ROS и антиоксидантной активности [25]. Аналогичным образом, линии риса с повышенной экспрессией OsAPX1 демонстрируют улучшенное восстановление после теплового стресса благодаря контролируемому потоку ROS [18]. Интеграция молекулярных, биохимических и фенотипических данных показывает, что умеренное накопление ROS в условиях стресса действует как сигнал «прайминга», который готовит злаковые растения к будущим испытаниям [10, 15]. Это явление, называемое окислительным прай-мингом, часто приводит к устойчивым меткам ацетилирования гистонов в генах, связанных со стрессом, связывая сигнализацию ROS непосредственно с динамикой хроматина [19]. Такие выводы подчеркивают двойственную природу ROS – как опасную, так и перспективную – и показывают, как злаки используют окислительно-восстановительные сигналы для баланса роста, защиты и продуктивности [23].
Совокупность результатов показывает, что ROS не только сигнальные медиаторы, но и регуляторы эпигенетического ландшафта злаков. Модулируя экспрессию гистон-моди-фицирующих ферментов и состояние хроматина, они обеспечивают гибкость транскрипционных сетей и формируют долговременную стресс-память. Редокс-эпигенетический интерфейс становится ключевым механизмом адаптации зерновых к изменяющемуся климату и создаёт фундамент для новых биотехнологических подходов в селекции устойчивых и высокопродуктивных культур.
Заключение
Реактивные формы кислорода (ROS) играют фундаментальную роль в регуляции роста, метаболизма и адаптации злаков. Их двойственная природа – от сигнальных медиаторов до потенциальных агентов повреждения – определяет сложный баланс между защитой и продуктивностью растений. В ус- ловиях абиотического и биотического стресса ROS интегрируют восприятие сигналов с транскрипционными и гормональными ответами, формируя сеть редокс-зависимых реакций.
Ключевым результатом является осознание того, что редокс-состояние напрямую связано с эпигенетической регуляцией: ROS контролируют активность гистон-модифицирую-щих ферментов, изменяют доступность хроматина и обеспечивают формирование стрес- совой памяти. Этот механизм превращает ROS из простых побочных продуктов метаболизма в универсальные сигналы адаптации.
Эпигенетическая сеть, управляемая ROS, представляет собой молекулярный компас для адаптации к стрессу и создания устойчивых к климату и высокоурожайных зерновых культур. Понимание этих механизмов открывает перспективы для направленной селекции и редактирования генома, направленных на усиление природных адаптивных стратегий растений.
